
Mechanisms of Hepatitis C Viral Resistance to Direct Acting Antivirals

Abstract
:1. Introduction
2. Emergence of DAA Resistance
3. Direct Acting Antiviral Drugs
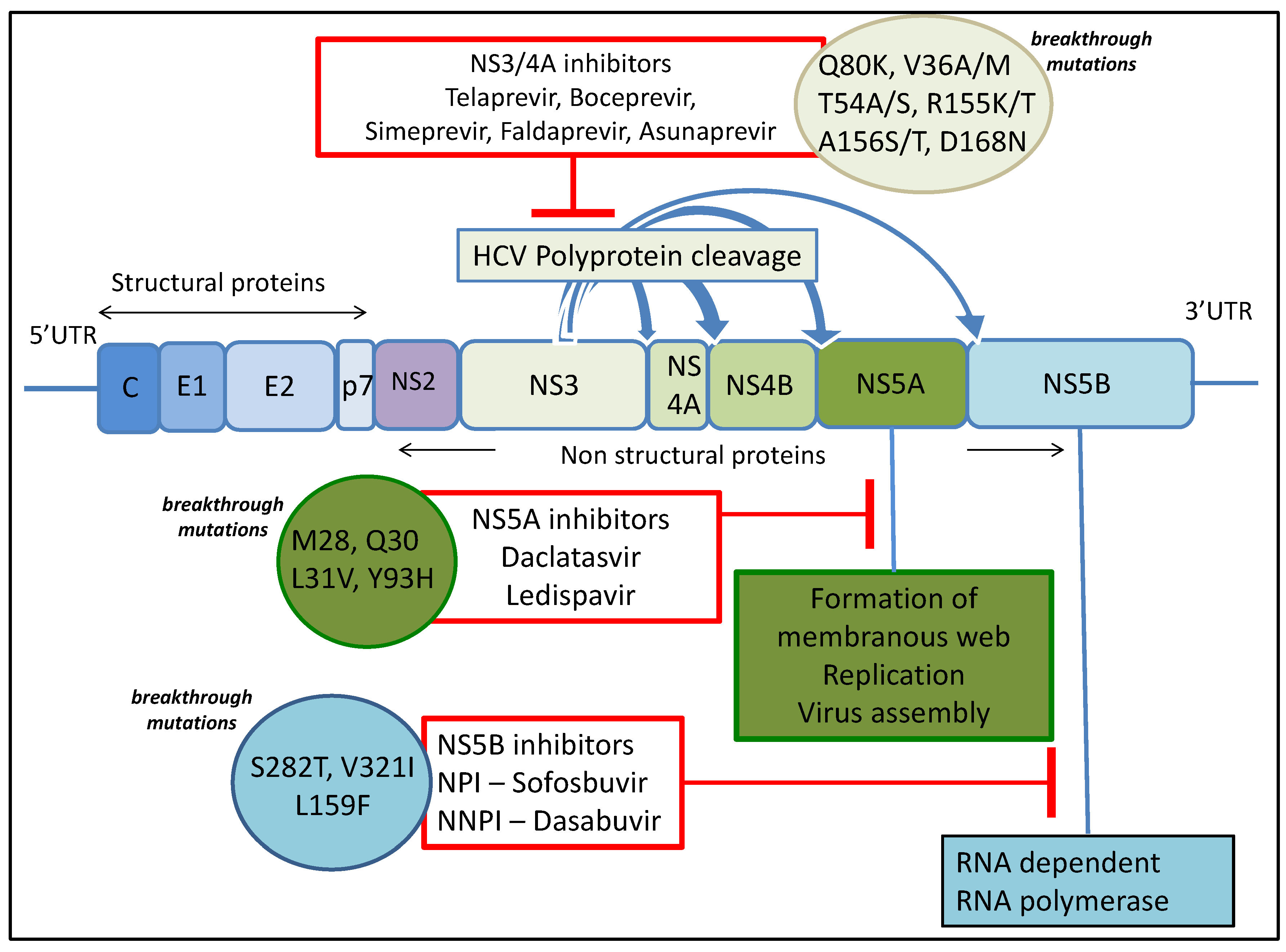
3.1. NS3-4A Protease Inhibitors
3.2. NS5A Inhibitors
3.3. NS5B Inihibitors
3.3.1. Nucleoside Polymerase Inhibitors (NPI)
3.3.2. Non-Nucleoside Polymerase Inhibitors (NNPI)
4. Prevalence before and after Treatment with DAA

{kind=link}
| Drug Class | Example | Common RAV | Resistance | Prevalence |
|---|---|---|---|---|
| NS3/4A inhibitor | telaprevir | V36M | low | <1% |
| boceprevir | T54S/A | low | 2%–3% | |
| simeprevir | V55A | low | 0.4%–3% | |
| asunaprevir | Q80K | low | 0.5%–75% | |
| faldaprevir | R155K | high | <1% | |
| NS5A inhibitor | daclatasvir | M28 | high | 0.5%–4% |
| ombitasvir | Q30 | high | 0.3%–1.3% (geno 1) 50%–100% (geno 3,4) | |
| ledipasvir | L31V | high | 0.9%–6.3% (geno 1) 74%–100%(geno 2,4) | |
| Y93H | high | 1.5%–14% | ||
| NS5B NPI | sofosbuvir | L159F | n.d. | 5.2% |
| V321A | n.d. | 2.2% | ||
| S282R | low | 0.4% | ||
| NS5B NNPI | dasabuvir | C316N | low | 11%–36% |
5. Clinical Significance of Baseline RAVs
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mohd Hanafiah, K.; Groeger, J.; Flaxman, A.D.; Wiersma, S.T. Global epidemiology of hepatitis C virus infection: New estimates of age-specific antibody to HCV seroprevalence. Hepatology 2013, 57, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Humphreys, I.; Flaxman, A.; Brown, A.; Cooke, G.S.; Pybus, O.G.; Barnes, E. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology 2015, 61, 77–87. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Hepatitis C. Available online: http://www.who.int/mediacentre/factsheets/fs164/en/ (accessed on 29 September 2015).
- Lauer, G.M.; Walker, B.D. Hepatitis C virus infection. N. Engl. J. Med. 2001, 345, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, I.M.; McHutchison, J.G.; Dusheiko, G.; di Bisceglie, A.M.; Reddy, K.R.; Bzowej, N.H.; Marcellin, P.; Muir, A.J.; Ferenci, P.; Flisiak, R.; et al. Telaprevir for previously untreated chronic hepatitis C virus infection. N. Engl. J. Med. 2011, 364, 2405–2416. [Google Scholar] [CrossRef] [PubMed]
- Poordad, F.; McCone, J., Jr.; Bacon, B.R.; Bruno, S.; Manns, M.P.; Sulkowski, M.S.; Jacobson, I.M.; Reddy, K.R.; Goodman, Z.D.; Boparai, N.; et al. Boceprevir for untreated chronic HCV genotype 1 infection. N. Engl. J. Med. 2011, 364, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Lawitz, E.; Mangia, A.; Wyles, D.; Rodriguez-Torres, M.; Hassanein, T.; Gordon, S.C.; Schultz, M.; Davis, M.N.; Kayali, Z.; Reddy, K.R.; et al. Sofosbuvir for previously untreated chronic hepatitis C infection. N. Engl. J. Med. 2013, 368, 1878–1887. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, I.M.; Gordon, S.C.; Kowdley, K.V.; Yoshida, E.M.; Rodriguez-Torres, M.; Sulkowski, M.S.; Shiffman, M.L.; Lawitz, E.; Everson, G.; Bennett, M.; et al. Sofosbuvir for hepatitis C genotype 2 or 3 in patients without treatment options. N. Engl. J. Med. 2013, 368, 1867–1877. [Google Scholar] [CrossRef] [PubMed]
