2.1. Poxviridae
Poxviruses are the largest and most complex of the viruses causing human disease. The most infamous would be variola virus, the causative agent of smallpox, which was eradicated by a concerted global effort overseen by the World Health Organization; the last naturally occurring case was in 1977. Infections with a poxvirus will produce pock(s), or pustule(s), on the skin and also internally on visceral organs with some species. The genera of poxviruses that can cause human disease include Orthopoxvirus, Parapoxvirus, Molluscipoxvirus, and Yatapoxvirus.
Although poxviruses contain DNA, the DNA replication and virus assembly do not take place in the nucleus but rather in the cytoplasm. Virus factories, or virosomes, are created and nascent crescents are formed and engulf the unit genome. Immature particles are spherical, but condense down to a dumbbell-shaped intracellular mature virus (IMV). The IMV is engulfed by Golgi vesicles, migrates to the cell surface or into microvilli and fuses with the cell membrane, releasing a particle wrapped in a single membrane known as an enveloped extracellular virus (EEV).
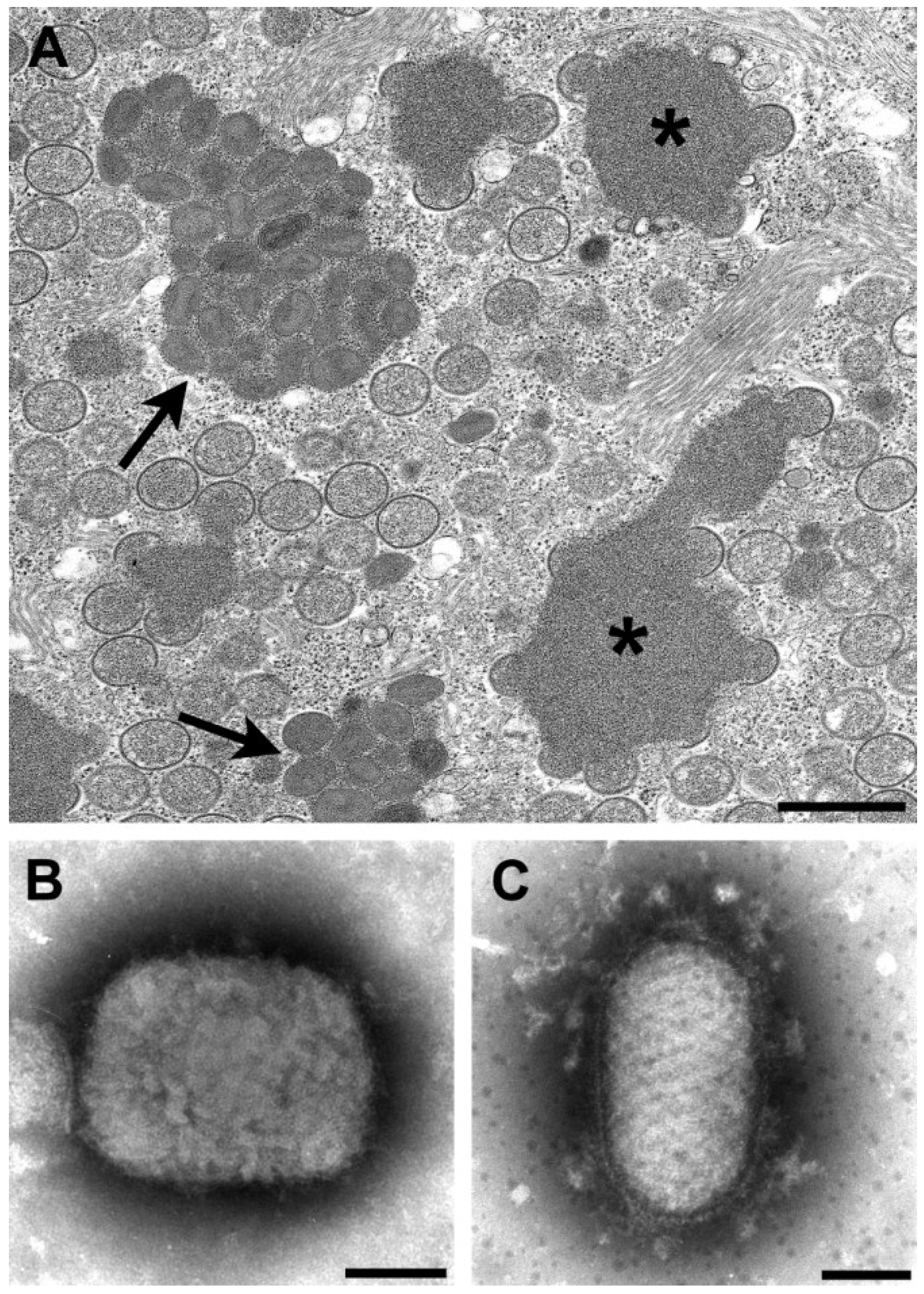
By thin section EM, in addition to the viral factories, some poxviruses have other cytoplasmic structures know as acidophilic-type inclusions (A-type inclusions) consisting of a matrix containing the A-type inclusion protein and other proteins, with occluded intracellular mature particles [
10,
11] (
Figure 1A). Virus species with these inclusions include cowpox, ectromelia, raccoonpox, skunkpox, volepox, and fowlpox viruses.
