CPB1 of Aedes aegypti Interacts with DENV2 E Protein and Regulates Intracellular Viral Accumulation and Release from Midgut Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Cell Cultures
2.2. Plasmid Construction and Yeast Transfection
2.3. Aedes aegypti Mosquitoes
2.4. Aedes aegypti Primary Midgut Cell Preparations
2.5. cDNA Library Construction
2.6. Yeast Two-Hybrid Screening
2.7. Mammalian Two-Hybrid Analysis
2.8. Double Immunofluorescence Assay
2.9. Co-Immunoprecipitation Analysis
2.10. Molecular Docking Analysis
2.11. CPB1 Upregulation Study
3. Results
3.1. Ae. aegypti Midgut cDNA Library Construction
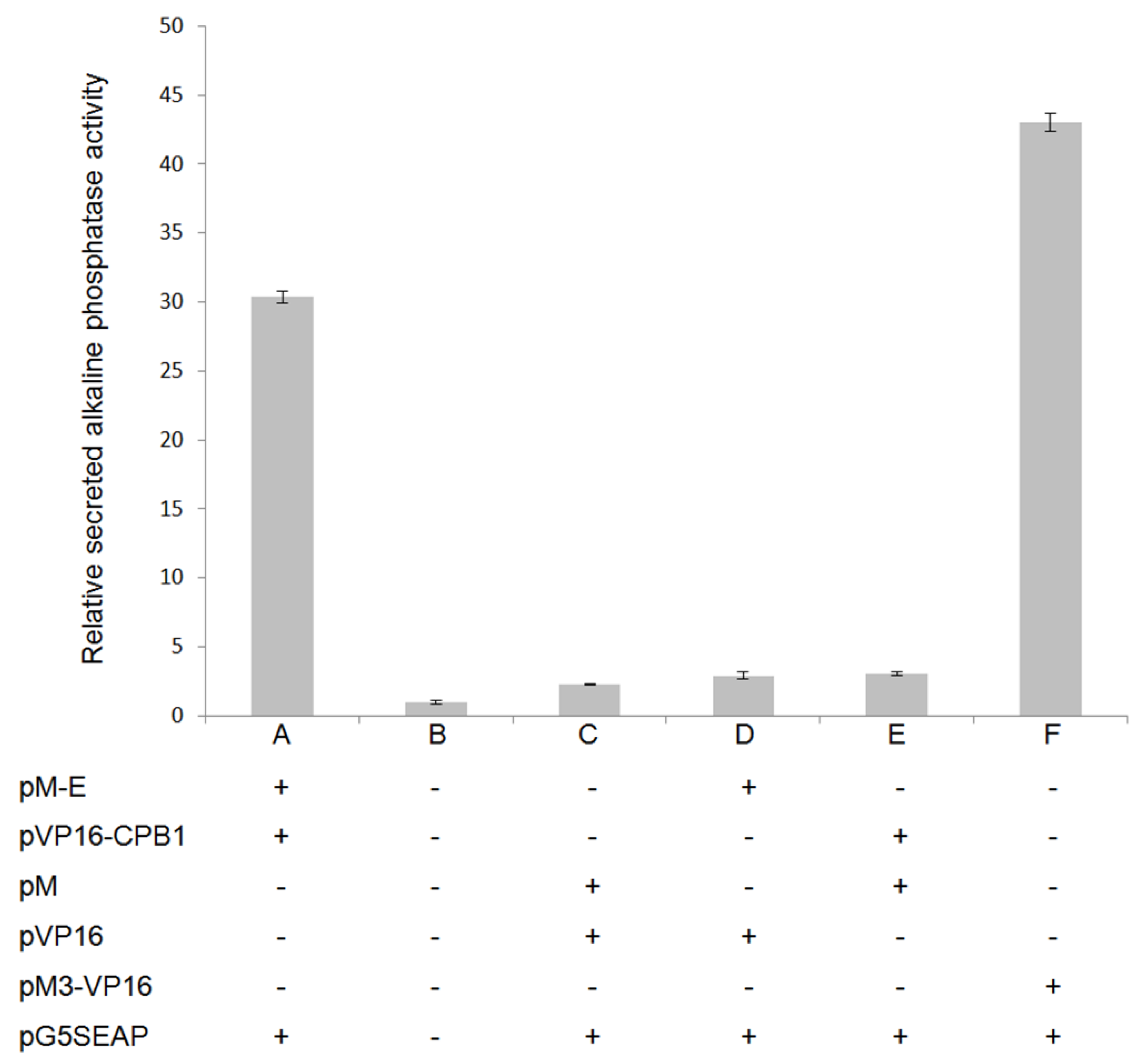
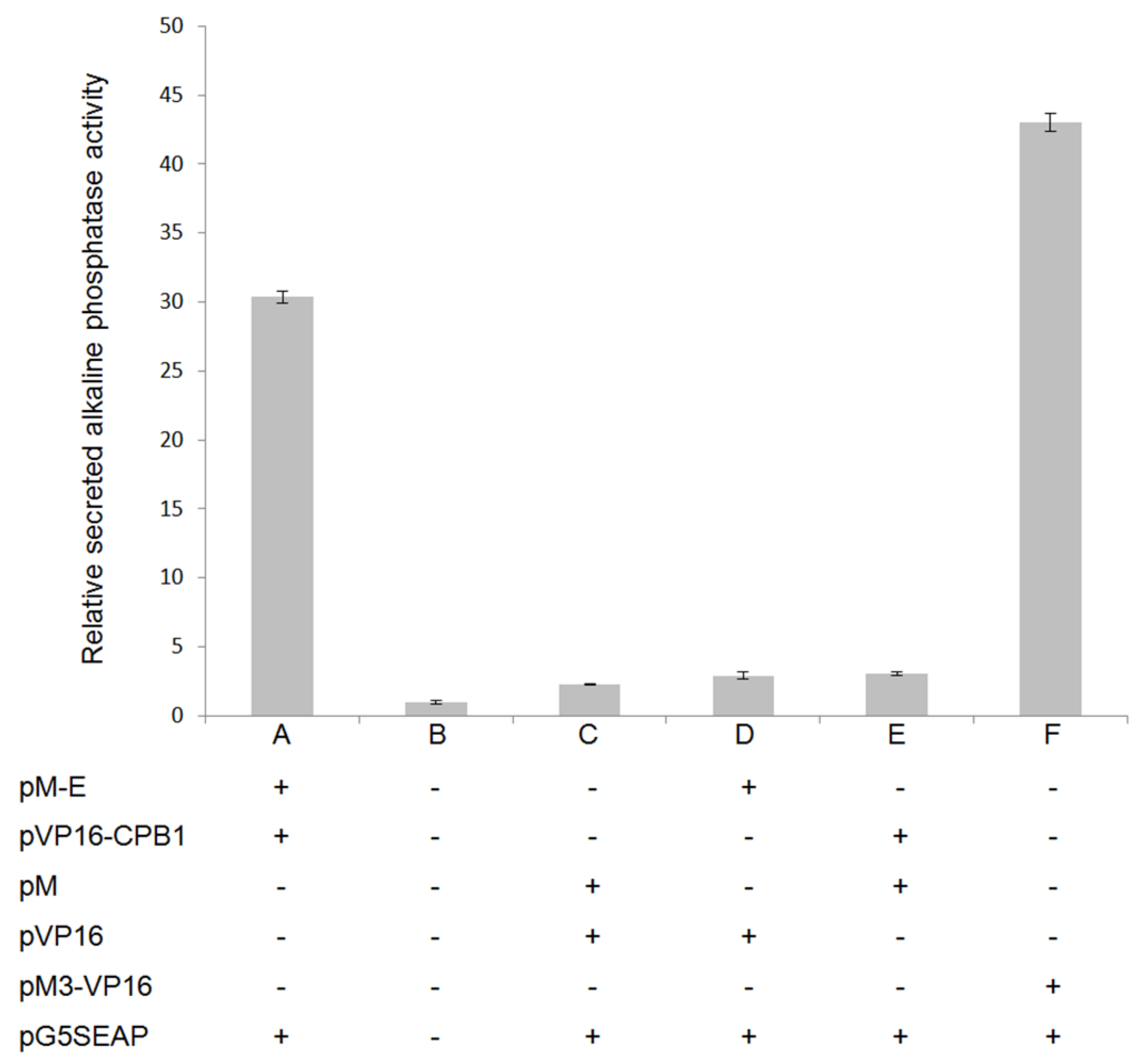
3.2. CPB1 Protein Interacts with the DENV2 E Protein
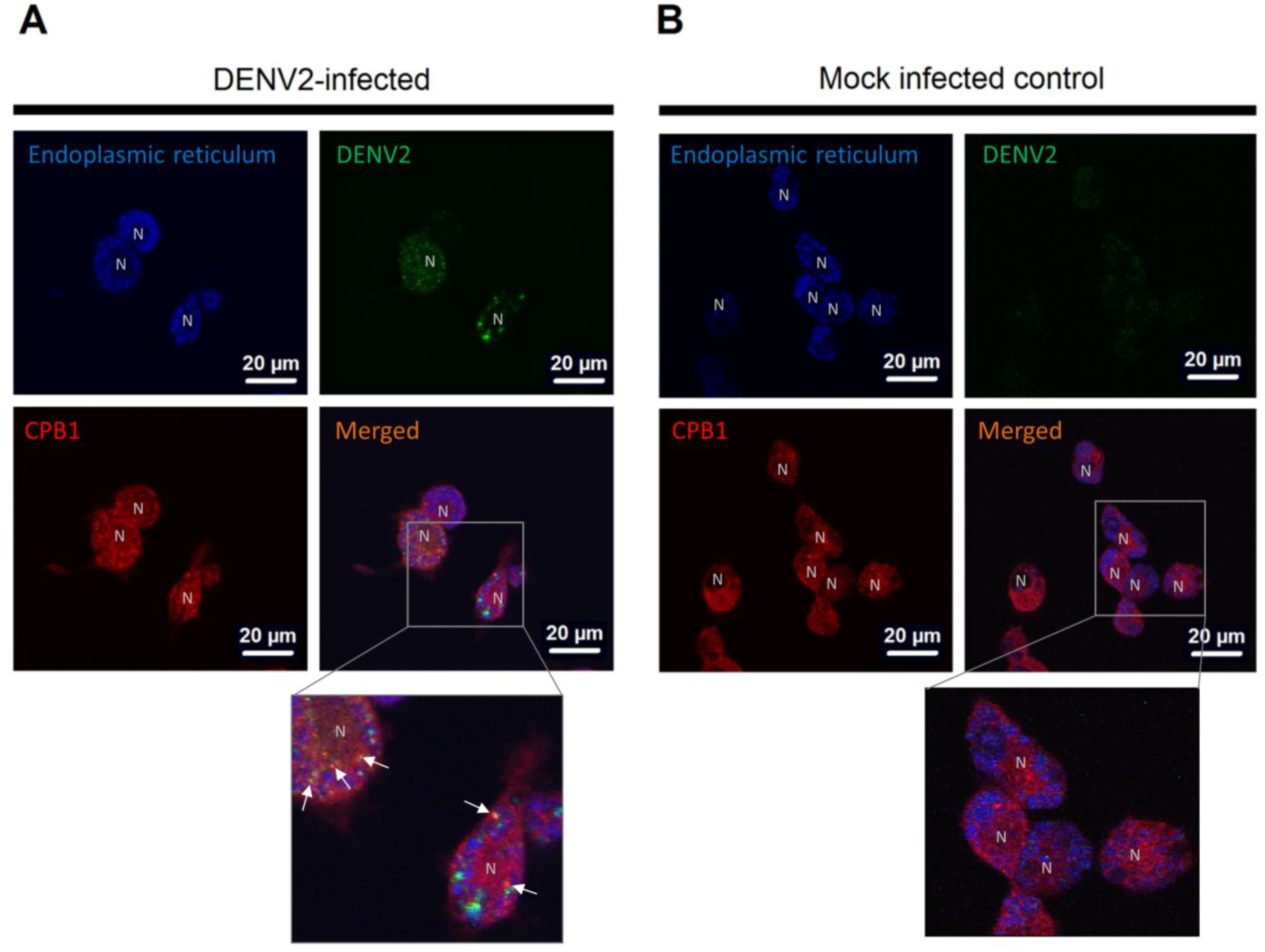
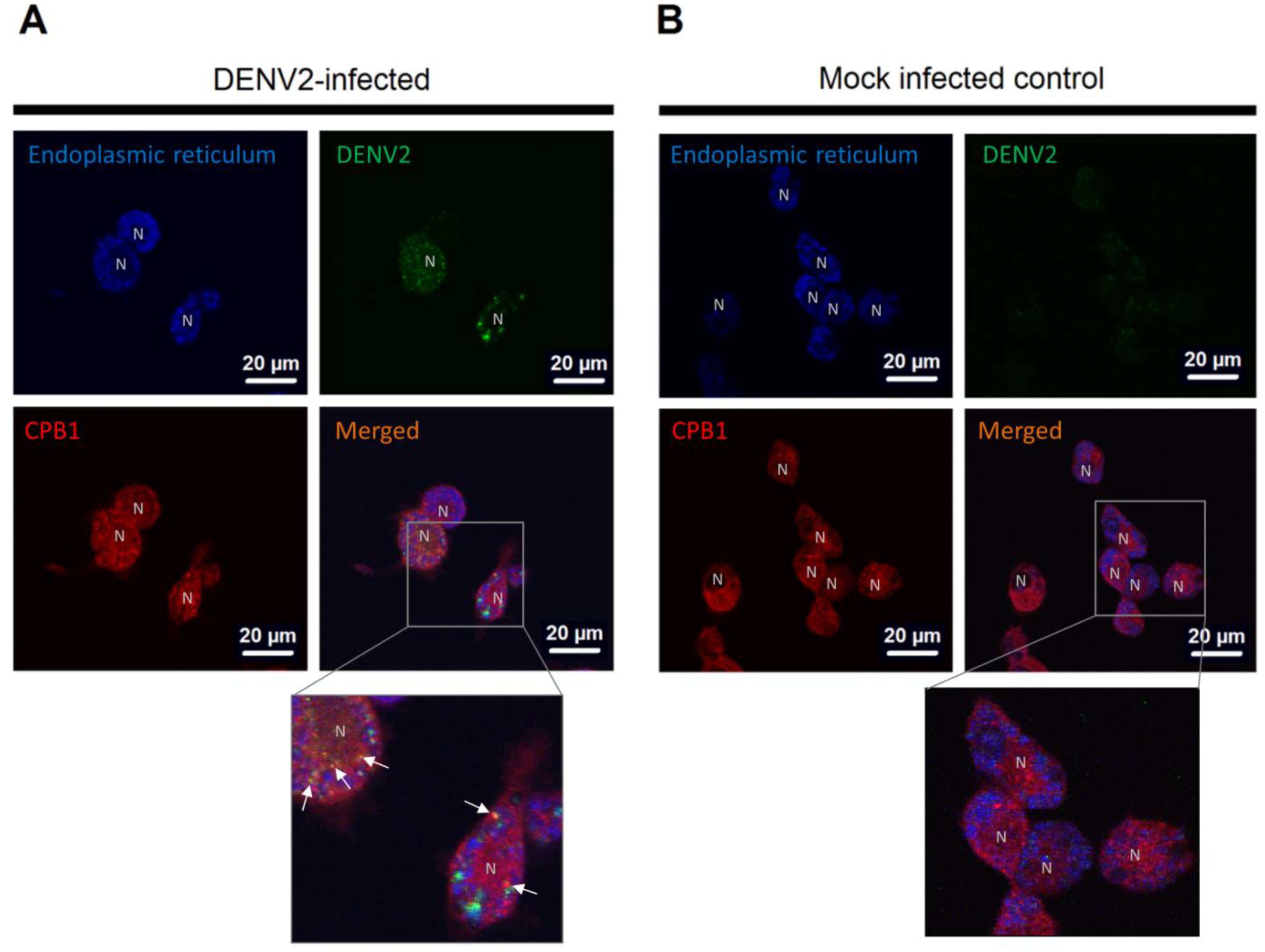
3.3. CPB1 Co-Localizes with Dengue Virus 2 E Protein in the Endoplasmic Reticulum (ER) of Ae. aegypti Primary Midgut Cells
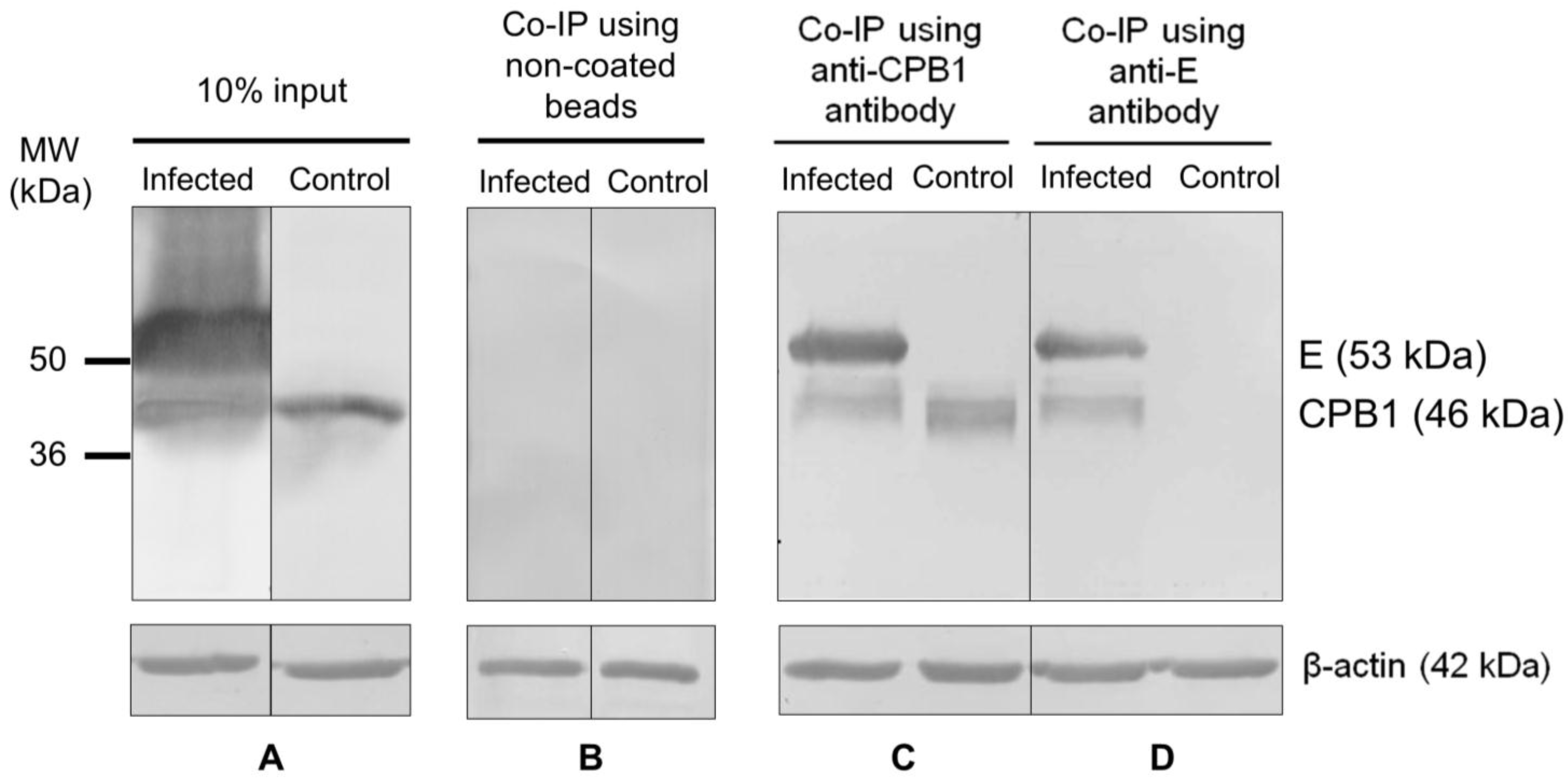
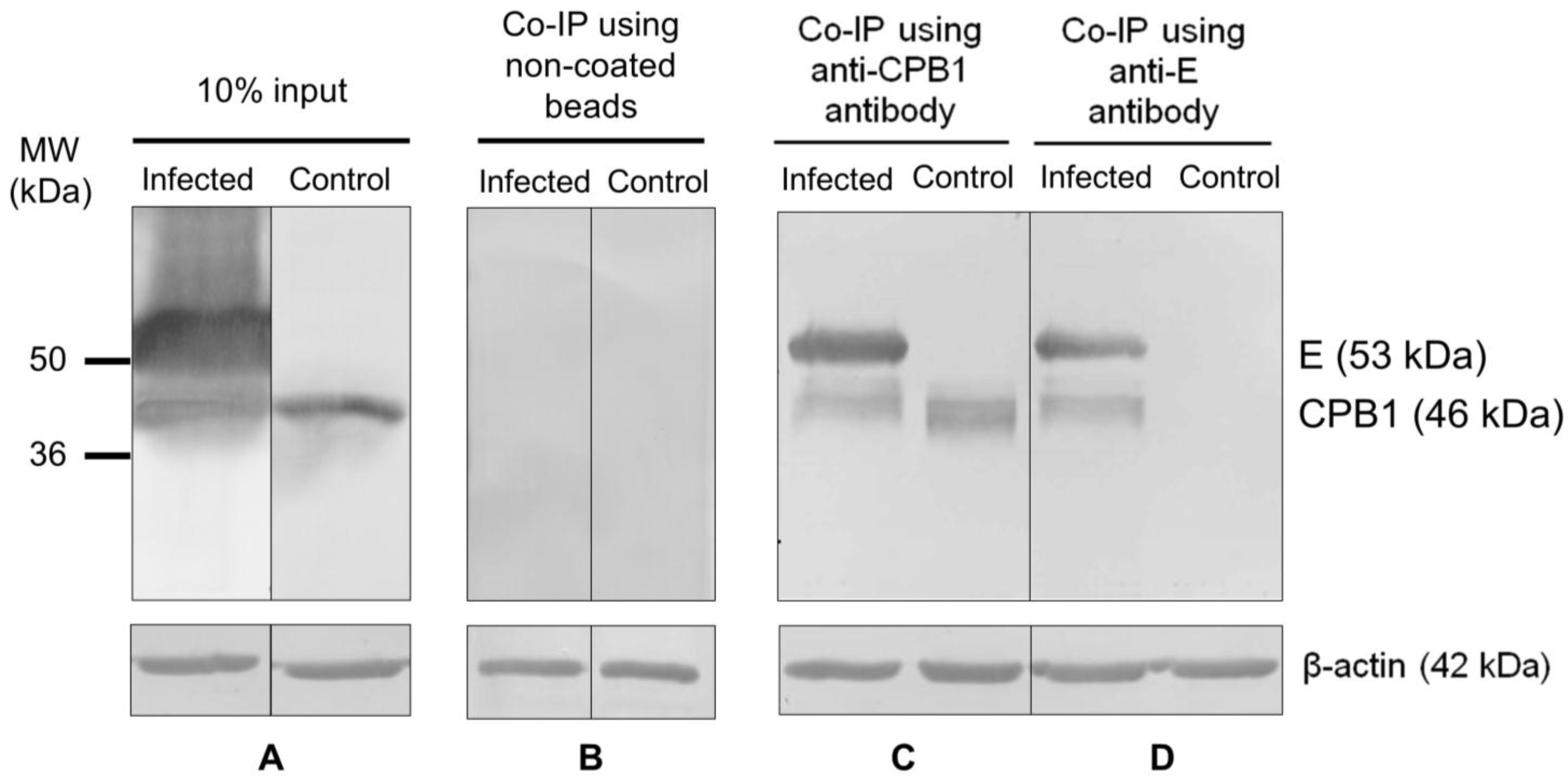
3.4. CPB1-E Protein Complexes Co-Immunoprecipitate from DENV2-Infected Ae. aegypti Primary Midgut Cells
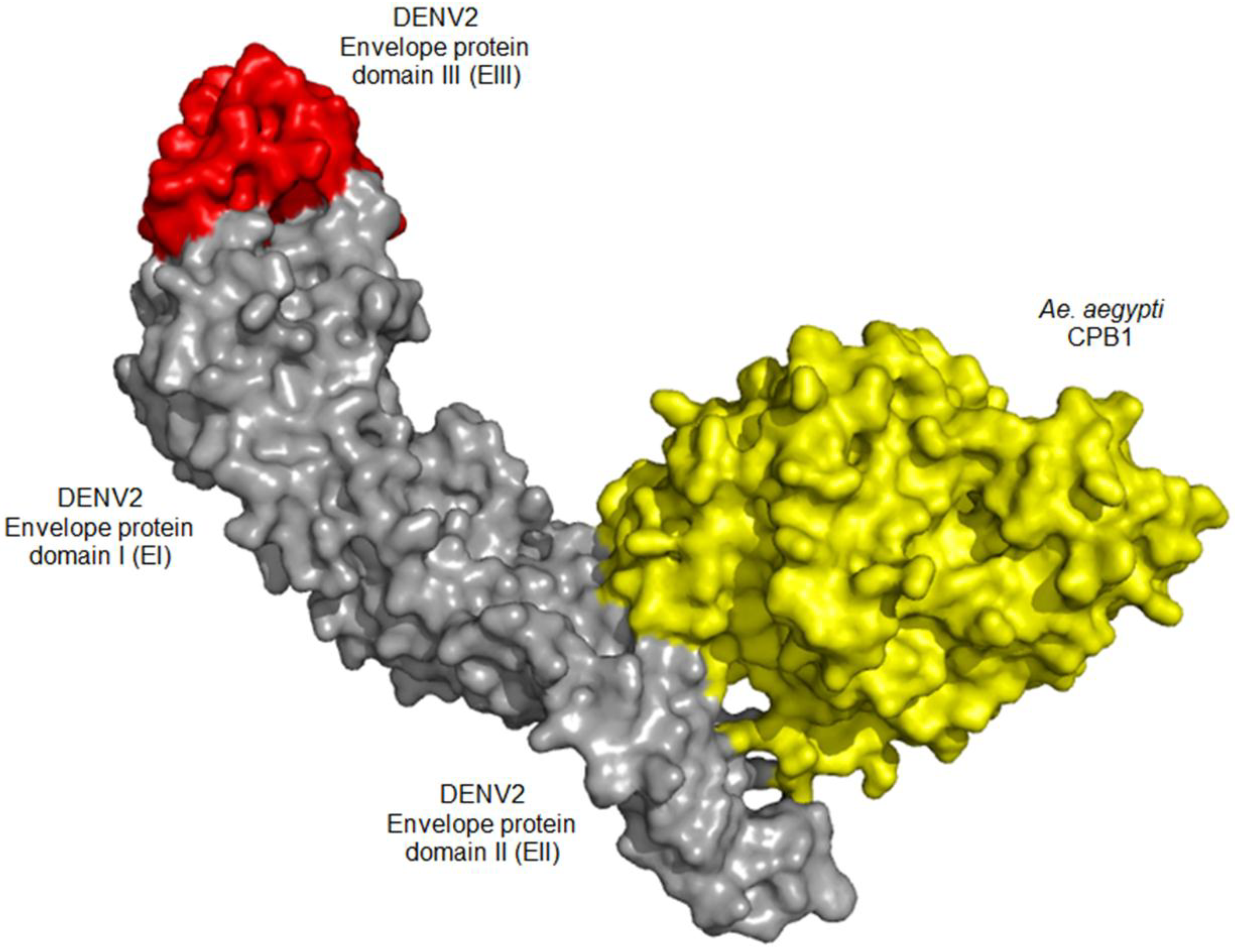
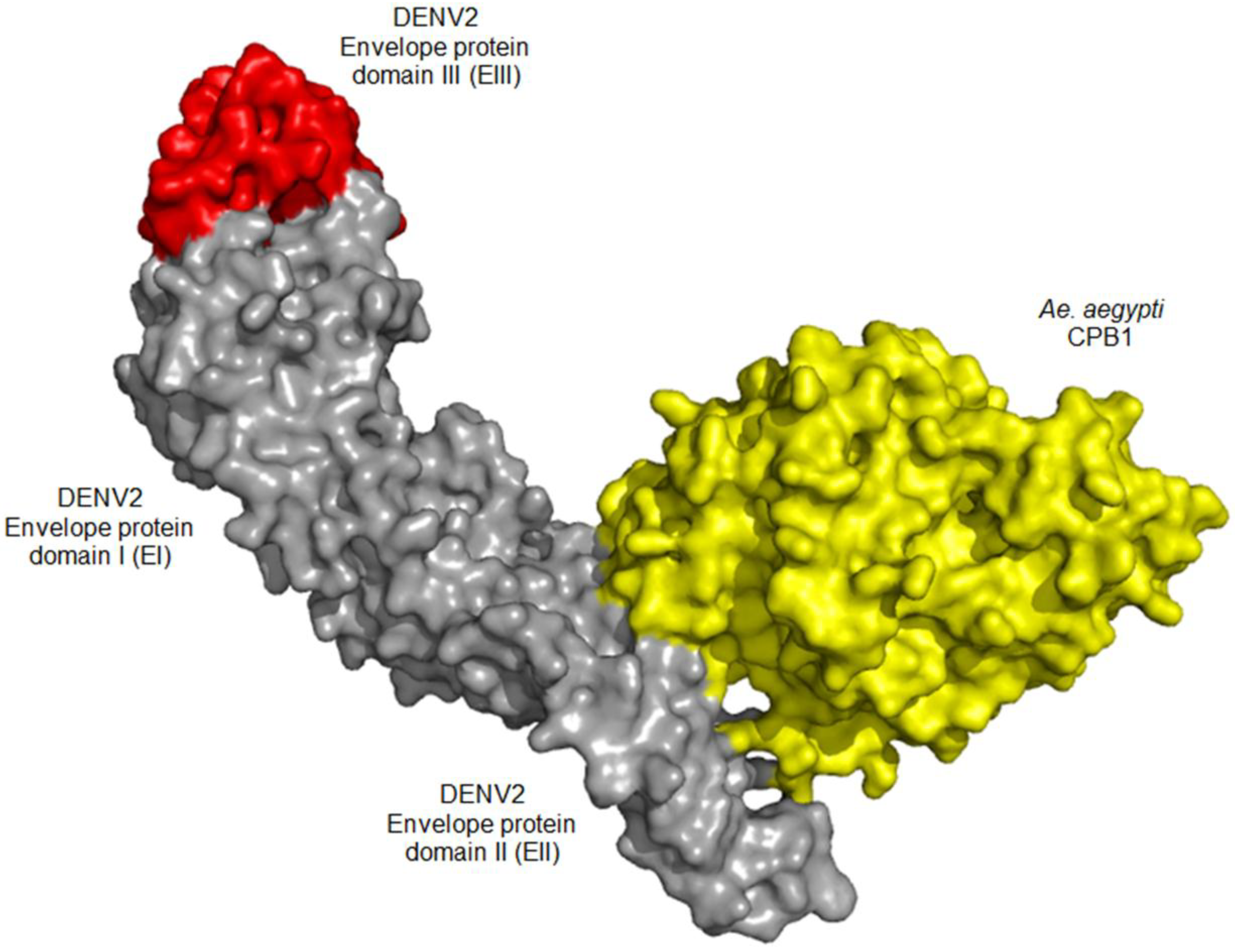
3.5. In Silico Docking Suggests Possible CPB1-E Interaction
3.6. Overexpression of CPB1 in Mosquito C6/36 Cells Results in Intracellular Accumulation of DENV2 Genomic RNA and Viral Components
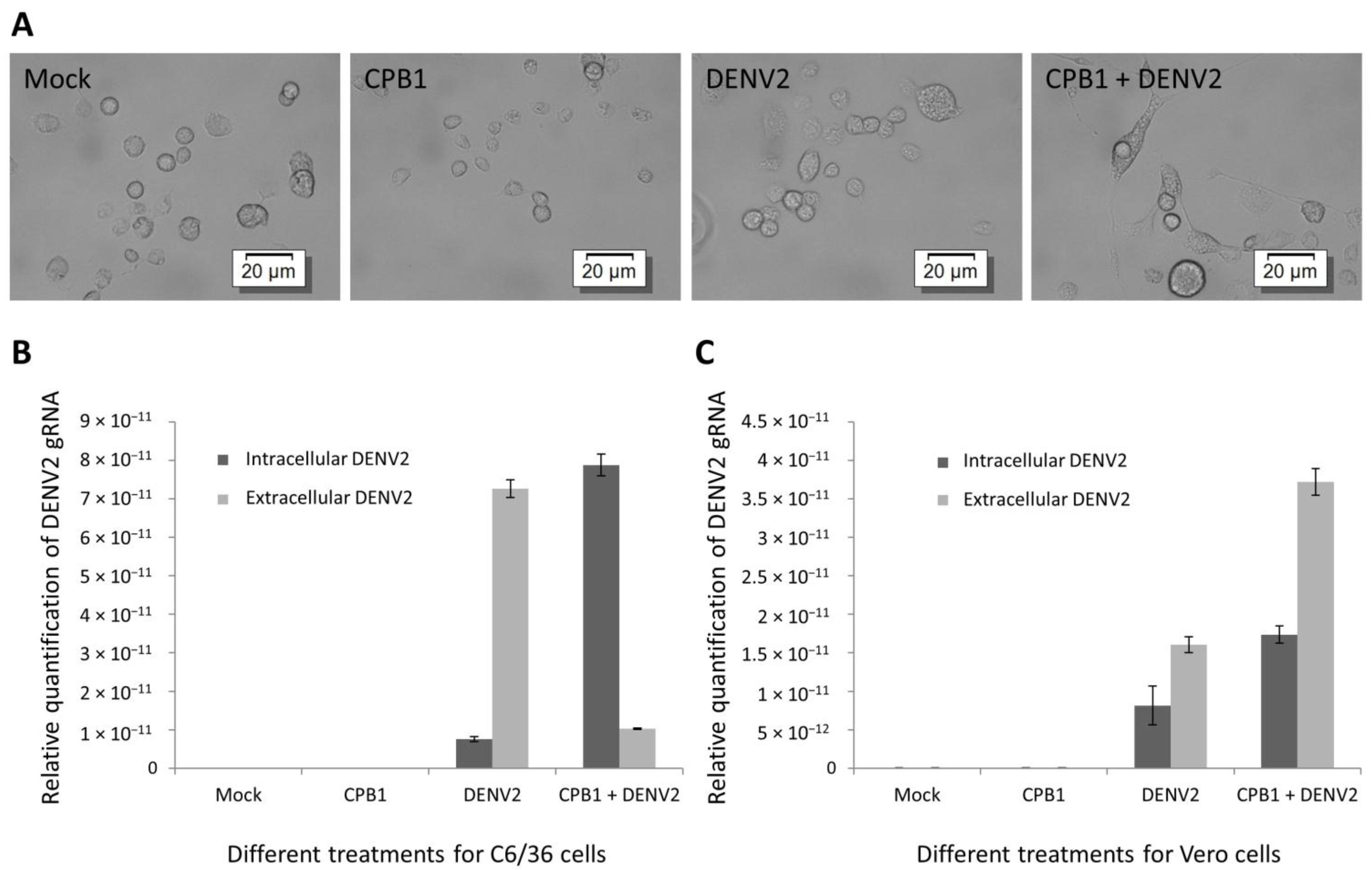
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Samsa, M.M.; Mondotte, J.A.; Iglesias, N.G.; Assuncao-Miranda, I.; Barbosa-Lima, G.; Da Poian, A.T.; Bozza, P.T.; Gamarnik, A.V. Dengue virus capsid protein usurps lipid droplets for viral particle formation. PLoS Pathog. 2009, 5, e1000632. [Google Scholar] [CrossRef] [PubMed]
- Rodenhuis-Zybert, I.A.; Wilschut, J.; Smit, J.M. Dengue virus life cycle: Viral and host factors modulating infectivity. Cell Mol. Life Sci. 2010, 67, 2773–2786. [Google Scholar] [CrossRef] [PubMed]