- Zeuzem, S.; Dusheiko, G.M.; Salupere, R.; Mangia, A.; Flisiak, R.; Hyland, R.H.; Illeperuma, A.; Svarovskaia, E.; Brainard, D.M.; Symonds, W.T.; et al. Sofosbuvir and ribavirin in HCV genotypes 2 and 3. N. Engl. J. Med. 2014, 370, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Foster, G.R.; Pianko, S.; Brown, A.; Forton, D.; Nahass, R.G.; George, J.; Barnes, E.; Brainard, D.M.; Massetto, B.; Lin, M.; et al. Efficacy of sofosbuvir plus ribavirin with or without peginterferon-α in patients with HCV genotype 3 infection and treatment-experienced patients with cirrhosis and HCV genotype 2 infection. Gastroenterology 2015, 149, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.U.; Lam, N.P.; Dahari, H.; Gretch, D.R.; Wiley, T.E.; Layden, T.J.; Perelson, A.S. Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-α therapy. Science 1998, 282, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Ogata, N.; Alter, H.J.; Miller, R.H.; Purcell, R.H. Nucleotide sequence and mutation rate of the H strain of hepatitis C virus. Proc. Natl. Acad. Sci. USA 1991, 88, 3392–3396. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Thomas, D. Hepatitis C: Mandell, Douglas and Bennett's Principles and Practice of Infectious Diseases, 7th ed.; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Herrmann, E.; Neumann, A.U.; Schmidt, J.M.; Zeuzem, S. Hepatitis C virus kinetics. Antivir. Ther. 2000, 5, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.; Spindler, K.; Horodyski, F.; Grabau, E.; Nichol, S.; VandePol, S. Rapid evolution of RNA genomes. Science 1982, 215, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Martell, M.; Esteban, J.I.; Quer, J.; Genesca, J.; Weiner, A.; Esteban, R.; Guardia, J.; Gomez, J. Hepatitis C virus (HCV) circulates as a population of different but closely related genomes: Quasispecies nature of HCV genome distribution. J. Virol. 1992, 66, 3225–3229. [Google Scholar] [PubMed]
- Bartenschlager, R.; Lohmann, V. Replication of hepatitis C virus. J. Gen. Virol. 2000, 81, 1631–1648. [Google Scholar] [CrossRef] [PubMed]
- Gregori, J.; Esteban, J.I.; Cubero, M.; Garcia-Cehic, D.; Perales, C.; Casillas, R.; Alvarez-Tejado, M.; Rodriguez-Frias, F.; Guardia, J.; Domingo, E.; et al. Ultra-deep pyrosequencing (UDPS) data treatment to study amplicon HCV minor variants. PLoS ONE 2013, 8, e83361. [Google Scholar] [CrossRef] [PubMed]
- Vermehren, J.; Sarrazin, C. The role of resistance in HCV treatment. Best Pract. Res. Clin. Gastroenterol. 2012, 26, 487–503. [Google Scholar] [CrossRef] [PubMed]
- Fafi-Kremer, S.; Fofana, I.; Soulier, E.; Carolla, P.; Meuleman, P.; Leroux-Roels, G.; Patel, A.H.; Cosset, F.L.; Pessaux, P.; Doffoel, M.; et al. Viral entry and escape from antibody-mediated neutralization influence hepatitis C virus reinfection in liver transplantation. J. Exp. Med. 2010, 207, 2019–2031. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, T.L.; Kwong, A.D.; Picchio, G.R. Viral resistance to specifically targeted antiviral therapies for hepatitis C (STAT-Cs). J. Antimicrob. Chemother. 2010, 65, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Pockros, P.J. New direct-acting antivirals in the development for hepatitis C virus infection. Ther. Adv. Gastroenterol. 