The viruses in the genera
Orthopoxvirus and
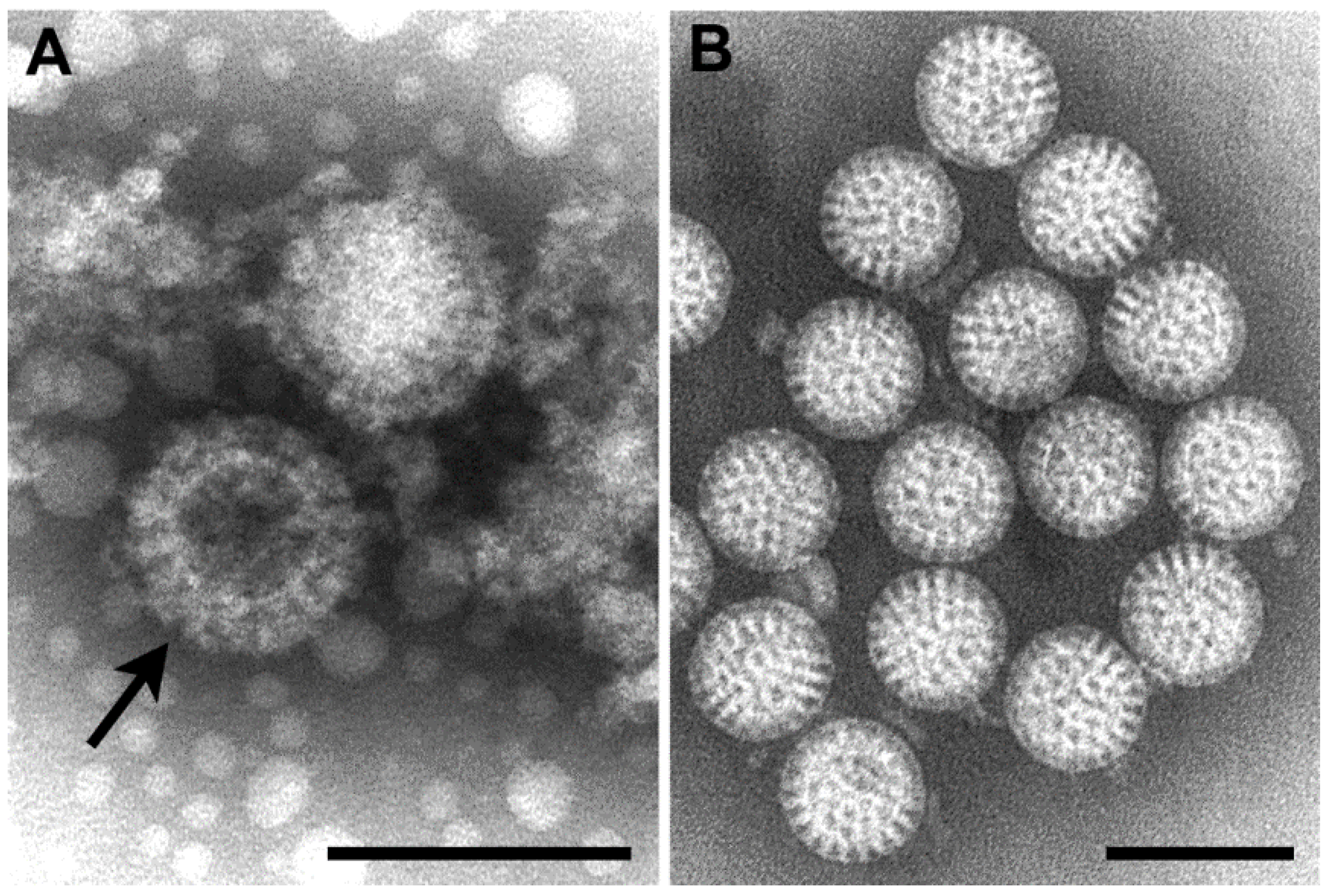
Parapoxvirus can be distinguished by negative stain EM (
Figure 1B,C). Orthopoxviruses are rectangular, approximately 225 × 300 nm in size and have a surface pattern of short, whorled filaments. On the other hand, parapoxviruses are oval, average only about 150 × 200 nm in size, and have a crisscross filamentous surface pattern. Unfortunately, these two genera cannot be definitively differentiated by thin section EM.
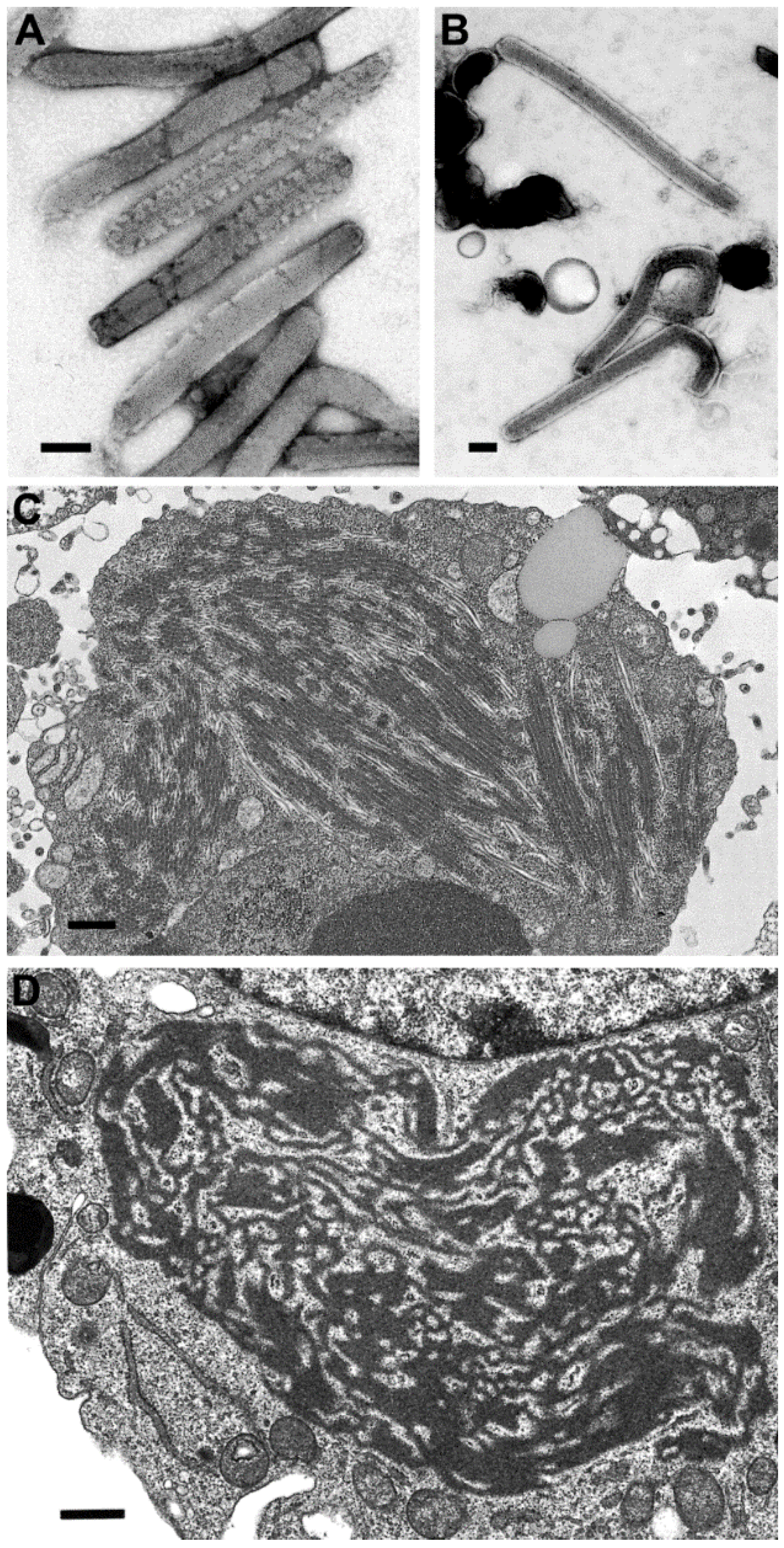
Figure 1.
(A) Thin section image of raccoonpox, showing viral factories (*) and A-type inclusions (arrows). Bar, 500 nm; (B) Negative stain image of a clinical sample of monkeypox virus (genus Orthopoxvirus). Bar, 100 nm; (C) Negative stain image of a clinical sample of orf virus (genus Parapoxvirus). Bar, 100 nm.
Figure 1.
(A) Thin section image of raccoonpox, showing viral factories (*) and A-type inclusions (arrows). Bar, 500 nm; (B) Negative stain image of a clinical sample of monkeypox virus (genus Orthopoxvirus). Bar, 100 nm; (C) Negative stain image of a clinical sample of orf virus (genus Parapoxvirus). Bar, 100 nm.
2.2. Reoviridae
Reoviruses derive their name from
Respiratory
Enteric
Orphan viruses. This paper will describe viruses that are members of the genera
Orthoreovirus and
Rotavirus. Orthoreoviruses are in the subfamily
Spinoreovirinae and contain large spikes or turrets at the 12 icosahedral vertices of the core particle, while rotaviruses are in the subfamily
Sedoreovirinae and do not have large surface projections on the core particles [
12].
Orthoreoviruses are spread by the respiratory or fecal-oral routes. The genome consists of 10 segments of linear double-stranded RNA. By negative stain EM, virions are approximately 85 nm in diameter, are roughly spherical, and possess a double-layered protein capsid (
