Low-Frequency NNRTI-Resistant HIV-1 Variants and Relationship to Mutational Load in Antiretroviral-Naïve Subjects

Abstract
:1. Introduction
2. Materials and Methods
3. Statistical Methods
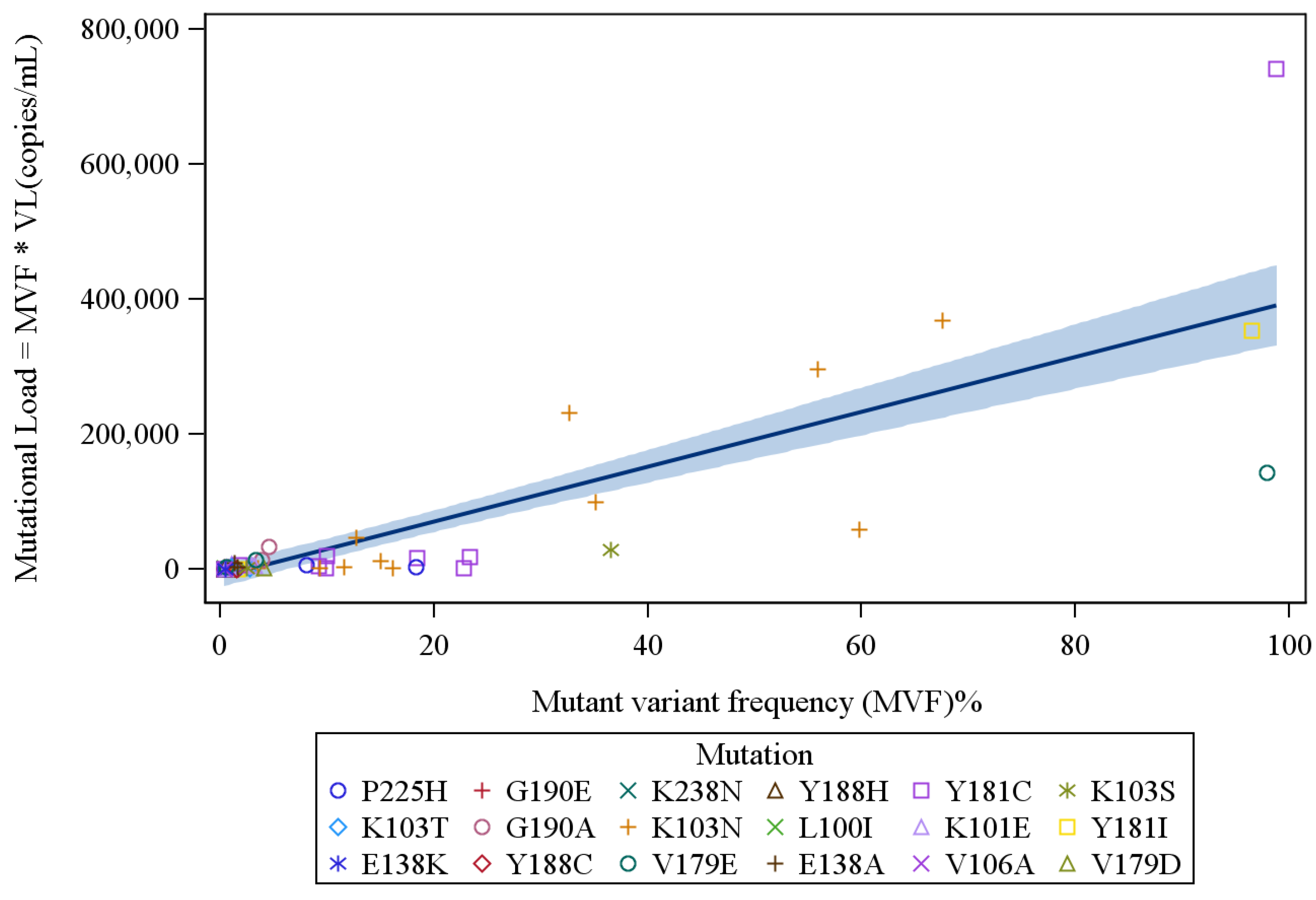
4. Results

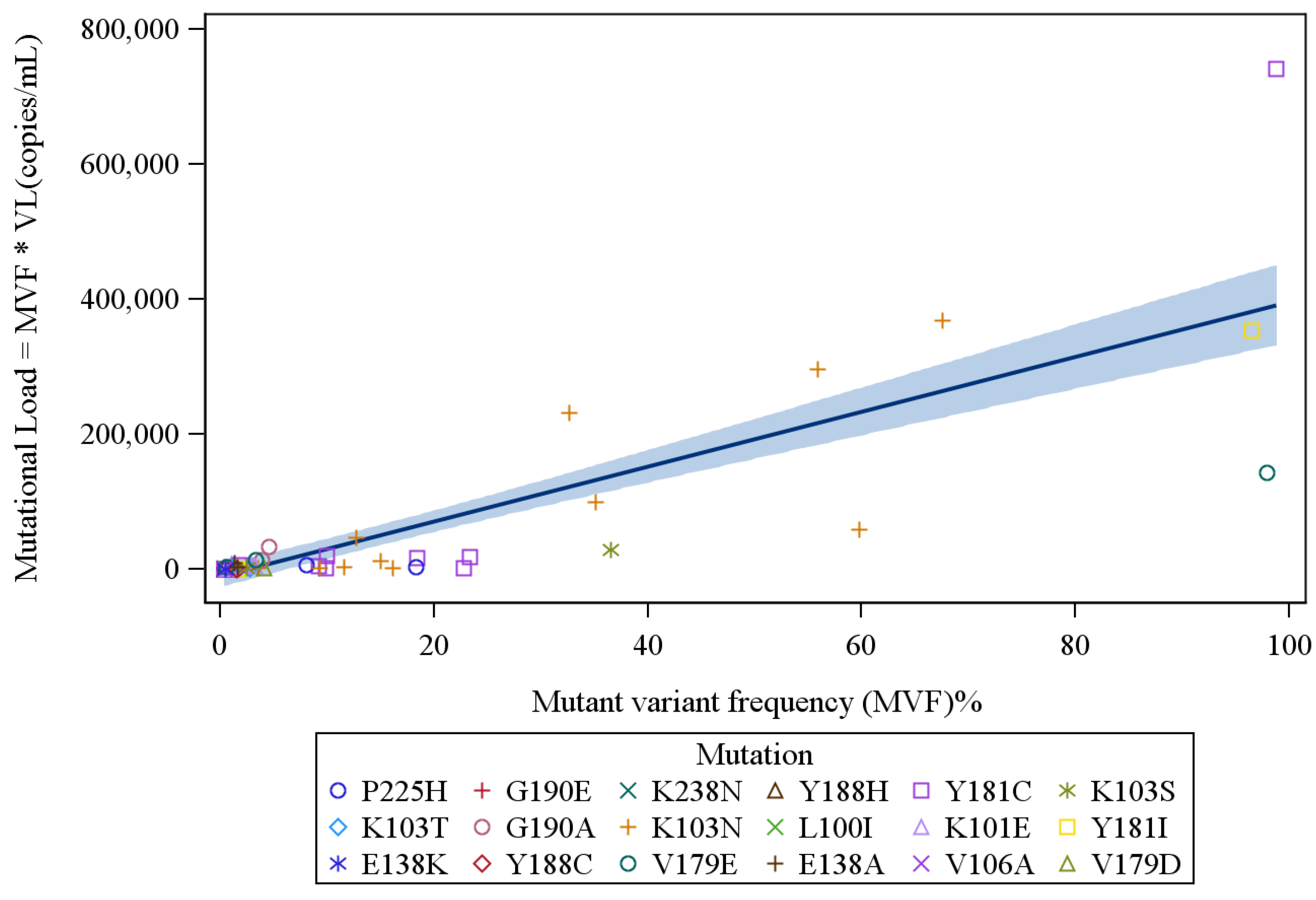{kind=link}
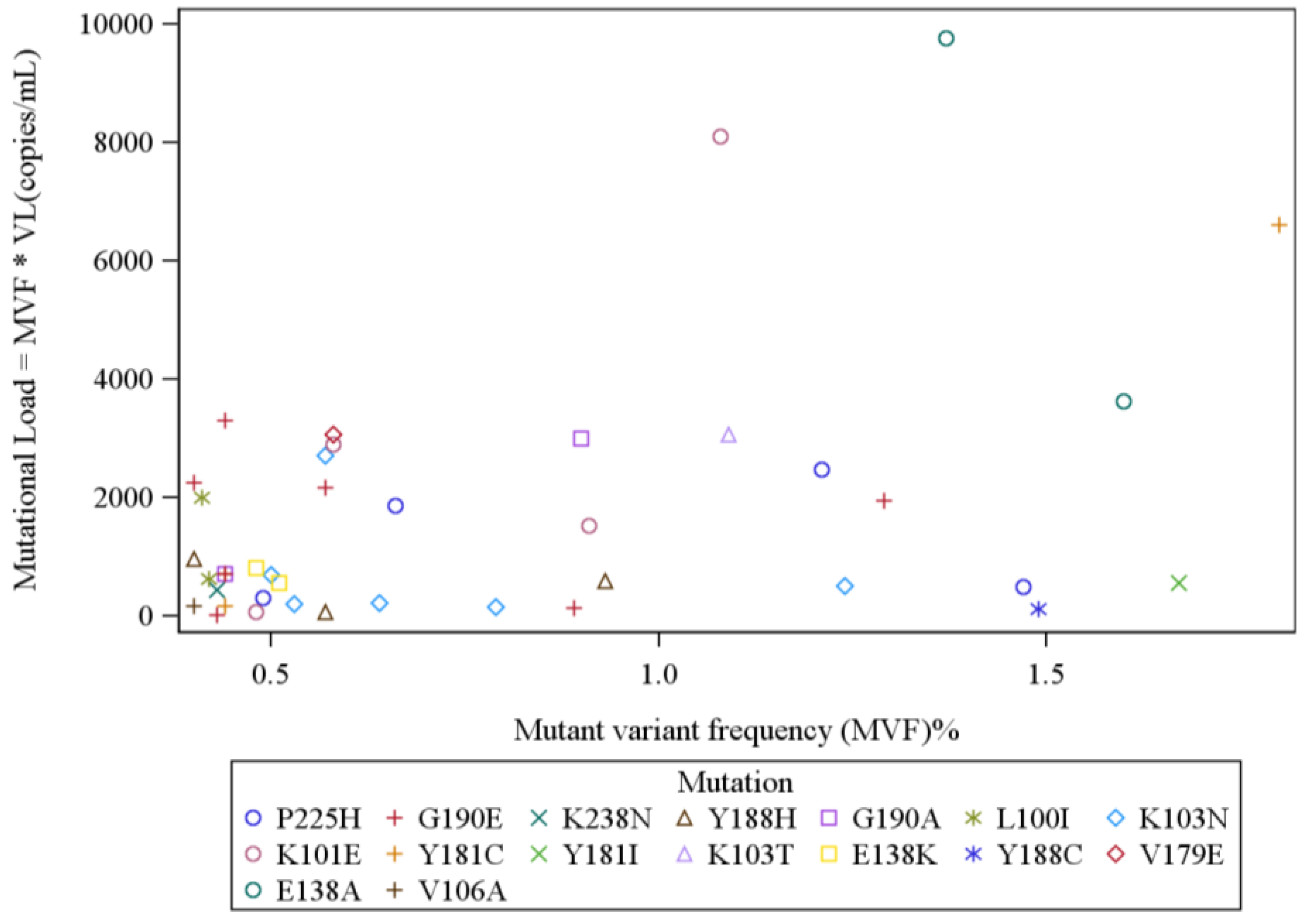{kind=link}
| Mutant Variant | Number Detected | Number of Patient Samples Tested | % Patient Samples with Mutant Variant | Range of Mutant Variant Frequency (%) | Range of Mutational Load (copies/mL) |
|---|---|---|---|---|---|
| L100I | 2 | 204 | 1.0 | 0.41–0.42 | 638–2005 |
| K101E | 6 | 204 | 2.9 | 0.48–3.28 | 76–8100 |
| K103N | 17 | 204 | 8.3 | 0.50–67.60 | 163–368,420 |
| K103S | 2 | 204 | 1.0 | 2.50–36.50 | 2483–28,835 |
| K103T | 2 | 204 | 1.0 | 1.09–2.79 | 2204–3074 |
| V106A | 1 | 204 | 0.5 | 0.40 | 182 |
| E138A * | 2 | 56 | 3.6 | 1.37–1.60 | 3632–9754 |
| E138K * | 2 | 56 | 3.6 | 0.48–0.51 | 571–811 |
| V179D * | 1 | 56 | 1.8 | 4.07 | 1400 |
| V179E * | 3 | 56 | 5.4 | 0.58–97.92 | 3074–142,963 |
| Y181C | 10 | 204 | 4.9 | 0.44–98.80 | 168–741,001 |
| Y181I | 2 | 204 | 1.0 | 1.67–96.46 | 558–354,008 |
| Y188C | 1 | 204 | 0.5 | 1.49 | 124 |
| Y188H | 3 | 204 | 1.5 | 0.40–0.93 | 75–972 |
| G190A | 4 | 204 | 2.0 | 0.44–4.57 | 717–34,275 |
| G190E | 7 | 204 | 3.4 | 0.40–1.29 | 25–3300 |
| P225H | 6 | 204 | 2.9 | 0.49–18.33 | 307–7124 |
| K238N | 1 | 204 | 0.5 | 0.43 | 439 |

| Subtype | Subjects from Each Subtype | Unique Subjects with NNRTI Mutations | NNRTI Mutations | Range of Frequency Prevalence ≥0.4% |