- Yu, I.M.; Zhang, W.; Holdaway, H.A.; Li, L.; Kostyuchenko, V.A.; Chipman, P.R.; Kuhn, R.J.; Rossmann, M.G.; Chen, J. Structure of the immature dengue virus at low pH primes proteolytic maturation. Science 2008, 319, 1834–1837. [Google Scholar] [CrossRef] [PubMed]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-mediated endocytosis. J. Gen. Virol. 2008, 89, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lormeau, V.M. Dengue viruses binding proteins from Aedes aegypti and Aedes polynesiensis salivary glands. Virol. J. 2009, 6, e35. [Google Scholar] [CrossRef]
- Mairiang, D.; Zhang, H.; Sodja, A.; Murali, T.; Suriyaphol, P.; Malasit, P.; Limjindaporn, T.; Finley, R.L., Jr. Identification of new protein interactions between dengue fever virus and its hosts, human and mosquito. PLoS One 2013, 8, e53535. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Jupatanakul, N.; Ramirez, J.L.; Kang, S.; Romero-Vivas, C.M.; Mohammed, H.; Dimopoulos, G. Transcriptomic profiling of diverse Aedes aegypti strains reveals increased basal-level immune activation in dengue virus-refractory populations and identifies novel virus-vector molecular interactions. PLoS Negl. Trop. Dis. 2013, 7, e2295. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, G.M. Fighting the global pest problem: Preface to the special Toxicon issue on insecticidal toxins and their potential for insect pest control. Toxicon 2007, 49, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Shearman, D.C. The evolution of sex determination systems in dipteran insects other than Drosophila. Genetica 2002, 116, 25–43. [Google Scholar] [CrossRef] [PubMed]
- George, T.; Shaikh, A.I.; Thomas, L.; Kundavaram, A.P. Severe methemoglobinemia due to insecticide poisoning. Indian J. Crit. Care Med. 2014, 18, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Heudorf, U.; Angerer, J.; Drexler, H. Current internal exposure to pesticides in children and adolescents in Germany: Urinary levels of metabolites of pyrethroid and organophosphorus insecticides. Int. Arch. Occup. Environ. Health 2004, 77, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M. Environmental toxins and health—The health impact of pesticides. Aust. Fam. Physician 2007, 36, 1002–1004. [Google Scholar] [PubMed]
- Ramwell, C.T.; Leak, J.; Cooper, S.E.; Taylor, W.A. The potential environmental impact of pesticides removed from sprayers during cleaning. Pest Manag. Sci. 2007, 63, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanam, M.; Forgash, A.J. Environmental impact of mosquito pesticides: Influence of temefos on the brain acetylcholinesterase of killifish. Environ. Physiol. Biochem. 1975, 5, 451–459. [Google Scholar] [PubMed]