Figure 2A).
Rotaviruses are the cause of severe diarrheal disease in infants and young children, and were first recognized by EM in 1973 [
13]. The genome is composed of 11 segments of linear double-stranded RNA. By negative stain EM, virus particles are 70 nm in diameter and are constructed of three concentric protein layers. Virions have a wheel-like appearance (
rota is Latin for “wheel”) with a sharp definition of the outer margin (
Figure 2B).
Figure 2.
(A) Negative stain EM image of orthoreovirus particles, with a stain-penetrated particle (arrow) showing the double capsid layers; (B) Negative stain EM image of rotavirus particles. Bars, 100 nm. (Figure B, courtesy of Charles D. Humphrey, Centers for Disease Control and Prevention, Atlanta, GA, USA.)
Figure 2.
(A) Negative stain EM image of orthoreovirus particles, with a stain-penetrated particle (arrow) showing the double capsid layers; (B) Negative stain EM image of rotavirus particles. Bars, 100 nm. (Figure B, courtesy of Charles D. Humphrey, Centers for Disease Control and Prevention, Atlanta, GA, USA.)
2.3. Retroviridae
The retroviruses are divided into two subfamilies. The genera in the subfamily Orthoretrovirinae that can infect humans are Alpharetrovirus, Betaretrovirus, Deltaretrovirus, Gammaretrovirus, and Lentivirus. Spumavirus is the only genus in the subfamily Spumaretrovirinae. The viral genome for all but the spumaviruses consists of a dimer of positive-sense, single-stranded RNA held together by hydrogen bonds. Spumaviruses contain double-stranded DNA. All retroviruses use the enzyme reverse transcriptase to transcribe an RNA template into complementary DNA.