2010, 3, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Foy, E.; Li, K.; Wang, C.; Sumpter, R., Jr.; Ikeda, M.; Lemon, S.M.; Gale, M., Jr. Regulation of interferon regulatory factor-3 by the hepatitis C virus serine protease. Science 2003, 300, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Foy, E.; Ferreon, J.C.; Nakamura, M.; Ferreon, A.C.; Ikeda, M.; Ray, S.C.; Gale, M., Jr.; Lemon, S.M. Immune evasion by hepatitis C virus NS3/4A protease-mediated cleavage of the toll-like receptor 3 adaptor protein TRIF. Proc. Natl. Acad. Sci. USA 2005, 102, 2992–2997. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Zhong, J.; Chisari, F.V. Inhibition of dsRNA-induced signaling in hepatitis C virus-infected cells by NS3 protease-dependent and -independent mechanisms. Proc. Natl. Acad. Sci. USA 2006, 103, 8499–8504. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, C.; Zeuzem, S. Resistance to direct antiviral agents in patients with hepatitis C virus infection. Gastroenterology 2010, 138, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, T.L.; de Meyer, S.; Bartels, D.J.; Sullivan, J.C.; Zhang, E.Z.; Tigges, A.; Dierynck, I.; Spanks, J.; Dorrian, J.; Jiang, M.; et al. Hepatitis C viral evolution in genotype 1 treatment-naive and treatment-experienced patients receiving telaprevir-based therapy in clinical trials. PLoS ONE 2012, 7, e34372. [Google Scholar] [CrossRef] [PubMed]
- Ogert, R.A.; Howe, J.A.; Vierling, J.M.; Kwo, P.Y.; Lawitz, E.J.; McCone, J.; Schiff, E.R.; Pound, D.; Davis, M.N.; Gordon, S.C.; et al. Resistance-associated amino acid variants associated with boceprevir plus pegylated interferon-α2b and ribavirin in patients with chronic hepatitis C in the sprint-1 trial. Antivir. Ther. 2013, 18, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Halfon, P.; Locarnini, S. Hepatitis C virus resistance to protease inhibitors. J. Hepatol. 2011, 55, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Kuntzen, T.; Timm, J.; Berical, A.; Lennon, N.; Berlin, A.M.; Young, S.K.; Lee, B.; Heckerman, D.; Carlson, J.; Reyor, L.L.; et al. Naturally occurring dominant resistance mutations to hepatitis C virus protease and polymerase inhibitors in treatment-naive patients. Hepatology 2008, 48, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Dierynck, I.; Ghys, A.; Witek, J.; Luo, D.; Janssen, K.; Daems, B.; Picchio, G.; Buti, M.; De Meyer, S. Incidence of virological failure and emergence of resistance with twice-daily vs every 8-h administration of telaprevir in the optimize study. J. Viral Hepat. 2014, 21, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Howe, J.A.; Ogert, R.A.; Zeuzem, S.; Poordad, F.; Gordon, S.C.; Ralston, R.; Tong, X.; Sniukiene, V.; Strizki, J.; et al. Analysis of boceprevir resistance associated amino acid variants (RAVs) in two phase 3 boceprevir clinical studies. Virology 2013, 444, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Zeuzem, S.; Sulkowski, M.; Zoulim, F.; Sherman, K.; Alberti, A.; Wei, L.; van Baelen, B.; Sullivan, J.; Kieffer, T.; de Meyer, S.; et al. Long-term follow-up of patients with chronic hepatitis C treated with telaprevir in combination with peginterferon alfa-2a and rabivirin: Interim analysis of extend study. Hepatology 2010, 52, 436A. [Google Scholar]
