
The Association between Hantavirus Infection and Selenium Deficiency in Mainland China


Abstract
:1. Introduction
2. Results
Ecological Analysis
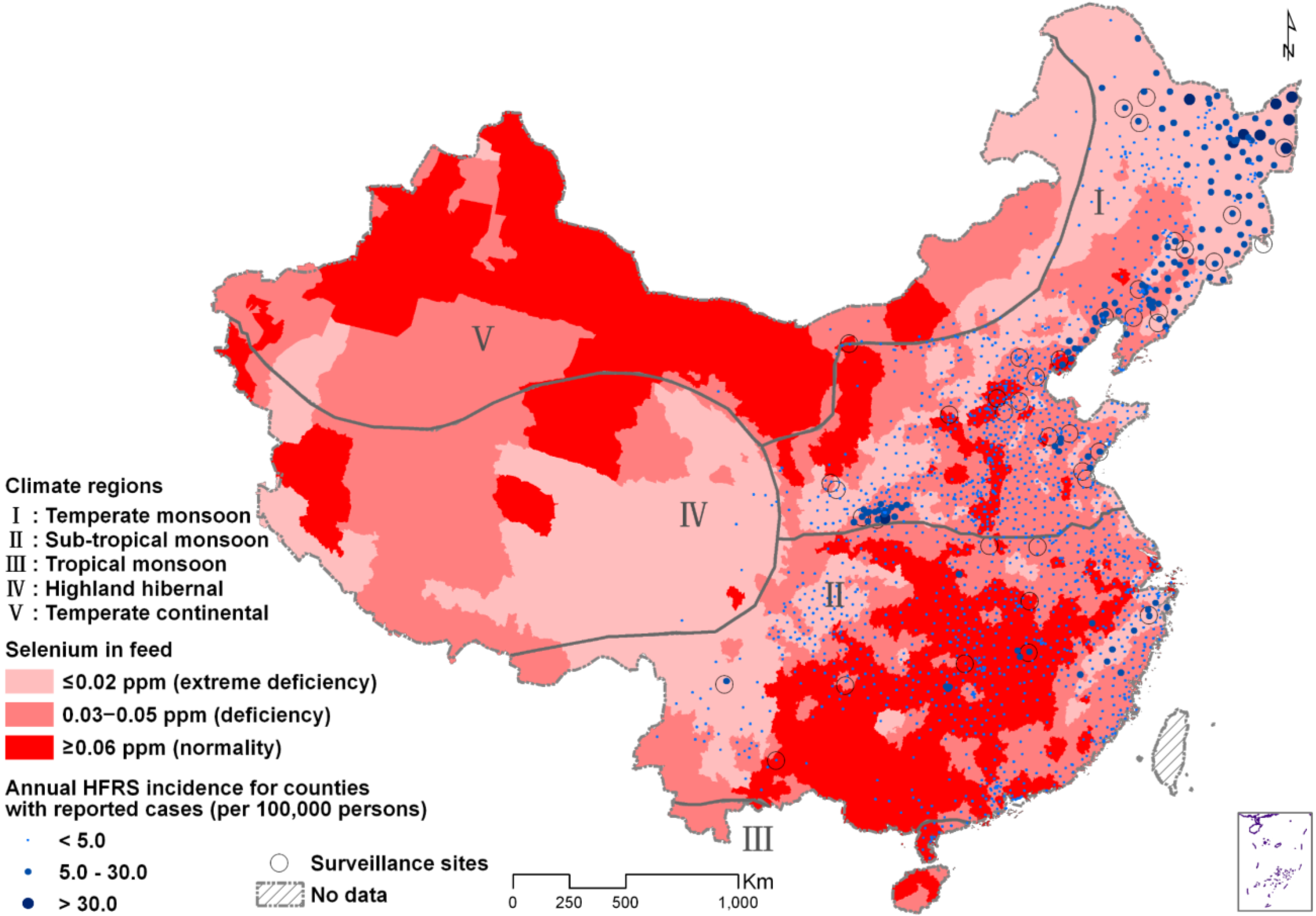

{kind=link}
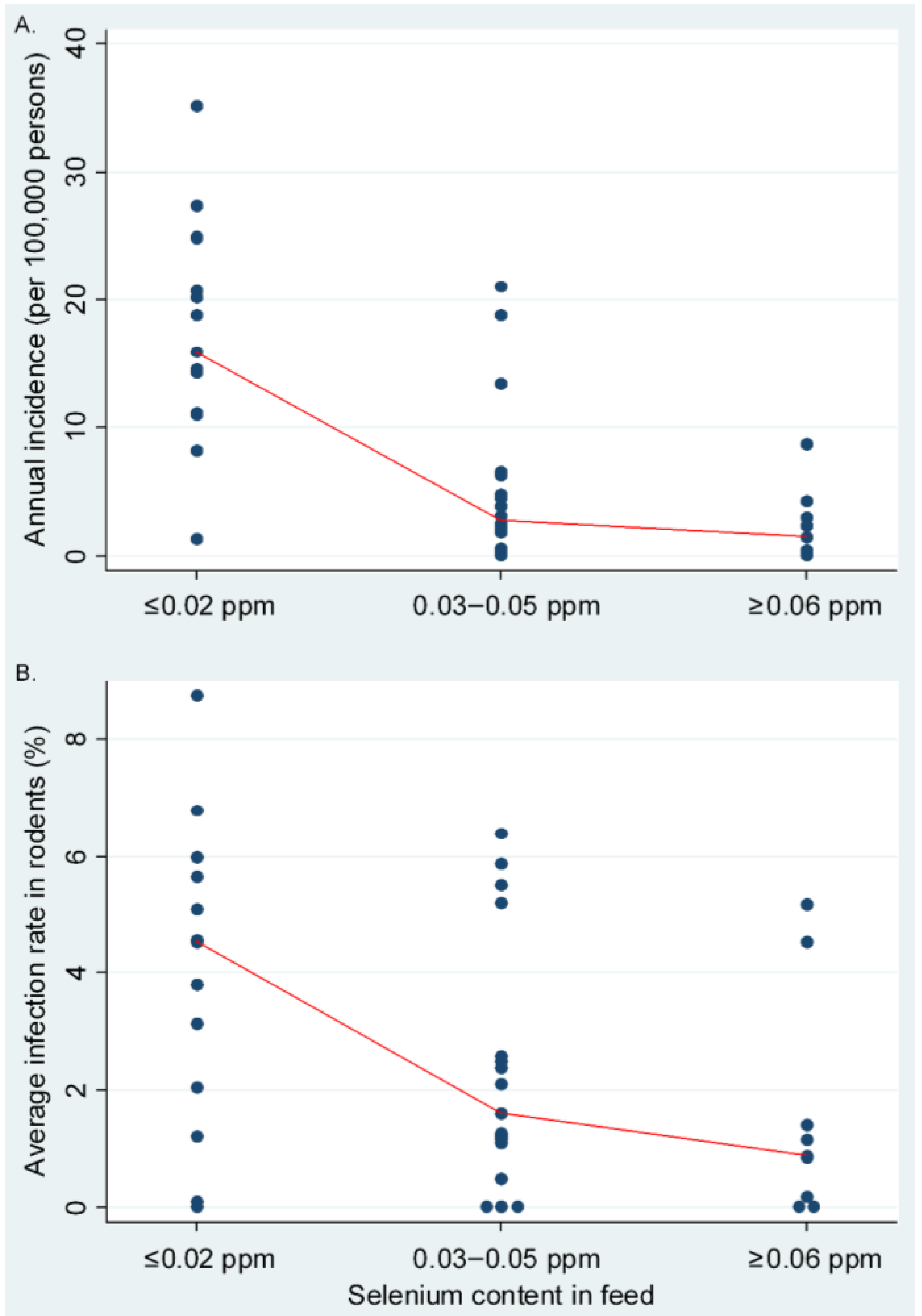{kind=link}
{kind=link}
{kind=link}
| Variables (Unit) a | Average yearly incidence (95% CI, per 100,000 person-years) | Univariate analysis | Multivariate analysis | ||
|---|---|---|---|---|---|
| Crude IRR (95% CI) | p-value | Adjusted IRR (95% CI) | p-value | ||
| Selenium content (categorical, ppm) | |||||
| ≥0.06 (non-deficient areas) | 0.40 (0.32–0.48) | 1 | - | 1 | - |
| 0.03–0.05 (moderate deficient areas) | 0.83 (0.70–0.95) | 2.07 (1.73–2.46) | <0.001 | 2.43 (2.04–2.88) | <0.001 |
| ≤0.02 (severe deficient areas) | 2.27 (1.85–2.70) | 5.68 (4.64–6.95) | <0.001 | 7.77 (6.29–9.61) | <0.001 |
| Elevation (categorical, 1000 m) | |||||
| <0.4 | 1.33 (1.14–1.52) | - | - | - | - |
| 0.4– | 1.37 (1.05–1.70) | - | - | - | - |
| 0.8– | 0.58 (0.35–0.81) | - | - | - | - |
| 1.6– | 0.17 (0.00–0.33) | - | - | - | - |
| >3.2 | 0.08 (0.00–0.16) | - | - | - | - |
| Elevation (continuous, 1000 m) | - | 0.46 (0.41–0.50) | <0.001 | 0.40 (0.35–0.46) | <0.001 |
| Croplands (categorical, %) | |||||
| <20 | 0.98 (0.76–1.20) | 1 | - | 1 | - |
| 20– | 1.03 (0.84–1.23) | 1.04 (0.86–1.26) | 0.659 | 1.14 (0.93–1.39) | 0.200 |
| >50 | 1.13 (0.91–1.35) | 1.14 (0.94–1.38) | 0.185 | 1.03 (0.84–1.27) | 0.766 |
| Forests (categorical, %) | |||||
| <5 | 0.90 (0.72–1.09) | 1 | - | 1 | - |
| 5– | 1.01 (0.81–1.21) | 1.11 (4.62–7.11) | 0.285 | 1.53 (1.26–1.87) | <0.001 |
| >40 | 1.21 (0.98–1.45) | 1.32 (1.42–2.27) | 0.004 | 1.53 (1.26–1.86) | <0.001 |
| Grasslands (categorical, %) | |||||
| <2 | 1.20 (1.02–1.38) | 1 | - | 1 | - |
| 2– | 1.41 (1.13–1.70) | 1.18 (0.98–1.41) | 0.082 | 0.92 (0.77–1.10) | 0.365 |
| >20 | 0.44 (0.32–0.55) | 0.38 (0.31–0.45) | <0.001 | 0.58 (0.45–0.74) | <0.001 |