|---|---|---|---|---|
| A | 1 | 0 | ||
| AE | 7 | 2 | K101E, K103N, Y181C, P225H | 0.58–18.44 |
| B | 144 | 38 | L100I, K101E, K103N/S/T, E138A/K, V106A, V179D/E Y181C/I, P225H, G190E, K238N, Y188C/H, G190A/E, P225H | 0.4–97.92 |
| BF | 18 | 7 | K101E, K103N, Y181C, Y188H, G190E | 0.5–98.8 |
| C | 27 | 5 | K103N, Y181C, G190A P225H | 0.44–9.3 |
| CRFs | 5 | 0 | ||
| F | 2 | 0 | ||
| Total | 204 | 52 |
5. Discussion
Acknowledgments
Conflicts of Interest
References and Notes
- Simen, B.B.; Simons, J.F.; Hullsiek, K.H.; Novak, R.M.; Macarthur, R.D.; Baxter, J.D.; Huang, C.; Lubeski, C.; Turenchalk, G.S.; Braverman, M.S.; et al. Low-abundance drug-resistant viral variants in chronically HIV-infected, antiretroviral treatment-naive patients significantly impact treatment outcomes. J. Infect. Dis. 2009, 199, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Peuchant, O.; Thiébaut, R.; Capdepont, S.; Lavignolle-Aurillac, V.; Neau, D.; Morlat, P.; Dabis, F.; Fleury, H.; Masquelier, B.; ANRS CO3 Aquitaine Cohort. Transmission of HIV-1 minority-resistant variants and response to first-line antiretroviral therapy. AIDS 2008, 22, 1417–1423. [Google Scholar]
- Paredes, R.; Lalama, C.M.; Ribaudo, H.J.; Schackman, B.R.; Shikuma, C.; Giguel, F.; Meyer, W.A., 3rd; Johnson, V.A.; Fiscus, S.A.; D’Aquila, R.T.; AIDS Clinical Trials Group (ACTG) A5095 Study Team; et al. Preexisting minority drug-resistant HIV-1 variants, adherence, and risk of antiretroviral treatment failure. J. Infect. Dis. 2010, 201, 662–671. [Google Scholar]