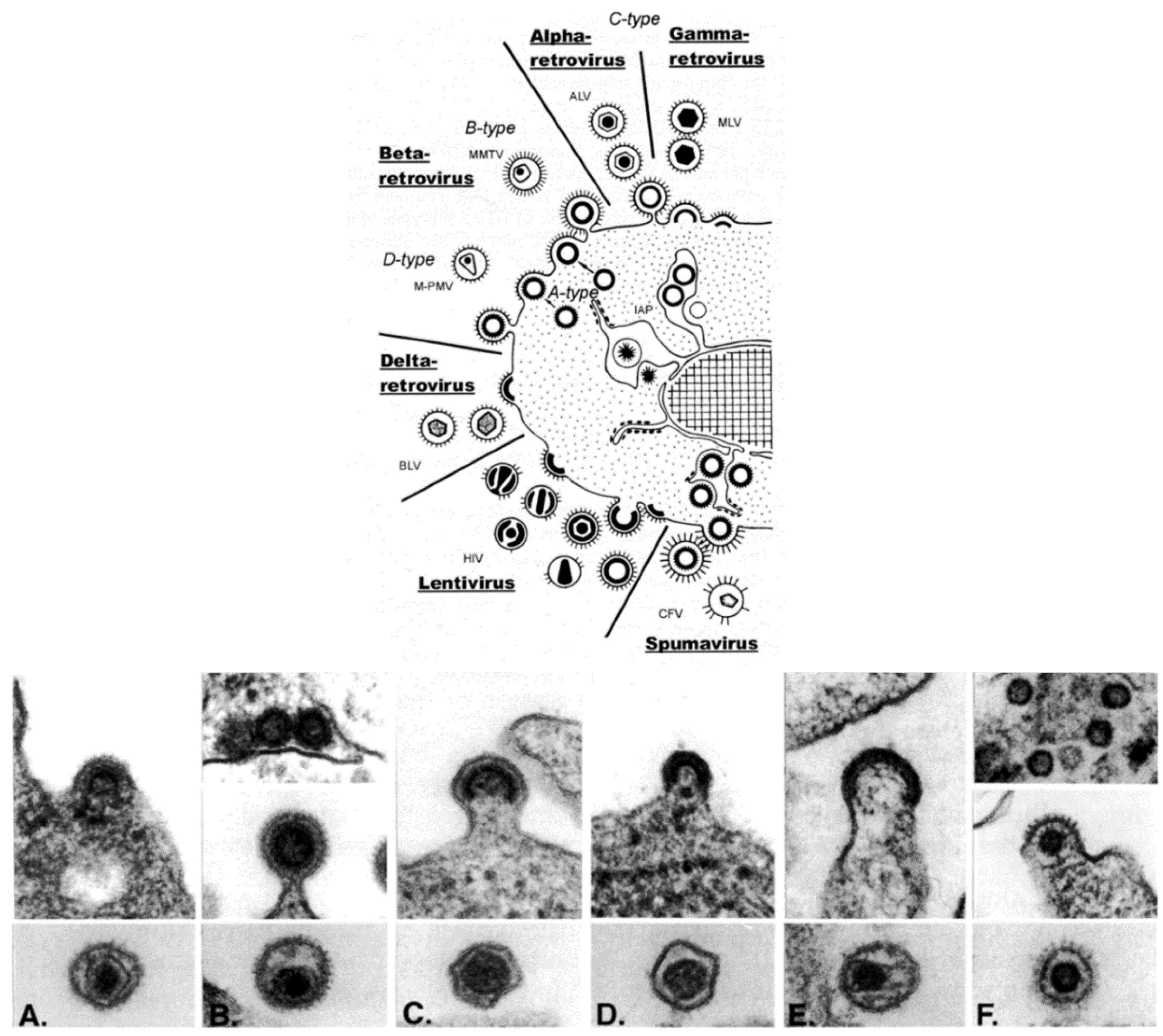
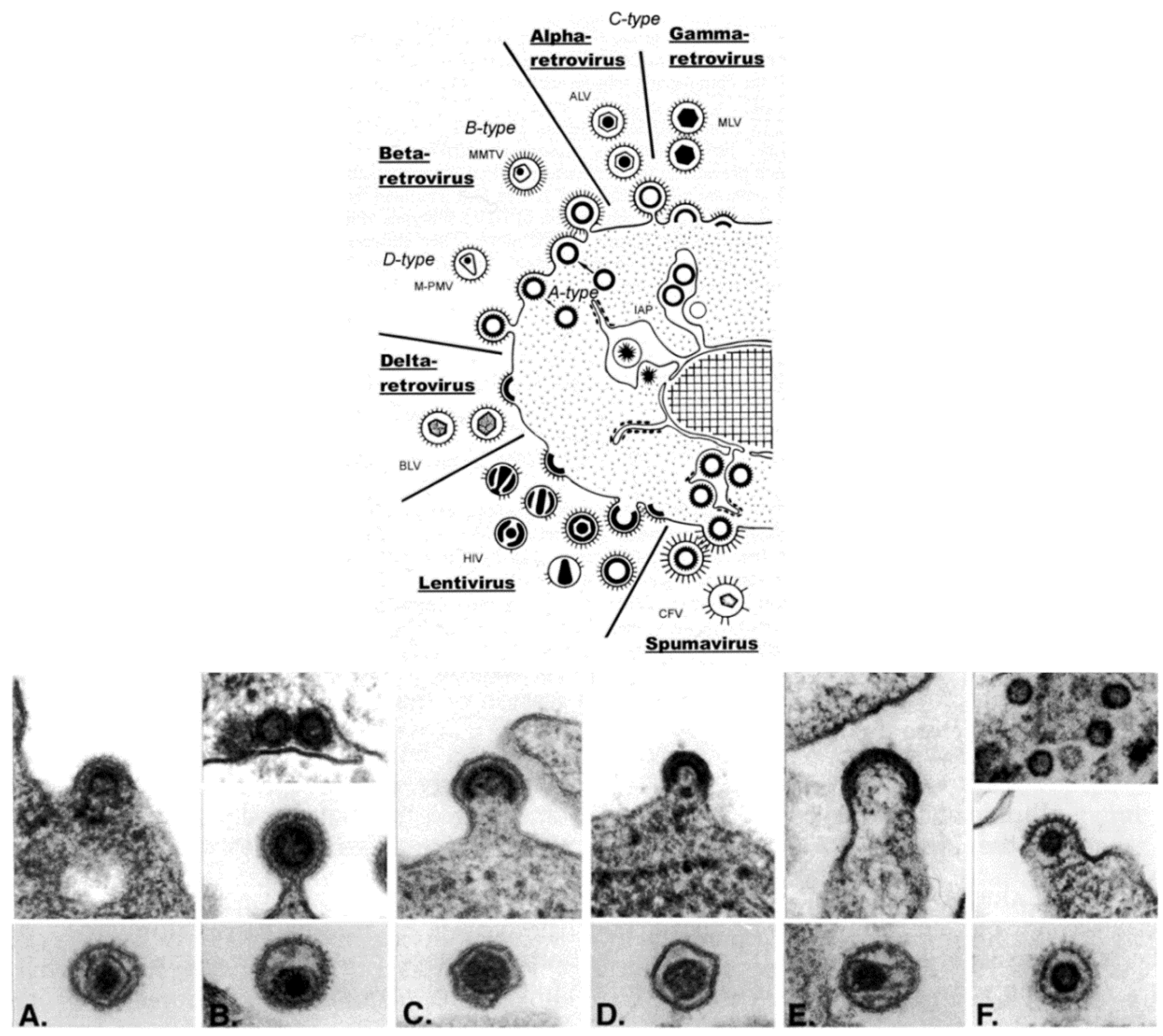
Retroviridae is, morphologically, a multi-faceted family of viruses. As illustrated in a drawing that appeared in a publication by Gelderblom and Boller [
14] (
Figure 3, top), in thin section EM, members of the family can be assigned to a particular genus based on the morphogenesis of the virus and on the appearance of mature virus particles [
15,
16] (
Figure 3, bottom).