| Population density (categorical, 1000 persons per km2) | |||||
| <0.2 | 1.38 (1.10–1.65) | - | - | - | - |
| 0.2– | 0.98 (0.82–1.14) | - | - | - | - |
| >0.5 | 0.73 (0.61–0.84) | - | - | - | - |
| Population density (continuous, 1000 persons per km2) | - | 0.97 (0.95–0.99) | 0.001 | 1.00 (0.98–1.02) | 0.728 |
| GDP (categorical, 10 million Yuan per km2) | |||||
| <0.08 | 1.22 (0.94–1.50) | - | - | - | - |
| 0.08– | 1.07 (0.90–1.25) | - | - | - | - |
| >0.4 | 0.84 (0.70–0.96) | - | - | - | - |
| GDP (continuous, 10 million Yuan per km2) | - | 0.91 (0.88–0.93) | <0.001 | 0.93 (0.90–0.96) | <0.001 |
| Variables (Unit) | Region I | Region II & Region III | ||
|---|---|---|---|---|
| Selenium content of crops and feed (categorical, ppm) | Adjusted IRR (95% CI) | p-value | Adjusted IRR (95% CI) | p-value |
| ≥0.06 (non-deficient areas) | 1 | - | 1 | - |
| 0.03–0.05 (moderate deficient areas) | 1.70 (1.27–2.27) | <0.001 | 0.85 (0.67–1.08) | 0.181 |
| ≤0.02 (severe deficient areas) | 3.45 (2.49–4.79) | <0.001 | 2.80 (1.92–4.79) | <0.001 |
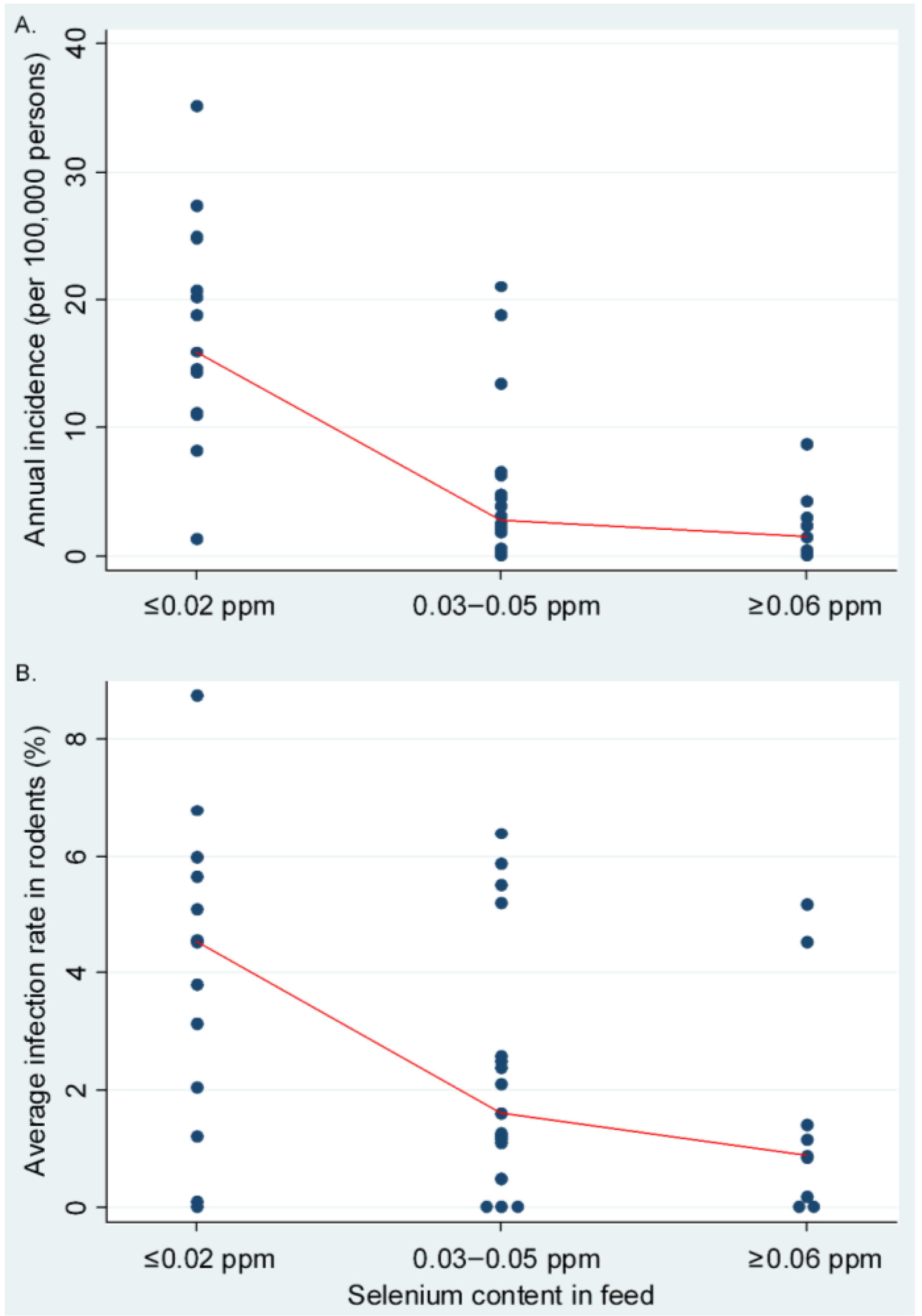
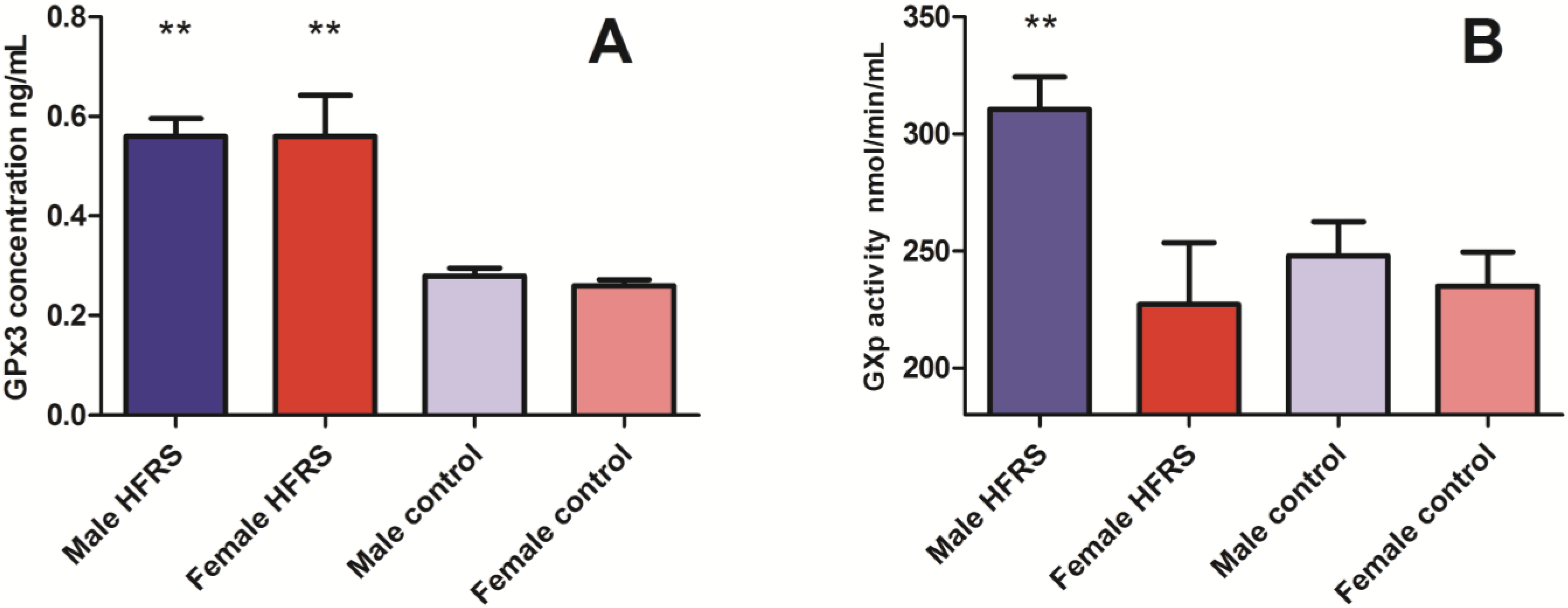
3. Clinical Analysis
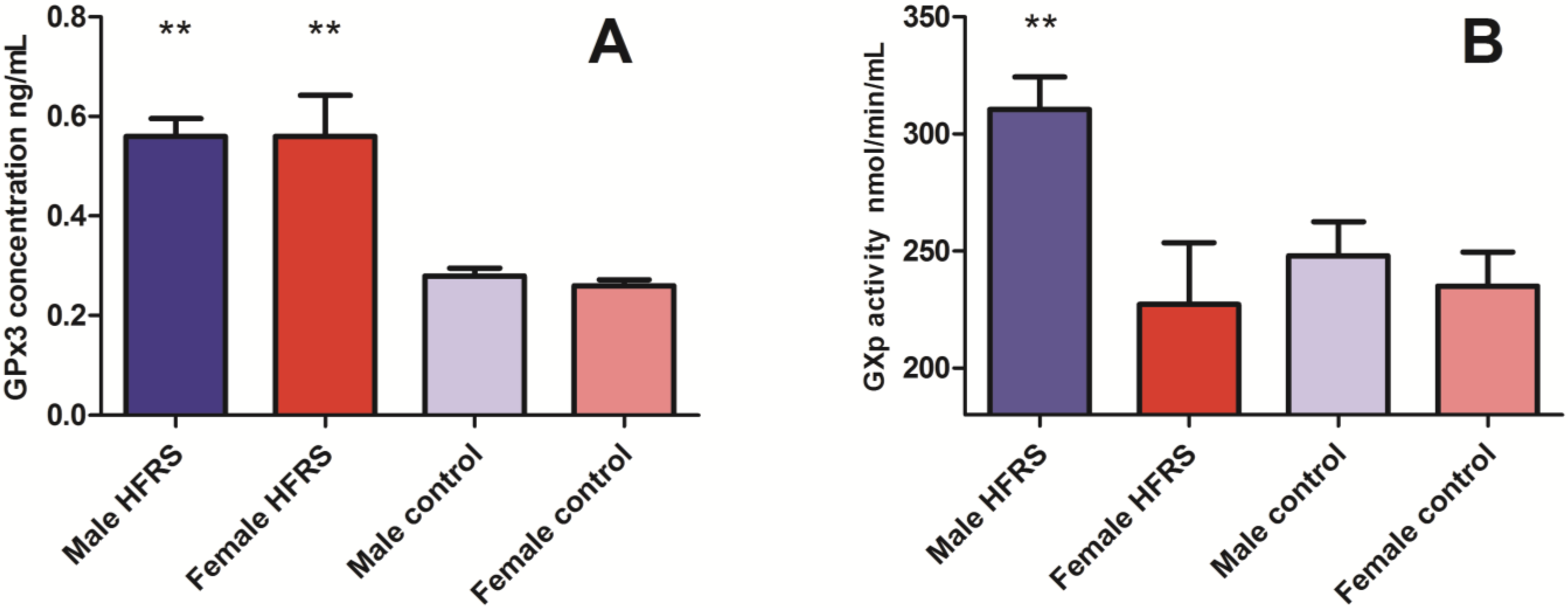
3.1. Glutathione Peroxidase (GPx3) Concentrations and GPx Activity in Acute HFRS Patients
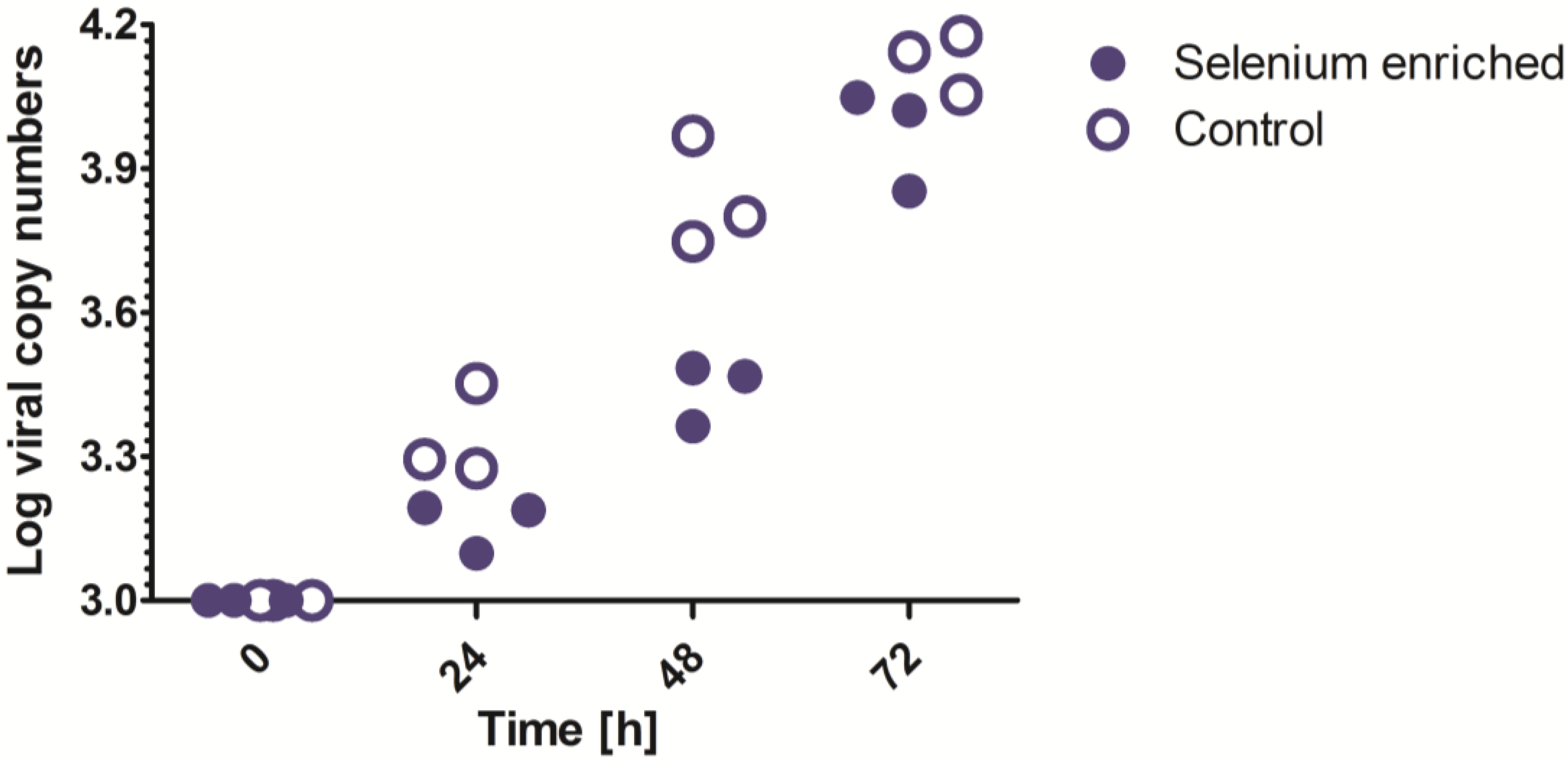
3.2. In Vitro Analysis of the Influence of Selenium on Hantavirus Replication
4. Discussion
5. Materials and Methods
5.1. Epidemiological Data Collection and Management
5.2. Spatial Analysis of HFRS Incidence
5.3. Ecological Analysis
5.4. Clinical Epidemiological Study
Selenium Levels in Acute HFRS Patients and Controls
5.5. In Vitro Assessment of the Influence of Selenium on Hantavirus Replication