- Jakobsen, M.R.; Tolstrup, M.; Søgaard, O.S.; Jørgensen, L.B.; Gorry, P.R.; Laursen, A.; Ostergaard, L. Transmission of HIV-1 drug-resistant variants: Prevalence and effect on treatment outcome. Clin. Infect. Dis. 2010, 50, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.A.; Li, J.F.; Wei, X.; Lipscomb, J.; Irlbeck, D.; Craig, C.; Smith, A.; Bennett, D.E.; Monsour, M.; Sandstrom, P.; et al. Minority HIV-1 drug resistance mutations are present in antiretroviral treatment-naïve populations and associated with reduced treatment efficacy. PLoS Med. 2008, 5, e158. [Google Scholar] [CrossRef] [PubMed]
- Metzner, K.J.; Rauch, P.; Braun, P.; Knechten, H.; Ehret, R.; Korn, K.; Kaiser, R.; Sichtig, N.; Ranneberg, B.; van Lunzen, J.; et al. Prevalence of key resistance mutations K65R, K103N, and M184Vas minority HIV-1 variants in chronically HIV-1 infected, treatment-naïve patients. J. Clin. Virol. 2011, 50, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Goodman, D.D.; Zhou, Y.; Margot, N.A.; McColl, D.J.; Zhong, L.; Borroto-Esoda, K.; Miller, M.D.; Svarovskaia, E.S. Low level of the K103N HIV-1 above a threshold is associated with virological failure in treatment-naive individuals undergoing efavirenz-containing therapy. AIDS 2011, 25, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Balduin, M.; Oette, M.; Däumer, M.P.; Hoffmann, D.; Pfister, H.J.; Kaiser, R. Prevalence of minor variants of HIV strains at reverse transcriptase position 103 in therapy-naive patients and their impact on the virological failure. J. Clin. Virol. 