- Ciota, A.T.; Kramer, L.D. Vector-virus interactions and transmission dynamics of West Nile virus. Viruses 2013, 5, 3021–3047. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Schiestl, R.H. Quick and easy yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.L.; Phong, T.V.; Rohani, A. Effects of ribavirin and hydroxyurea on oral infection of Aedes aegypti (L.) with dengue virus. Southeast Asian J. Trop. Med. Publ. Health 2012, 43, 1358–1364. [Google Scholar]
- Rohani, A.; Zamree, I.; Joseph, R.T.; Lee, H.L. Persistency of transovarial dengue virus in Aedes aegypti (Linn.). Southeast Asian J. Trop. Med. Publ. Health 2008, 39, 813–816. [Google Scholar]
- Hakim, R.S.; Caccia, S.; Loeb, M.; Smagghe, G. Primary culture of insect midgut cells. Vitro Anim. Cell. Dev. Biol. 2009, 45, 106–110. [Google Scholar] [CrossRef]
- Tchioffo, M.T.; Boissiere, A.; Churcher, T.S.; Abate, L.; Gimonneau, G.; Nsango, S.E.; Awono-Ambene, P.H.; Christen, R.; Berry, A.; Morlais, I. Modulation of malaria infection in Anopheles gambiae mosquitoes exposed to natural midgut bacteria. PLoS One 2013, 8, e81663. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Brayner, F.A.; Alves, L.C.; Dixit, R.; Barillas-Mury, C. Hemocyte differentiation mediates innate immune memory in Anopheles gambiae mosquitoes. Science 2010, 329, 1353–1355. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.; Juhn, J.; James, A.A. Dissection of midgut and salivary glands from Ae. aegypti mosquitoes. J. Vis. Exp. 2007, 5, e228. [Google Scholar]
- Gao, X.; Jia, R.; Wang, M.; Zhu, D.; Chen, S.; Lin, M.; Yin, Z.; Wang, Y.; Chen, X.; Cheng, A. Construction and identification of a cDNA library for use in the yeast two-hybrid system from duck embryonic fibroblast cells post-infected with duck enteritis virus. Mol. Biol. Rep. 2014, 41, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Liu, B.; Zhang, X.Y.; Zhou, Q.M.; Zhang, T.; Li, H.; Yu, Y.F.; Zhang, X.L.; Hao, X.Y.; Wang, M.; Wang, L.; Wei, J.C. Genome characteristics reveal the impact of lichenization on lichen-forming fungus Endocarpon pusillum Hedwig (Verrucariales, Ascomycota). BMC Genomics 2014, 15, e34. [Google Scholar] [CrossRef]