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Goeijenbier, M.; Wagenaar, J.; Goris, M.; Martina, B.; Henttonen, H.; Vaheri, A.; Reusken, C.; Hartskeerl, R.; Osterhaus, A.; Van, G.E. Rodent-borne hemorrhagic fevers: Under-recognized, widely spread and preventable-epidemiology, diagnostics and treatment. Crit Rev. Microbiol. 2013, 39, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Vaheri, A.; Strandin, T.; Hepojoki, J.; Sironen, T.; Henttonen, H.; Makela, S.; Mustonen, J. Uncovering the mysteries of hantavirus infections. Nat. Rev. Microbiol. 2013, 11, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.C.; Sargianou, M.; Papa, A.; Chra, P.; Starakis, I.; Panos, G. Epidemiology of Hantavirus infections in humans: A comprehensive, global overview. Crit. Rev. Microbiol. 2014, 40, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Qiu, F.X.; Dong, B.J.; Ji, S.Z.; Li, Y.T.; Wang, Y.; Wang, H.M.; Zuo, G.F.; Tao, X.X.; Gao, S.Y. Epidemiological studies on hemorrhagic fever with renal syndrome in China. J. Infect. Dis. 1986, 154, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Song, G. Epidemiological progresses of hemorrhagic fever with renal syndrome in China. Chin. Med. J. (Engl.) 1999, 112, 472–477. [Google Scholar]
- Zhang, Y.Z.; Zou, Y.; Fu, Z.F.; Plyusnin, A. Hantavirus infections in humans and animals, China. Emerg. Infect. Dis. 2010, 16, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Yan, L.; Liang, S.; de Vlas, S.J.; Feng, D.; Han, X.; Zhao, W.; Xu, B.; Bian, L.; Yang, H.; et al. Spatial analysis of hemorrhagic fever with renal syndrome in China. BMC. Infect. Dis. 2006, 26, 6–77. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, C.B.; Figueiredo, L.T.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet. 2012, 379, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Molnar, J.; MacPherson, A.; Barclay, I.; Molnar, P. Selenium content of convenience and fast foods in Ayrshire, Scotland. Int. J. Food Sci. Nutr. 1995, 46, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wei, Z.; Tang, S.; Qi, Z. Distribution of selenium and cadmium in soil-rice system of selenium-rich area in Hainan, China. Pak. J. Pharm. Sci. 2014, 27, 1633–1639. [Google Scholar] [PubMed]