2009, 45, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Geretti, A.M.; Fox, Z.V.; Booth, C.L.; Smith, C.J.; Phillips, A.N.; Johnson, M.; Li, J.F.; Heneine, W.; Johnson, J.A. Low-frequency K103N strengthens the impact of transmitted drug resistance on virologic responses to first-line efavirenz or nevirapine-based highly active antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2009, 52, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Metzner, K.J.; Giulieri, S.G.; Knoepfel, S.A.; Rauch, P.; Burgisser, P.; Yerly, S.; Gunthard, H.F.; Cavassini, M. Minority quasispecies of drug-resistant HIV-1 that lead to early therapy failure in treatment-naive and -adherent patients. Clin. Infect. Dis. 2009, 48, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Kuritzkes, D.R.; Lalama, C.M.; Ribaudo, H.J.; Marcial, M.; Meyer, W.A., 3rd; Shikuma, C.; Johnson, V.A.; Fiscus, S.A.; D’Aquila, R.T.; Schackman, B.R.; et al. Preexisting resistance to nonnucleoside reverse-transcriptase inhibitors predicts virologic failure of an efavirenz-based regimen in treatment-naive HIV-1-infected subjects. J. Infect. Dis. 2008, 197, 867–870. [Google Scholar]
- Li, J.Z.; Paredes, R.; Ribaudo, H.J.; Svarovskaia, E.S.; Metzner, K.J.; Kozal, M.J.; Huppler Hullsiek, K.; Balduin, M.; Jakobsen, M.R.; Geretti, A.M.; et al. Minority HIV-1 Drug Resistance Mutations and the Risk of NNRTI-based Antiretroviral Treatment Failure: A Systematic Review and Pooled Analysis. JAMA 2011, 305, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Kozal, M.J.; Lupo, S.; DeJesus, E.; Molina, J.M.; McDonald, C.; Raffi, F.; Benetucci, J.; Mancini, M.; Yang, R.; Wirtz, V.; The Spartan Study Team; et al. A Nucleoside- and Ritonavir-Sparing Regimen Containing Atazanavir Plus Raltegravir in Antiretroviral Treatment-Naïve HIV-Infected Patients: SPARTAN Study Results. HIV Clin. Trials 2012, 13, 119–130. [Google Scholar]
- Lataillade, M.; Chiarella, J.; Yang, R.; Schnittman, S.; Wirtz, V.; Uy, J.; Seekins, D.; Krystal, M.; Mancini, M.; McGrath, D.; et al. Prevalence and clinical significance of HIV drug resistance mutations by ultra-deep sequencing in antiretroviral-naïve subjects in the CASTLE study. PLoS One 2010, 5, e10952. [Google Scholar]
- Kozal, M.J.; Chiarella, J.; St John, E.P.; Moreno, E.A.; Simen, B.B.; Arnold, T.E.; Lataillade, M. Prevalence of low-level HIV-1 variants with reverse transcriptase mutation K65R and the effect of antiretroviral drug exposure on variant levels. Antivir. Ther. 2011, 16, 925–929. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.Y.; Gonzales, M.J.; Kantor, R.; Betts, B.J.; Ravela, J.; Shafer, R.W. Human immunodeficiency virus reverse transcriptase and protease sequence database. Nucleic Acids Res. 2003, 31, 298–303. [Google Scholar] [CrossRef] [PubMed]
- SAS/STAT, version 9.3; SAS Institute Inc.: Cary, NC, 2011.