- Thomas, X.V.; de Bruijne, J.; Sullivan, J.C.; Kieffer, T.L.; Ho, C.K.; Rebers, S.P.; de Vries, M.; Reesink, H.W.; Weegink, C.J.; Molenkamp, R.; et al. Evaluation of persistence of resistant variants with ultra-deep pyrosequencing in chronic hepatitis C patients treated with telaprevir. PLoS ONE 2012, 7, e41191. [Google Scholar] [CrossRef] [PubMed]
- Lenz, O.; Verbinnen, T.; Fevery, B.; Tambuyzer, L.; Vijgen, L.; Peeters, M.; Buelens, A.; Ceulemans, H.; Beumont, M.; Picchio, G.; et al. Virology analyses of HCV isolates from genotype 1-infected patients treated with simeprevir plus peginterferon/ribavirin in phase IIb/III studies. J. Hepatol. 2015, 62, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Bae, A.; Sun, S.C.; Qi, X.; Chen, X.; Ku, K.; Worth, A.; Wong, K.A.; Harris, J.; Miller, M.D.; Mo, H. Susceptibility of treatment-naive hepatitis C virus (HCV) clinical isolates to HCV protease inhibitors. Antimicrob. Agents Chemother. 2010, 54, 5288–5297. [Google Scholar] [CrossRef] [PubMed]
- McPhee, F.; Friborg, J.; Levine, S.; Chen, C.; Falk, P.; Yu, F.; Hernandez, D.; Lee, M.S.; Chaniewski, S.; Sheaffer, A.K.; et al. Resistance analysis of the hepatitis C virus NS3 protease inhibitor asunaprevir. Antimicrob. Agents Chemother. 2012, 56, 3670–3681. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.; Marcellin, P.; Poordad, F.; de Araujo, E.S.; Buti, M.; Horsmans, Y.; Janczewska, E.; Villamil, F.; Scott, J.; Peeters, M.; et al. Simeprevir with pegylated interferon alfa 2a or 2b plus ribavirin in treatment-naive patients with chronic hepatitis C virus genotype 1 infection (QUEST-2): A randomised, double-blind, placebo-controlled phase 3 trial. Lancet (Lond. Engl.) 2014, 384, 414–426. [Google Scholar] [CrossRef]
- Lawitz, E.; Poordad, F.; Gutierrez, J.; Kakuda, T.; Picchio, G.; Beets, G.; Vandevoorde, A.; Peter, V.R.; Jaquesmyn, B.; Quinn, G.; et al. SVR 12 results from the phase II, open-label impact study of simeprevir (SMV) in combination with daclatasvir (DCV) and sofosbuvir (SOF) in treatment naive and experienced patients with chronic HCV genotype 1/4 infection and decompensated liver disease. Hepatology 2015, 62, 227A. [Google Scholar]
- Pawlotsky, J.M. NS5A inhibitors in the treatment of hepatitis C. J. Hepatol. 2013, 59, 375–382. [Google Scholar] [CrossRef] [PubMed]
- McGivern, D.R.; Masaki, T.; Williford, S.; Ingravallo, P.; Feng, Z.; Lahser, F.; Asante-Appiah, E.; Neddermann, P.; de Francesco, R.; Howe, A.Y.; et al. Kinetic analyses reveal potent and early blockade of hepatitis C virus assembly by NS5A inhibitors. Gastroenterology 2014, 147, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.; Romero-Brey, I.; Radujkovic, D.; Terreux, R.; Zayas, M.; Paul, D.; Harak, C.; Hoppe, S.; Gao, M.; Penin, F.; et al. Daclatasvir-like inhibitors of NS5A block early biogenesis of hepatitis C virus-induced membranous replication factories, independent of RNA replication. Gastroenterology 2014, 147, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Tellinghuisen, T.L.; Marcotrigiano, J.; Gorbalenya, A.E.; Rice, C.M. The NS5A protein of hepatitis C virus is a zinc metalloprotein. J. Biol. Chem. 2004, 279, 48576–48587. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, V.; Berger, K.L.; Kelly, S.M.; Thomas, M.; Deiters, A.; Randall, G. Daclatasvir inhibits hepatitis C virus NS5A motility and hyper-accumulation of phosphoinositides. Virology 2015, 476, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Reghellin, V.; Donnici, L.; Fenu, S.; Berno, V.; Calabrese, V.; Pagani, M.; Abrignani, S.; Peri, F.; de Francesco, R.; Neddermann, P. NS5A inhibitors impair NS5A-phosphatidylinositol 4-kinase iiialpha complex formation and cause a decrease of phosphatidylinositol 4-phosphate and cholesterol levels in hepatitis C virus-associated membranes. Antimicrob. Agents Chemother. 2014, 58, 7128–7140. [Google Scholar] [CrossRef] [PubMed]
- Sulkowski, M.S.; Gardiner, D.F.; Rodriguez-Torres, M.; Reddy, K.R.; Hassanein, T.; Jacobson, I.; Lawitz, E.; Lok, A.S.; Hinestrosa, F.; Thuluvath, P.J.; et al. Daclatasvir plus sofosbuvir for previously treated or untreated chronic HCV infection. N. Engl. J. Med. 2014, 370, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Fridell, R.A.; Wang, C.; Sun, J.H.; O’Boyle, D.R., 2nd; Nower, P.; Valera, L.; Qiu, D.; Roberts, S.; Huang, X.; Kienzle, B.; et al. Genotypic and phenotypic analysis of variants resistant to hepatitis C virus nonstructural protein 5A replication complex inhibitor BMS-790052 in humans: In vitro and in vivo correlations. Hepatology 2011, 54, 1924–1935. [Google Scholar] [PubMed]
- Dore, G.J.; Lawitz, E.; Hezode, C.; Shafran, S.; Ramji, A.; Tatum, H.; Taliani, G.; Tran, A.; Brunetto, M.; Zaltron, S.; et al. Daclatasvir combined with peginterferon alfa-2a and ribavirin for 12 or 16 weeks in patients with HCV genotype 1 or 3 infection: Command GT2/3 study. J. Hepatol. 2013, 58, S570–S571. [Google Scholar] [CrossRef]
- McPhee, F.; Hernandez, D.; Yu, F.; Ueland, J.; Monikowski, A.; Carifa, A.; Falk, P.; Wang, C.; Fridell, R.; Eley, T.; et al. Resistance analysis of hepatitis C virus genotype 1 prior treatment null responders receiving daclatasvir and asunaprevir. Hepatology 2013, 58, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Zeuzem, S.; Mizokami, M.; Pianko, S.; Mangia, A.; Han, K.; Martin, R.; Svarovskaia, E.; Dvory-Sobol, H.; Doehle, B.; Pang, P.; et al. Prevalence of pre-treatment NS5A resistance associated variants in genotype 1 patients across different regions using deep sequencing and effect on treatment outcome with LDV/SOF. Hepatology 2015, 62, 254A. [Google Scholar]
- Peiffer, K.H.; Sommer, L.; Susser, S.; Vermehren, J.; Herrmann, E.; Doring, M.; Dietz, J.; Perner, D.; Berkowski, C.; Zeuzem, S.; et al. IFN λ 4 genotypes and resistance-associated variants in HCV genotype 1 and 3 infected patients. Hepatology 2015. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Cooper, J.N.; Lalezari, J.P.; Lawitz, E.; Pockros, P.J.; Gitlin, N.; Freilich, B.F.; Younes, Z.H.; Harlan, W.; Ghalib, R.; et al. All-oral 12-week treatment with daclatasvir plus sofosbuvir in patients with hepatitis C virus genotype 3 infection: Ally-3 phase III study. Hepatology 2015, 61, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Pawlotsky, J.M. Therapy: Avoiding treatment failures associated with HCV resistance. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 673–674. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.H.; O’Boyle, D.R., 2nd; Fridell, R.A.; Langley, D.R.; Wang, C.; Roberts, S.B.; Nower, P.; Johnson, B.M.; Moulin, F.; Nophsker, M.J.; et al. Resensitizing daclatasvir-resistant hepatitis C variants by allosteric modulation of NS5A. Nature 2015, 527, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Ranjith-Kumar, C.; Kao, C. Biochemical Activities of the HCV NS5B RNA-Dependent RNA Polymerase; Taylor & Francis: Wymondham, England, 2006; pp. 293–310. [Google Scholar]
- Lontok, E.; Harrington, P.; Howe, A.; Kieffer, T.; Lennerstrand, J.; Lenz, O.; McPhee, F.; Mo, H.; Parkin, N.; Pilot-Matias, T.; et al. Hepatitis C virus drug resistance-associated substitutions: State of the art summary. Hepatology 2015, 62, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- Lam, A.M.; Espiritu, C.; Bansal, S.; Micolochick Steuer, H.M.; Niu, C.; Zennou, V.; Keilman, M.; Zhu, Y.; Lan, S.; Otto, M.J.; et al. Genotype and subtype profiling of PSI-7977 as a nucleotide inhibitor of hepatitis C virus. Antimicrob. Agents Chemother. 2012, 56, 3359–3368. [Google Scholar] [CrossRef] [PubMed]
- Svarovskaia, E.S.; Dvory-Sobol, H.; Parkin, N.; Hebner, C.; Gontcharova, V.; Martin, R.; Ouyang, W.; Han, B.; Xu, S.; Ku, K.; et al. Infrequent development of resistance in genotype 1–6 hepatitis C virus-infected subjects treated with sofosbuvir in phase 2 and 3 clinical trials. Clin. Infect. Dis. 2014, 59, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- Gane, E.J.; Stedman, C.A.; Hyland, R.H.; Ding, X.; Svarovskaia, E.; Symonds, W.T.; Hindes, R.G.; Berrey, M.M. Nucleotide polymerase inhibitor sofosbuvir plus ribavirin for hepatitis C. N. Engl. J. Med. 2013, 368, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Osinusi, A.; Meissner, E.G.; Lee, Y.J.; Bon, D.; Heytens, L.; Nelson, A.; Sneller, M.; Kohli, A.; Barrett, L.; Proschan, M.; et al. Sofosbuvir and ribavirin for hepatitis C genotype 1 in patients with unfavorable treatment characteristics: A randomized clinical trial. JAMA 2013, 310, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Gane, E.; Abergel, A.; Metivier, S.; Nahass, R.; Ryan, M.; Stedman, C.A.; Svarovskaia, E.; M, H.; Doehle, B.; Dvory-Sobol, H.; et al. The emergence of NS5B resistant associated variant S282T after sofosbuvir based treatment. Hepatology 2015, 62, 322 A. [Google Scholar]
- Gerber, L.; Welzel, T.M.; Zeuzem, S. New therapeutic strategies in HCV: Polymerase inhibitors. Liver Int.: Off. J. Int. Assoc. Study Liver 2013, 33, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Kati, W.; Koev, G.; Irvin, M.; Beyer, J.; Liu, Y.; Krishnan, P.; Reisch, T.; Mondal, R.; Wagner, R.; Molla, A.; et al. In vitro activity and resistance profile of dasabuvir, a nonnucleoside hepatitis C virus polymerase inhibitor. Antimicrob. Agents Chemother. 2015, 59, 1505–1511. [Google Scholar] [CrossRef] [PubMed]