- De, T.L.; Waegeneers, N.; Thiry, C.; Du, L.G.; Tack, F.; Ruttens, A. Selenium content of Belgian cultivated soils and its uptake by field crops and vegetables. Sci. Total Environ. 2014, 468–469, 77–82. [Google Scholar] [PubMed]
- Stone, R. Diseases. A medical mystery in middle China. Science 2009, 324, 1378–1381. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, G.; Chen, J.; Chen, X.; Wen, Z.; Ge, K. Studies on the relations of selenium and Keshan disease. Biol. Trace Elem. Res. 1980, 2, 91–107. [Google Scholar] [CrossRef] [PubMed]
- Ge, K.; Yang, G. The epidemiology of selenium deficiency in the etiological study of endemic diseases in China. Am. J. Clin. Nutr. 1993, 57, 259S–263S. [Google Scholar] [PubMed]
- Whanger, P.D. Selenium and its relationship to cancer: An update. Br. J. Nutr. 2004, 91, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Rayman, M.P. The argument for increasing selenium intake. Proc. Nutr. Soc. 2002, 61, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.A.; Levander, O.A.; Handy, J. Selenium deficiency and viral infection. J. Nutr. 2003, 133, 1463S–1467S. [Google Scholar] [PubMed]
- Li, W.; Beck, M.A. Selenium deficiency induced an altered immune response and increased survival following influenza A/Puerto Rico/8/34 infection. Exp Biol. Med. (Maywood.) 2007, 232, 412–419. [Google Scholar]
- Beck, M.A.; Shi, Q.; Morris, V.C.; Levander, O.A. Rapid genomic evolution of a non-virulent coxsackievirus B3 in selenium-deficient mice results in selection of identical virulent isolates. Nat. Med. 1995, 1, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Levander, O.A.; Beck, M.A. Selenium and viral virulence. Br. Med. Bull. 1999, 55, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Kohrle, J. Pathophysiological relevance of selenium. J. Endocrinol. Invest. 2013, 36, 1–7. [Google Scholar] [PubMed]