The genera Alpharetrovirus and Gammaretrovirus, such as avian leukosis virus (ALV) and murine leukemia virus (MLV), respectively, were previously known as C-type particles; the cores form concomitantly with budding, and are centered in the middle of mature particles. Viruses previously known as B-type (such as mouse mammary tumor virus (MMTV)) and D-type (such as Mason-Pfizer monkey virus (M-PMV)) are now part of the genus Betaretrovirus. Both types are formed by envelopment of pre-formed cores, which are known as A-type particles, and mature into eccentrically located cores surrounded by the viral envelope. The viruses in the genus Deltaretrovirus, such as bovine leukemia virus (BLV), have a crescent-shaped budding profile which is composed of an electron-dense nucleoid and the nascent viral envelope. The cores of the mature virions are somewhat pleomorphic and fairly homogeneous, and there is often an electron-lucent space between the core and the envelope. Human immunodeficiency virus (HIV) is an example of the genus Lentivirus, and these viruses also have a crescent-shaped budding profile, which can be released from the cell to form a doughnut-shaped particle. The nucleoid then condenses into an electron-dense core that is cone-shaped, but can appear as a rod. The viruses in the genus Spumavirus, such as chimpanzee foamy virus (CFV), are seen as contaminants in some cell cultures derived from animal organs. Pre-formed cores are enveloped at cellular membranes or the plasma membrane, and the cores do not condense.
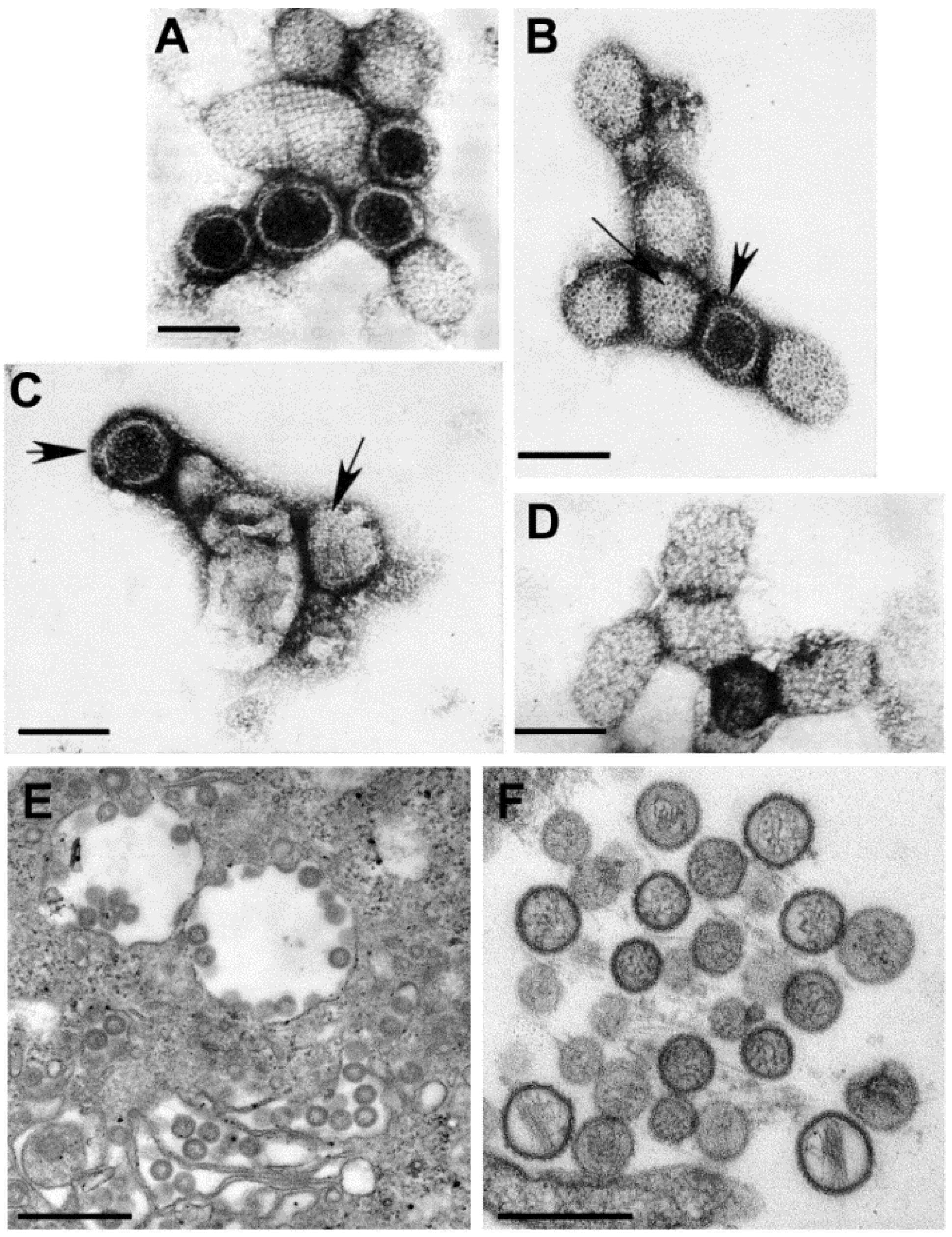
Figure 3.