- Kowdley, K.V.; Lawitz, E.; Poordad, F.; Cohen, D.E.; Nelson, D.R.; Zeuzem, S.; Everson, G.T.; Kwo, P.; Foster, G.R.; Sulkowski, M.S.; et al. Phase 2b trial of interferon-free therapy for hepatitis C virus genotype 1. N. Engl. J. Med. 2014, 370, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Feld, J.J.; Kowdley, K.V.; Coakley, E.; Sigal, S.; Nelson, D.R.; Crawford, D.; Weiland, O.; Aguilar, H.; Xiong, J.; Pilot-Matias, T.; et al. Treatment of HCV with ABT-450/r-ombitasvir and dasabuvir with ribavirin. N. Engl. J. Med. 2014, 370, 1594–1603. [Google Scholar] [CrossRef] [PubMed]
- Zeuzem, S.; Jacobson, I.M.; Baykal, T.; Marinho, R.T.; Poordad, F.; Bourliere, M.; Sulkowski, M.S.; Wedemeyer, H.; Tam, E.; Desmond, P.; et al. Retreatment of HCV with ABT-450/r-ombitasvir and dasabuvir with ribavirin. N. Engl. J. Med. 2014, 370, 1604–1614. [Google Scholar] [CrossRef] [PubMed]
- Ferenci, P.; Bernstein, D.; Lalezari, J.; Cohen, D.; Luo, Y.; Cooper, C.; Tam, E.; Marinho, R.T.; Tsai, N.; Nyberg, A.; et al. ABT-450/r-ombitasvir and dasabuvir with or without ribavirin for HCV. N. Engl. J. Med. 2014, 370, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Andreone, P.; Colombo, M.G.; Enejosa, J.V.; Koksal, I.; Ferenci, P.; Maieron, A.; Mullhaupt, B.; Horsmans, Y.; Weiland, O.; Reesink, H.W.; et al. ABT-450, ritonavir, ombitasvir, and dasabuvir achieves 97% and 100% sustained virologic response with or without ribavirin in treatment-experienced patients with HCV genotype 1b infection. Gastroenterology 2014, 147, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Fevery, B.; Thys, K.; Van Rossem, E.; Verbinnen, T.; Picchio, G.; Aerssens, J.; De Meyer, S.; Beumont, M.; Kerland, D.; Lenz, O. Deep Sequencing Analysis in HCV Genotype 1 Infected Patients Treated with Simeprevir Plus Sofosbuvir with/withour Ribavirin in the Cosmos Study; EASL: Vienna, Austria, 2015; p. P0780. [Google Scholar]
- Sarrazin, C.; Kieffer, T.L.; Bartels, D.; Hanzelka, B.; Muh, U.; Welker, M.; Wincheringer, D.; Zhou, Y.; Chu, H.M.; Lin, C.; et al. Dynamic hepatitis C virus genotypic and phenotypic changes in patients treated with the protease inhibitor telaprevir. Gastroenterology 2007, 132, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Susser, S.; Welsch, C.; Wang, Y.; Zettler, M.; Domingues, F.S.; Karey, U.; Hughes, E.; Ralston, R.; Tong, X.; Herrmann, E.; et al. Characterization of resistance to the protease inhibitor boceprevir in hepatitis C virus-infected patients. Hepatology 2009, 50, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- Susser, S.; Flinders, M.; Reesink, H.W.; Zeuzem, S.; Lawyer, G.; Ghys, A.; van Eygen, V.; Witek, J.; de Meyer, S.; Sarrazin, C. Evolution of hepatitis C virus quasispecies during repeated treatment with the ns3/4a protease inhibitor telaprevir. Antimicrob. Agents Chemother. 2015, 59, 2746–2755. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Abdelrahman, T.; Hughes, J.; Main, J.; McLauchlan, J.; Thursz, M.; Thomson, E. Next-generation sequencing sheds light on the natural history of hepatitis C infection in patients who fail treatment. Hepatology 2015, 61, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Ogishi, M.; Yotsuyanagi, H.; Tsutsumi, T.; Gatanaga, H.; Ode, H.; Sugiura, W.; Moriya, K.; Oka, S.; Kimura, S.; Koike, K. Deconvoluting the composition of low-frequency hepatitis C viral quasispecies: Comparison of genotypes and NS3 resistance-associated variants between HCV/HIV coinfected hemophiliacs and HCV monoinfected patients in Japan. PLoS ONE 2015, 10, e0119145. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, C.; Lathouwers, E.; Peeters, M.; Daems, B.; Buelens, A.; Witek, J.; Wyckmans, Y.; Fevery, B.; Verbinnen, T.; Ghys, A.; et al. Prevalence of the hepatitis C virus NS3 polymorphism Q80K in genotype 1 patients in the european region. Antivir. Res. 2015, 116, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Costantino, A.; Spada, E.; Equestre, M.; Bruni, R.; Tritarelli, E.; Coppola, N.; Sagnelli, C.; Sagnelli, E.; Ciccaglione, A.R. Naturally occurring mutations associated with resistance to HCV NS5B polymerase and ns3 protease inhibitors in treatment-naive patients with chronic hepatitis C. Virol. J. 2015, 12. [Google Scholar] [CrossRef] [PubMed]
- Margeridon-Thermet, S.; le Pogam, S.; Li, L.; Liu, T.F.; Shulman, N.; Shafer, R.W.; Najera, I. Similar prevalence of low-abundance drug-resistant variants in treatment-naive patients with genotype 1a and 1b hepatitis C virus infections as determined by ultradeep pyrosequencing. PLoS ONE 2014, 9, e105569. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, E.F.; Harrington, P.R.; O’Rear, J.J.; Naeger, L.K. Clinical evidence and bioinformatics characterization of potential hepatitis C virus resistance pathways for sofosbuvir. Hepatology 2015, 61, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, C. The importance of resistance to direct antiviral drugs in HCV infection in clinical practice. J. Hepatol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sun, J.H.; O’Boyle, D.R., 2nd; Nower, P.; Valera, L.; Roberts, S.; Fridell, R.A.; Gao, M. Persistence of resistant variants in hepatitis C virus-infected patients treated with the NS5A replication complex inhibitor daclatasvir. Antimicrob. Agents Chemother. 2013, 57, 2054–2065. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.; Queiroz, A.T.; Pessoa, M.G.; da Silva, E.F.; Mazo, D.F.; Carrilho, F.J.; Carvalho-Filho, R.J.; de Carvalho, I.M. The presence of resistance mutations to protease and polymerase inhibitors in hepatitis C virus sequences from the Los Alamos databank. J. Viral Hepat. 2013, 20, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, T.L.; George, S. Resistance to hepatitis C virus protease inhibitors. Curr. Opin. Virol. 2014, 8, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.D.; Sarrazin, C. Antiviral therapy of hepatitis C in 2014: Do we need resistance testing? Antivir. Res. 2014, 105, 64–71. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, A.; Felmlee, D.J. Mechanisms of Hepatitis C Viral Resistance to Direct Acting Antivirals. Viruses 2015, 7, 6716-6729. https://doi.org/10.3390/v7122968
Ahmed A, Felmlee DJ. Mechanisms of Hepatitis C Viral Resistance to Direct Acting Antivirals. Viruses. 2015; 7(12):6716-6729. https://doi.org/10.3390/v7122968
Chicago/Turabian StyleAhmed, Asma, and Daniel J. Felmlee. 2015. "Mechanisms of Hepatitis C Viral Resistance to Direct Acting Antivirals" Viruses 7, no. 12: 6716-6729. https://doi.org/10.3390/v7122968
APA StyleAhmed, A., & Felmlee, D. J. (2015). Mechanisms of Hepatitis C Viral Resistance to Direct Acting Antivirals. Viruses, 7(12), 6716-6729. https://doi.org/10.3390/v7122968