- Curran, J.E.; Jowett, J.B.; Elliott, K.S.; Gao, Y.; Gluschenko, K.; Wang, J.; Abel Azim, D.M.; Cai, G.; Mahaney, M.C.; Comuzzie, A.G.; et al. Genetic variation in selenoprotein S influences inflammatory response. Nat. Genet. 2005, 37, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.A.; Handy, J.; Levander, O.A. Host nutritional status: The neglected virulence factor. Trends Microbiol. 2004, 12, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox. Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [PubMed]
- Carlson, B.A.; Yoo, M.H.; Shrimali, R.K.; Irons, R.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Role of selenium-containing proteins in T-cell and macrophage function. Proc. Nutr. Soc. 2010, 69, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, W.; Biestro, A.; Galusso, F.; Torre, M.H.; Manay, N.; Pittini, G.; Facchin, G.; Hardy, G. Serum selenium and glutathione peroxidase-3 activity: Biomarkers of systemic inflammation in the critically ill? Intensive Care Med. 2009, 35, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Goeijenbier, M.; Meijers, J.C.M.; Anfasa, F.; Roose, J.M.; van de Weg, C.A.M.; Bakhtiari, K.; Henttonen, H.; Vaheri, A.; Osterhaus, A.D.M.E.; van Gorp, E.C.M.; et al. Effect of Puumala hantavirus infection on Human Umbilical Vein Endothelial Cell hemostatic function: Platelet interactions, increased tissue factor expression and fibrinolysis regulator release. Front. Microb. submitted.
- Schmaljohn, C.; Hjelle, B. Hantaviruses: A global disease problem. Emerg. Infect. Dis. 1997, 3, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Zhu, W.; Wang, W.; Li, R.; Hou, S.; Wang, D.; Yang, L. Selenium in soil and endemic diseases in China. Sci. Total Environ. 