(
Top) schematic diagram of the morphogenesis of the members of the family
Retroviridae; (
Bottom) budding profiles and mature virions. (
A) Avian leukosis virus (genus
Alpharetrovirus); (
B) Mouse mammary tumor virus (genus
Betaretrovirus); (
C) Murine leukemia virus (genus
Gammaretrovirus); (
D) Bovine leukemia virus (
Deltaretrovirus); (
E) Human immunodeficiency virus 1 (genus
Lentivirus); (
F) Simian foamy virus (genus
Spumavirus). (Top, reproduced with permission from Reference [
14]. Copyright 2002 Kluwer Academic/Plenum Publishers. Bottom, reproduced with permission from Reference [
17]. Copyright 1997 Cold Spring Harbor Laboratory.
Figure 3.
(
Top) schematic diagram of the morphogenesis of the members of the family
Retroviridae; (
Bottom) budding profiles and mature virions. (
A) Avian leukosis virus (genus
Alpharetrovirus); (
B) Mouse mammary tumor virus (genus
Betaretrovirus); (
C) Murine leukemia virus (genus
Gammaretrovirus); (
D) Bovine leukemia virus (
Deltaretrovirus); (
E) Human immunodeficiency virus 1 (genus
Lentivirus); (
F) Simian foamy virus (genus
Spumavirus). (Top, reproduced with permission from Reference [
14]. Copyright 2002 Kluwer Academic/Plenum Publishers. Bottom, reproduced with permission from Reference [
17]. Copyright 1997 Cold Spring Harbor Laboratory.
2.4. Herpesviridae
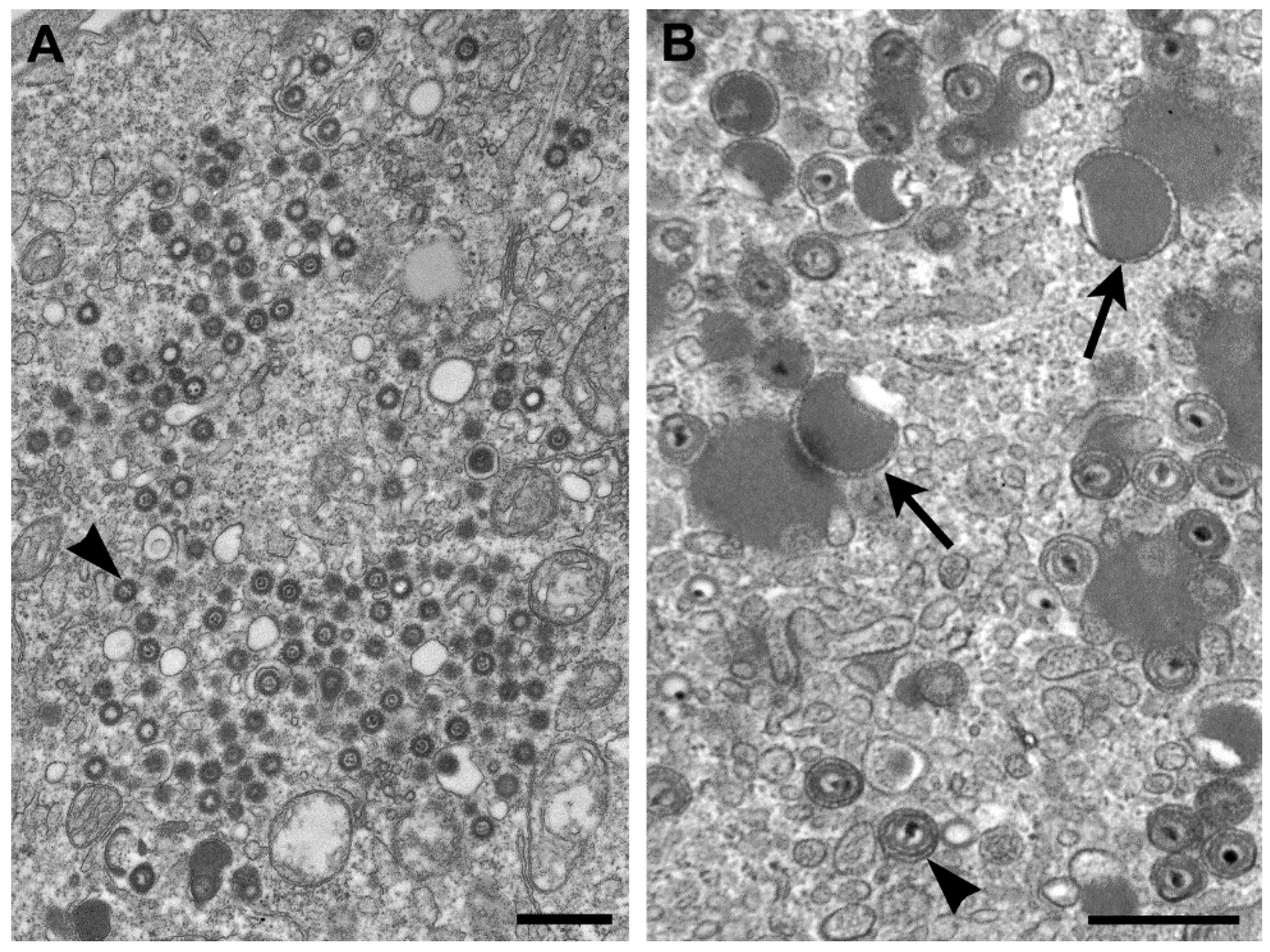
The genus
Cytomegalovirus (CMV) is within the subfamily
Betaherpesvirinae, and infection with these viruses typically results in an increase in cell volume (cytomeglia). CMV-infected cells have nuclear inclusions, characteristic of herpesviruses, but also have cytoplasmic inclusions. The genomes of herpesviruses in general are composed of linear, double-stranded DNA. Replication takes place in the nucleus where nucleocapsids are formed, are surrounded by an “inner” tegument, and travel to the cytoplasm by budding upon the inner nuclear membrane, passing through the perinuclear space, and fusing with the outer nuclear membrane to egress to the cytoplasm. Additional tegument proteins attach to the capsid, either within the cytosol and/or at the future envelopment site on the membranes of the Golgi complex [
18] (
Figure 4A).