- Zheng, Y.; Tan, X.; Pyczek, J.; Nolte, J.; Pantakani, D.V.; Engel, W. Generation and characterization of yeast two-hybrid cDNA libraries derived from two distinct mouse pluripotent cell types. Mol. Biotechnol. 2013, 54, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Le Breton, M.; Meyniel-Schicklin, L.; Deloire, A.; Coutard, B.; Canard, B.; de Lamballerie, X.; Andre, P.; Rabourdin-Combe, C.; Lotteau, V.; Davoust, N. Flavivirus NS3 and NS5 proteins interaction network: A high-throughput yeast two-hybrid screen. BMC Microbiol. 2011, 11, e234. [Google Scholar] [CrossRef]
- Silva, E.M.; Conde, J.N.; Allonso, D.; Nogueira, M.L.; Mohana-Borges, R. Mapping the Interactions of Dengue Virus NS1 Protein with Human Liver Proteins Using a Yeast Two-Hybrid System: Identification of C1q as an Interacting Partner. PLoS One 2013, 8, e57514. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Batalao, A.; Zhou, H.; Zhu, L. Mammalian two-hybrid system: A complementary approach to the yeast two-hybrid system. Biotechniques 1997, 22, 350–352. [Google Scholar] [PubMed]
- Limjindaporn, T.; Wongwiwat, W.; Noisakran, S.; Srisawat, C.; Netsawang, J.; Puttikhunt, C.; Kasinrerk, W.; Avirutnan, P.; Thiemmeca, S.; Sriburi, R.; Sittisombut, N.; Malasit, P.; Yenchitsomanus, P.T. Interaction of dengue virus envelope protein with endoplasmic reticulum-resident chaperones facilitates dengue virus production. Biochem. Biophys. Res. Commun. 2009, 379, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinformatics 2008, 9, e40. [Google Scholar] [CrossRef]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef] [PubMed]
- Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: Fast interaction refinement in molecular docking. Proteins 2007, 69, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Mashiach, E.; Schneidman-Duhovny, D.; Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: A web server for fast interaction refinement in molecular docking. Nucleic Acids Res. 2008, 36, W229–W232. [Google Scholar] [CrossRef] [PubMed]