2002, 284, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Alfthan, G.; Xu, G.L.; Tan, W.H.; Aro, A.; Wu, J.; Yang, Y.X.; Liang, W.S.; Xue, W.L.; Kong, L.H. Selenium supplementation of children in a selenium-deficient area in China: Blood selenium levels and glutathione peroxidase activities. Biol. Trace Elem. Res. 2000, 73, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.; Heyman, P. Factors driving hantavirus emergence in Europe. Curr. Opin. Virol. 2013, 3, 92–132. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Mengersen, K.; Bi, P.; Tong, S. Time-series analysis of the risk factors for haemorrhagic fever with renal syndrome: Comparison of statistical models. Epidemiol. Infect. 2007, 135, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.J.; McCormack, R.K.; Jonsson, C.B. Mathematical models for hantavirus infection in rodents. Bull. Math. Biol. 2006, 68, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Takata, Y.; Kristal, A.R.; Santella, R.M.; King, I.B.; Duggan, D.J.; Lampe, J.W.; Rayman, M.P.; Blount, P.L.; Reid, B.J.; Vaughan, T.L.; et al. Selenium, selenoenzymes, oxidative stress and risk of neoplastic progression from Barrett’s esophagus: Results from biomarkers and genetic variants. PLoS. One 2012, 7, e38612. [Google Scholar] [CrossRef] [PubMed]
- Erkekoglu, P.; Asci, A.; Ceyhan, M.; Kizilgun, M.; Schweizer, U.; Atas, C.; Kara, A.; Kocer, G.B. Selenium levels, selenoenzyme activities and oxidant/antioxidant parameters in H1N1-infected children. Turk. J. Pediatr. 2013, 55, 271–282. [Google Scholar] [PubMed]
- Nelson, H.K.; Shi, Q.; Van, D.P.; Schiffrin, E.J.; Blum, S.; Barclay, D.; Levander, O.A.; Beck, M.A. Host nutritional selenium status as a driving force for influenza virus mutations. FASEB J. 2001, 15, 1846–1848. [Google Scholar] [PubMed]
- Beck, M.A.; Nelson, H.K.; Shi, Q.; Van, D.P.; Schiffrin, E.J.; Blum, S.; Barclay, D.; Levander, O.A. Selenium deficiency increases the pathology of an influenza virus infection. FASEB J. 2001, 15, 1481–1483. [Google Scholar] [PubMed]