Unlike the other members of the family
Herpesviridae, the cytoplasm in cells infected by CMV contains numerous aggregations of enveloped tegument proteins that lack capsids (
Figure 4B). These are known as dense bodies [
19], which are highly immunogenic and have been proposed as CMV vaccine candidates since they induce both humoral and cellular immune responses [
20].
Figure 4.
(A) Thin section EM image of the cytoplasm of a cell infected with human herpesvirus 7. Note that although there are nucleocapsids surrounded by tegument (arrowhead), there are no dense bodies; (B) Cell infected with simian CMV, with dense bodies (arrows) and virus particles (arrowhead) in the cytoplasm. Bars, 500 nm. (Figure B, courtesy of Sara E. Miller, Duke University Medical Center, Durham, NC, USA.)
Figure 4.
(A) Thin section EM image of the cytoplasm of a cell infected with human herpesvirus 7. Note that although there are nucleocapsids surrounded by tegument (arrowhead), there are no dense bodies; (B) Cell infected with simian CMV, with dense bodies (arrows) and virus particles (arrowhead) in the cytoplasm. Bars, 500 nm. (Figure B, courtesy of Sara E. Miller, Duke University Medical Center, Durham, NC, USA.)
2.5. Filoviridae
This family includes Ebolavirus and Marburgvirus genera and the recently recognized Cuevavirus genus, which is not known to cause human disease. Filoviruses are nonsegmented, negative-sense, single-stranded RNA viruses. Virions are pleomorphic, appearing as long filamentous particles, but also as branched, 6-shaped, U-shaped, or circular particles.