- Oezbey, S.; Stengel, C.; Schlotzer-Schrehardt, U.; Ekici, A.; Rautenstrauss, B. Heterologous expression of wildtype and mutant myocilin in High Five insect cells shows comparable effects to cultivated trabecular meshwork cells. Biomol. Eng. 2007, 24, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Wen, N.; Wang, Y.; Wen, L.; Zhao, S.H.; Ai, Z.H.; Wang, Y.; Wu, B.; Lu, H.X.; Yang, H.; Liu, W.C.; Li, Y. Overexpression of FOXM1 predicts poor prognosis and promotes cancer cell proliferation, migration and invasion in epithelial ovarian cancer. J. Trans. Med. 2014, 12, e134. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, V.R.; Hong Wai, T.; Ario Tejo, B.; Omar, A.R.; Syed Hassan, S. Highly pathogenic avian influenza virus nucleoprotein interacts with TREX complex adaptor protein Aly/REF. PLoS One 2013, 8, e72429. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Srivastava, O.P. Deamidation affects structural and functional properties of human alphaA-crystallin and its oligomerization with alphaB-crystallin. J. Biol. Chem. 2004, 279, 44258–44269. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.J.; Lemos, F.J.; Donnelly-Doman, M.; Jacobs-Lorena, M. Rapid induction by a blood meal of a carboxypeptidase gene in the gut of the mosquito Anopheles gambiae. Insect Biochem. Mol. Biol. 1997, 27, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Isoe, J.; Zamora, J.; Miesfeld, R.L. Molecular analysis of the Aedes aegypti carboxypeptidase gene family. Insect Biochem. Mol. Biol. 2009, 39, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Raz, A.; Dinparast Djadid, N.; Zakeri, S. Molecular characterization of the Carboxypeptidase B1 of Anopheles stephensi and its evaluation as a target for transmission-blocking vaccine (TBV). Infect. Immun. 2013, 81, 2206–2216. [Google Scholar] [CrossRef] [PubMed]
- Breiner, K.M.; Urban, S.; Schaller, H. Carboxypeptidase D (gp180), a Golgi-resident protein, functions in the attachment and entry of avian hepatitis B viruses. J. Virol. 1998, 72, 8098–8104. [Google Scholar] [PubMed]
- Spangenberg, H.C.; Lee, H.B.; Li, J.; Tan, F.; Skidgel, R.; Wands, J.R.; Tong, S. A short sequence within domain C of duck carboxypeptidase D is critical for duck hepatitis B virus binding and determines host specificity. J. Virol. 2001, 75, 10630–10642. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Li, J.; Wands, J.R. Carboxypeptidase D is an avian hepatitis B virus receptor. J. Virol. 1999, 73, 8696–8702. [Google Scholar] [PubMed]