- Safronetz, D.; Prescott, J.; Feldmann, F.; Haddock, E.; Rosenke, R.; Okumura, A.; Brining, D.; Dahlstrom, E.; Porcella, S.F.; Ebihara, H.; et al. Pathophysiology ffof hantavirus pulmonary syndrome in rhesus macaques. Proc. Natl. Acad. Sci. USA 2014, 111, 7114–7119. [Google Scholar] [CrossRef] [PubMed]
- Nemirov, K.; Lundkvist, A.; Vaheri, A.; Plyusnin, A. Adaptation of Puumala hantavirus to cell culture is associated with point mutations in the coding region of the L segment and in the noncoding regions of the S segment. J. Virol. 2003, 77, 8793–8800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Zhang, F.X.; Wang, J.B.; Zhao, Z.W.; Li, M.H.; Chen, H.X.; Zou, Y.; Plyusnin, A. Hantaviruses in rodents and humans, Inner Mongolia Autonomous Region, China. Emerg. Infect. Dis. 2009, 15, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, H.; Han, Y.H.; Wang, X.F.; Wang, S.W.; Yin, W.W.; Li, Q.; Xu, Z. Epidemiology and surveillance programs on hemorrhagic fever with renal syndrome in Mainland China, 2. Zhonghua Liu Xing Bing Xue Za Zhi 2010, 31, 675–680. [Google Scholar] [PubMed]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the etiologic agent of Korean Hemorrhagic fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Wang, G.; Duan, Y.Q.; Lu, Z.; Liu, J.X. A Distribution Map of Selenium Content of Feed in Mainland China; China Agricultural Science and Technology Press: Beijing, China, 1985. [Google Scholar]
- The National Geospatial-Intelligence Agency and the National Aeronautics and Space Administration of USA (2009). The shuttle radar topography mission. 30 Arc Second scene SRTM. Available online: http://www.landcover.org/data/srtm/ (Accesed on 16 November 2013).
- Ministry of Education China. Ref Type: Internet Communication Institute Textbook (2006); Geography; Beijing People’s Education Press: Beijing, China, 2006. [Google Scholar]