- Lataillade, M.; Chiarella, J.; Yang, R.; DeGrosky, M.; Uy, J.; Seekins, D.; Simen, B.; St. John, E.; Moreno, E.; Kozal, M. Virologic failures on initial boosted-PI regimen infrequently possess low-level variants with major PI resistance mutations by ultra-deep sequencing. PLoS One 2012, 7, e30118. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.J.; Molina, J.M.; Cahn, P.; Clotet, B.; Fourie, J.; Grinsztejn, B.; Wu, H.; Johnson, M.A.; Saag, M.; Supparatpinyo, K.; ECHO and THRIVE Study Groups; et al. Efficacy and Safety of Rilpivirine (TMC278) versus Efavirenz at 48 Weeks in Treatment-Naive HIV-1-Infected Patients: Pooled Results From the Phase 3 Double-Blind Randomized ECHO and THRIVE Trials. J. Acquir. Immune Defic. Syndr. 2012, 60, 33–42. [Google Scholar]
- Van Leth, F.; Andrews, S.; Grinsztejn, B.; Wilkins, E.; Lazanas, M.K.; Lange, J.M.; Montaner, J.; 2NN Study Group. The effect of baseline CD4 cell count and HIV-1 viral load on the efficacy and safety of nevirapine or efavirenz-based first-line HAART. AIDS 2005, 19, 463–471. [Google Scholar]
- Rimsky, L.; Vingerhoets, J.; van Eygen, V.; Eron, J.; Clotet, B.; Hoogstoel, A.; Boven, K.; Picchio, G. Genotypic and phenotypic characterization of HIV-1 isolates obtained from patients on rilpivirine therapy experiencing virologic failure in the phase 3 ECHO and THRIVE studies: 48-week analysis. J. Acquir. Immune Defic. Syndr. 2012, 59, 39–46. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gupta, S.; Lataillade, M.; Kyriakides, T.C.; Chiarella, J.; John, E.P.S.; Webb, S.; Moreno, E.A.; Simen, B.B.; Kozal, M.J. Low-Frequency NNRTI-Resistant HIV-1 Variants and Relationship to Mutational Load in Antiretroviral-Naïve Subjects. Viruses 2014, 6, 3428-3437. https://doi.org/10.3390/v6093428
Gupta S, Lataillade M, Kyriakides TC, Chiarella J, John EPS, Webb S, Moreno EA, Simen BB, Kozal MJ. Low-Frequency NNRTI-Resistant HIV-1 Variants and Relationship to Mutational Load in Antiretroviral-Naïve Subjects. Viruses. 2014; 6(9):3428-3437. https://doi.org/10.3390/v6093428
Chicago/Turabian StyleGupta, Shaili, Max Lataillade, Tassos C. Kyriakides, Jennifer Chiarella, Elizabeth P. St. John, Suzin Webb, Elizabeth A. Moreno, Birgitte B. Simen, and Michael J. Kozal. 2014. "Low-Frequency NNRTI-Resistant HIV-1 Variants and Relationship to Mutational Load in Antiretroviral-Naïve Subjects" Viruses 6, no. 9: 3428-3437. https://doi.org/10.3390/v6093428
APA StyleGupta, S., Lataillade, M., Kyriakides, T. C., Chiarella, J., John, E. P. S., Webb, S., Moreno, E. A., Simen, B. B., & Kozal, M. J. (2014). Low-Frequency NNRTI-Resistant HIV-1 Variants and Relationship to Mutational Load in Antiretroviral-Naïve Subjects. Viruses, 6(9), 3428-3437. https://doi.org/10.3390/v6093428