- Harris, J.; Schwinn, N.; Mahoney, J.A.; Lin, H.H.; Shaw, M.; Howard, C.J.; da Silva, R.P.; Gordon, S. A vitellogenic-like carboxypeptidase expressed by human macrophages is localized in endoplasmic reticulum and membrane ruffles. Int. J. Exp. Pathol. 2006, 87, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Noriega, F.G.; Edgar, K.A.; Bechet, R.; Wells, M.A. Midgut exopeptidase activities in Aedes aegypti are induced by blood feeding. J. Insect Physiol. 2002, 48, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Kamen, A.A.; Bedard, C.; Tom, R.; Perret, S.; Jardin, B. On-line monitoring of respiration in recombinant-baculovirus infected and uninfected insect cell bioreactor cultures. Biotechnol. Bioeng. 1996, 50, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Hong, H.; Yue, J.; Wu, Y.; Li, X.; Jiang, L.; Li, L.; Li, Q.; Gao, G.; Yang, X. Inhibitory effect of small interfering RNA on dengue virus replication in mosquito cells. Virol. J. 2010, 7, 270. [Google Scholar] [CrossRef] [PubMed]
- Mateo, R.; Nagamine, C.M.; Spagnolo, J.; Mendez, E.; Rahe, M.; Gale, M., Jr.; Yuan, J.; Kirkegaard, K. Inhibition of cellular autophagy deranges dengue virion maturation. J. Virol. 2013, 87, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Heaton, N.S.; Randall, G. Dengue virus and autophagy. Viruses 2011, 3, 1332–1341. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Hosoya, T.; Leong, K.M.; Onogi, H.; Okuno, Y.; Hiramatsu, T.; Koyama, H.; Suzuki, M.; Hagiwara, M.; Garcia-Blanco, M.A. The kinase inhibitor SFV785 dislocates dengue virus envelope protein from the replication complex and blocks virus assembly. PLoS One 2011, 6, e23246. [Google Scholar] [CrossRef] [PubMed]
- Courageot, M.P.; Frenkiel, M.P.; Dos Santos, C.D.; Deubel, V.; Despres, P. Alpha-glucosidase inhibitors reduce dengue virus production by affecting the initial steps of virion morphogenesis in the endoplasmic reticulum. J. Virol. 2000, 74, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Whitby, K.; Pierson, T.C.; Geiss, B.; Lane, K.; Engle, M.; Zhou, Y.; Doms, R.W.; Diamond, M.S. Castanospermine, a potent inhibitor of dengue virus infection in vitro and in vivo. J. Virol. 2005, 79, 8698–8706. [Google Scholar] [CrossRef] [PubMed]