- Laupland, K.B.; Ross, T.; Gregson, D.B. Staphylococcus aureus bloodstream infections: Risk factors, outcomes, and the influence of methicillin resistance in Calgary, Canada, 2000–2006. J. Infect. Dis. 2008, 198, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.M.; Koraka, P.; Osterhaus, A.D.; Martina, B.E. Development of a strand-specific real-time qRT-PCR for the accurate detection and quantitation of West Nile virus RNA. J. Virol. Methods 2013, 194, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Johansson, P.; Yap, G.; Low, H.T.; Siew, C.C.; Kek, R.; Ng, L.C.; Bucht, G. Molecular characterization of two hantavirus strains from different rattus species in Singapore. Virol. J. 2010, 22, 7–15. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, L.-Q.; Goeijenbier, M.; Zuo, S.-Q.; Wang, L.-P.; Liang, S.; Klein, S.L.; Li, X.-L.; Liu, K.; Liang, L.; Gong, P.; et al. The Association between Hantavirus Infection and Selenium Deficiency in Mainland China. Viruses 2015, 7, 333-351. https://doi.org/10.3390/v7010333
Fang L-Q, Goeijenbier M, Zuo S-Q, Wang L-P, Liang S, Klein SL, Li X-L, Liu K, Liang L, Gong P, et al. The Association between Hantavirus Infection and Selenium Deficiency in Mainland China. Viruses. 2015; 7(1):333-351. https://doi.org/10.3390/v7010333
Chicago/Turabian StyleFang, Li-Qun, Marco Goeijenbier, Shu-Qing Zuo, Li-Ping Wang, Song Liang, Sabra L. Klein, Xin-Lou Li, Kun Liu, Lu Liang, Peng Gong, and et al. 2015. "The Association between Hantavirus Infection and Selenium Deficiency in Mainland China" Viruses 7, no. 1: 333-351. https://doi.org/10.3390/v7010333
APA StyleFang, L.-Q., Goeijenbier, M., Zuo, S.-Q., Wang, L.-P., Liang, S., Klein, S. L., Li, X.-L., Liu, K., Liang, L., Gong, P., Glass, G. E., Van Gorp, E., Richardus, J. H., Ma, J.-Q., Cao, W.-C., & De Vlas, S. J. (2015). The Association between Hantavirus Infection and Selenium Deficiency in Mainland China. Viruses, 7(1), 333-351. https://doi.org/10.3390/v7010333