- Christian, E.A.; Kahle, K.M.; Mattia, K.; Puffer, B.A.; Pfaff, J.M.; Miller, A.; Paes, C.; Davidson, E.; Doranz, B.J. Atomic-level functional model of dengue virus Envelope protein infectivity. Proc. Natl. Acad. Sci. USA 2013, 110, 18662–18667. [Google Scholar] [CrossRef] [PubMed]
- Talarico, L.B.; Noseda, M.D.; Ducatti, D.R.; Duarte, M.E.; Damonte, E.B. Differential inhibition of dengue virus infection in mammalian and mosquito cells by iota-carrageenan. J. Gen. Virol. 2011, 92, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Hacker, K.; White, L.; de Silva, A.M. N-linked glycans on dengue viruses grown in mammalian and insect cells. J. Gen. Virol. 2009, 90, 2097–2106. [Google Scholar] [CrossRef] [PubMed]
- Farias, K.J.; Machado, P.R.; da Fonseca, B.A. Chloroquine inhibits dengue virus type 2 replication in Vero cells but not in C6/36 cells. Sci. World J. 2013, 2013, e282734. [Google Scholar] [CrossRef]
- Acosta, E.G.; Piccini, L.E.; Talarico, L.B.; Castilla, V.; Damonte, E.B. Changes in antiviral susceptibility to entry inhibitors and endocytic uptake of dengue-2 virus serially passaged in Vero or C6/36 cells. Virus Res. 2014, 184C, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Cirimotich, C.M.; Scott, J.C.; Phillips, A.T.; Geiss, B.J.; Olson, K.E. Suppression of RNA interference increases alphavirus replication and virus-associated mortality in Aedes aegypti mosquitoes. BMC Microbiol. 2009, 9, e49. [Google Scholar] [CrossRef]
- Ding, S.W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef] [PubMed]
- Sabin, L.R.; Zhou, R.; Gruber, J.J.; Lukinova, N.; Bambina, S.; Berman, A.; Lau, C.K.; Thompson, C.B.; Cherry, S. Ars2 regulates both miRNA- and siRNA- dependent silencing and suppresses RNA virus infection in Drosophila. Cell 2009, 138, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.; Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito's RNA interference pathway. PLoS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [PubMed]
- Siu, R.W.; Fragkoudis, R.; Simmonds, P.; Donald, C.L.; Chase-Topping, M.E.; Barry, G.; Attarzadeh-Yazdi, G.; Rodriguez-Andres, J.; Nash, A.A.; Merits, A.; Fazakerley, J.K.; Kohl, A. Antiviral RNA interference responses induced by Semliki Forest virus infection of mosquito cells: Characterization, origin, and frequency-dependent functions of virus-derived small interfering RNAs. J. Virol. 2011, 85, 2907–2917. [Google Scholar] [CrossRef] [PubMed]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl. Acad. Sci. USA 2009, 106, 17841–17846. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.S.; Rico-Hesse, R. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. J. Virol. 2012, 86, 7637–7649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, X.; Wu, Y.; Gan, M.; He, A.; Li, Z.; Liu, J.; Zhan, X. Quantitative analysis of replication and tropisms of Dengue virus type 2 in Aedes albopictus. Am. J. Trop. Med. Hyg. 2010, 83, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Dubrulle, M.; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.B. Chikungunya virus and Aedes mosquitoes: Saliva is infectious as soon as two days after oral infection. PLoS One 2009, 4, e5895. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tham, H.-W.; Balasubramaniam, V.R.M.T.; Tejo, B.A.; Ahmad, H.; Hassan, S.S. CPB1 of Aedes aegypti Interacts with DENV2 E Protein and Regulates Intracellular Viral Accumulation and Release from Midgut Cells. Viruses 2014, 6, 5028-5046. https://doi.org/10.3390/v6125028
Tham H-W, Balasubramaniam VRMT, Tejo BA, Ahmad H, Hassan SS. CPB1 of Aedes aegypti Interacts with DENV2 E Protein and Regulates Intracellular Viral Accumulation and Release from Midgut Cells. Viruses. 2014; 6(12):5028-5046. https://doi.org/10.3390/v6125028
Chicago/Turabian StyleTham, Hong-Wai, Vinod R. M. T. Balasubramaniam, Bimo Ario Tejo, Hamdan Ahmad, and Sharifah Syed Hassan. 2014. "CPB1 of Aedes aegypti Interacts with DENV2 E Protein and Regulates Intracellular Viral Accumulation and Release from Midgut Cells" Viruses 6, no. 12: 5028-5046. https://doi.org/10.3390/v6125028
APA StyleTham, H.-W., Balasubramaniam, V. R. M. T., Tejo, B. A., Ahmad, H., & Hassan, S. S. (2014). CPB1 of Aedes aegypti Interacts with DENV2 E Protein and Regulates Intracellular Viral Accumulation and Release from Midgut Cells. Viruses, 6(12), 5028-5046. https://doi.org/10.3390/v6125028