The viruses in the genera Ebolavirus and Marburgvirus are well-known as being some of the deadliest known viruses, with case fatality rates for Ebola virus reported at 50% to 90%, and rates for Marburg virus at 24% to 88%. Early symptoms for both diseases include sudden onset of fever, headaches, weakness, muscle pains, and a sore throat. As the diseases progress, additional symptoms such as vomiting, diarrhea, impaired kidney and liver function, and sometimes a rash and internal and external bleeding may develop.
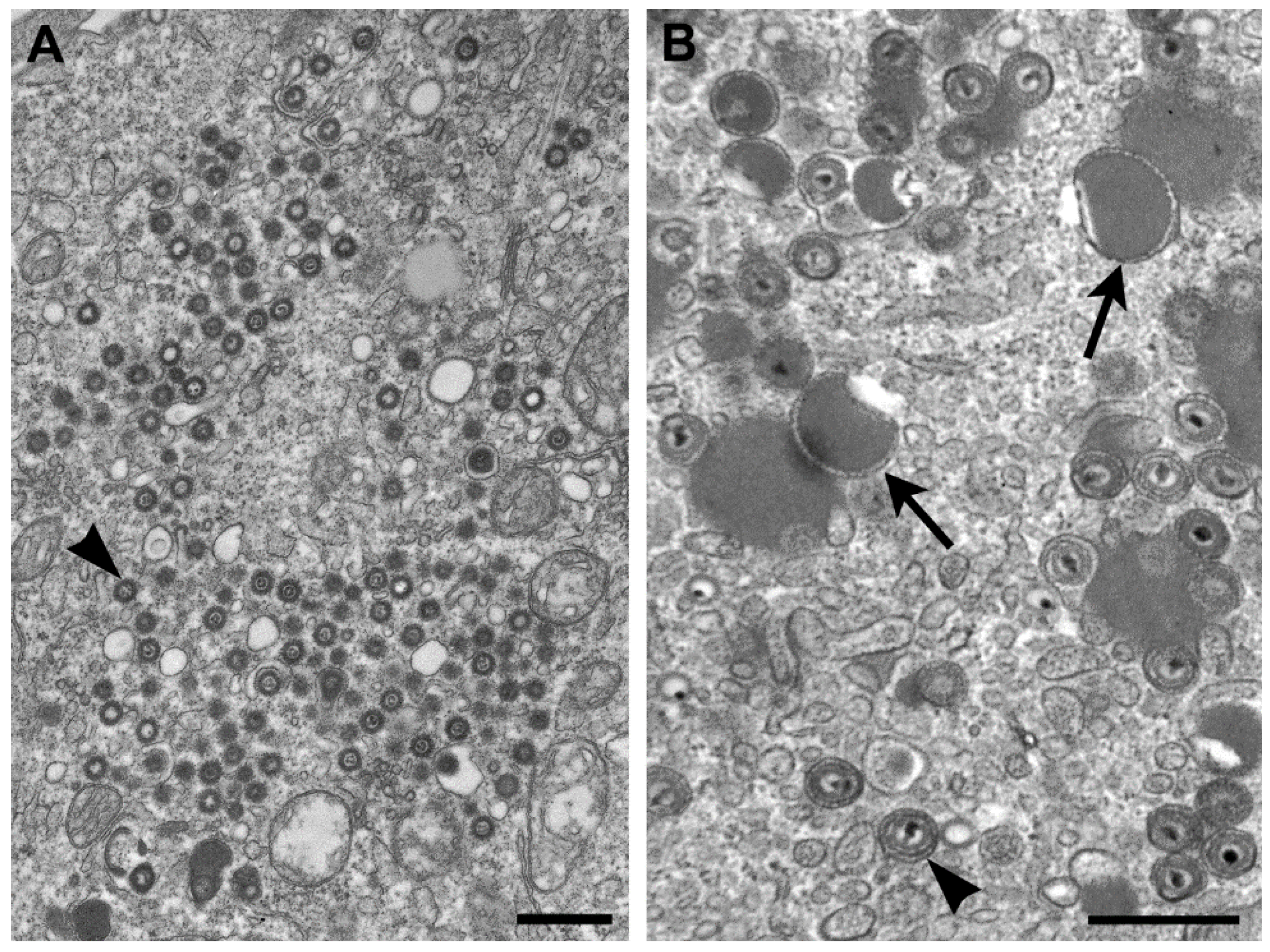
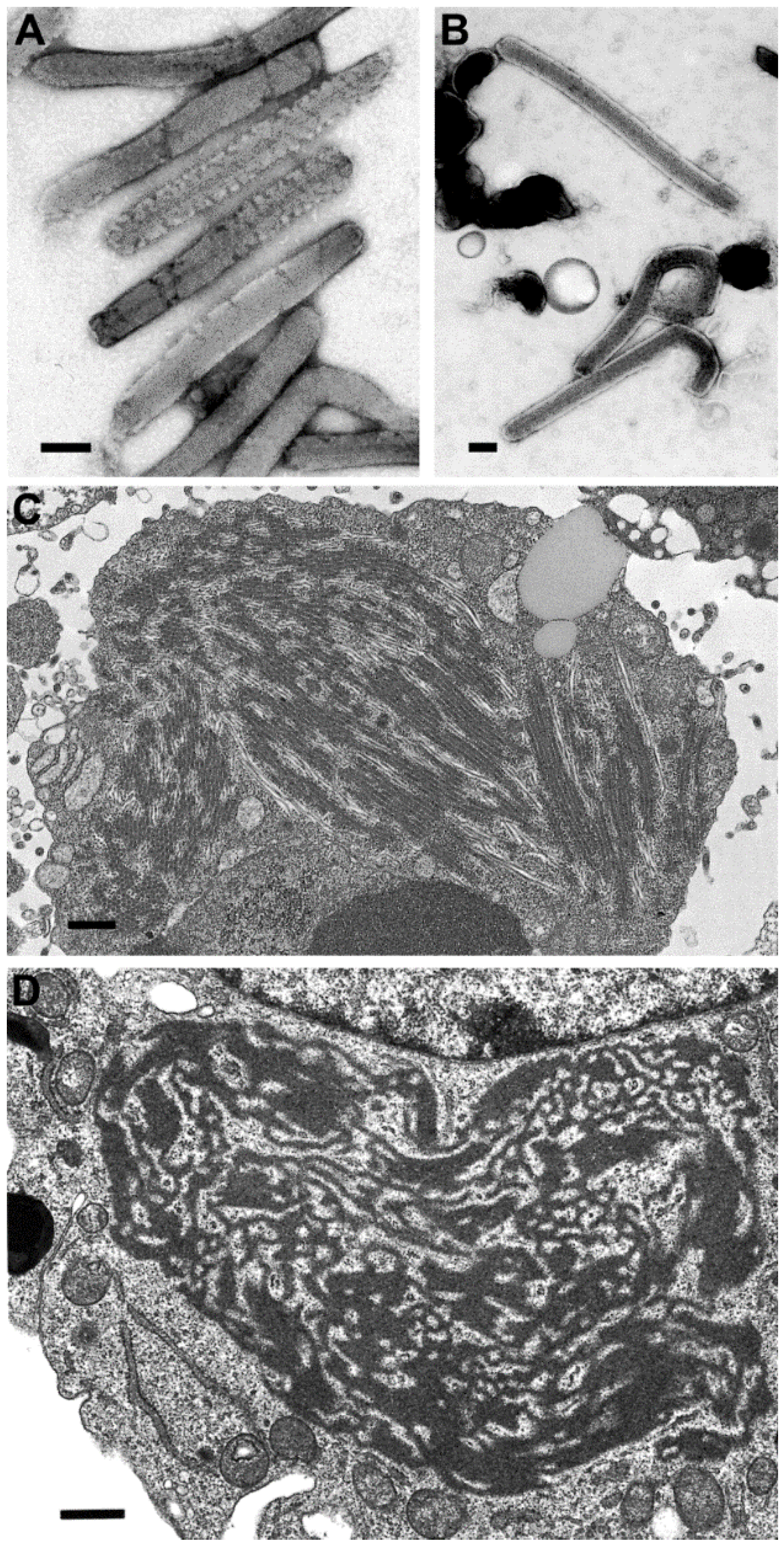
There have been differences reported in the morphologic features of Ebola and Marburg viruses. First, the lengths of the virus particles of the two genera vary, although there have been different lengths reported. For instance, there have been reports of 665 nm, 790 nm, and 860 nm lengths for Marburg viru
s, and 805 nm, 970 nm, and 1200 nm for Ebola virus, but clearly Ebola viruses are longer [
21,
22,
23] (
Figure 5A,B). A second difference is found in the ultrastructure of the intermediate inclusion of the viruses. Ebola virus inclusions have distinct preformed nucleocapsids mixed with lighter-staining matrix material and, at times, naked nucleocapsids are present (
Figure 5C). The inclusions in Marburg virus infections begin with light-staining nascent viral material, which increases in electron density as the infection progresses, and has 45–60 nm spheres of inclusion material surrounding the inclusion. Later, the intermediate inclusions show a dispersal of material and a loss of the spheres [
23,
24] (
Figure 5D).
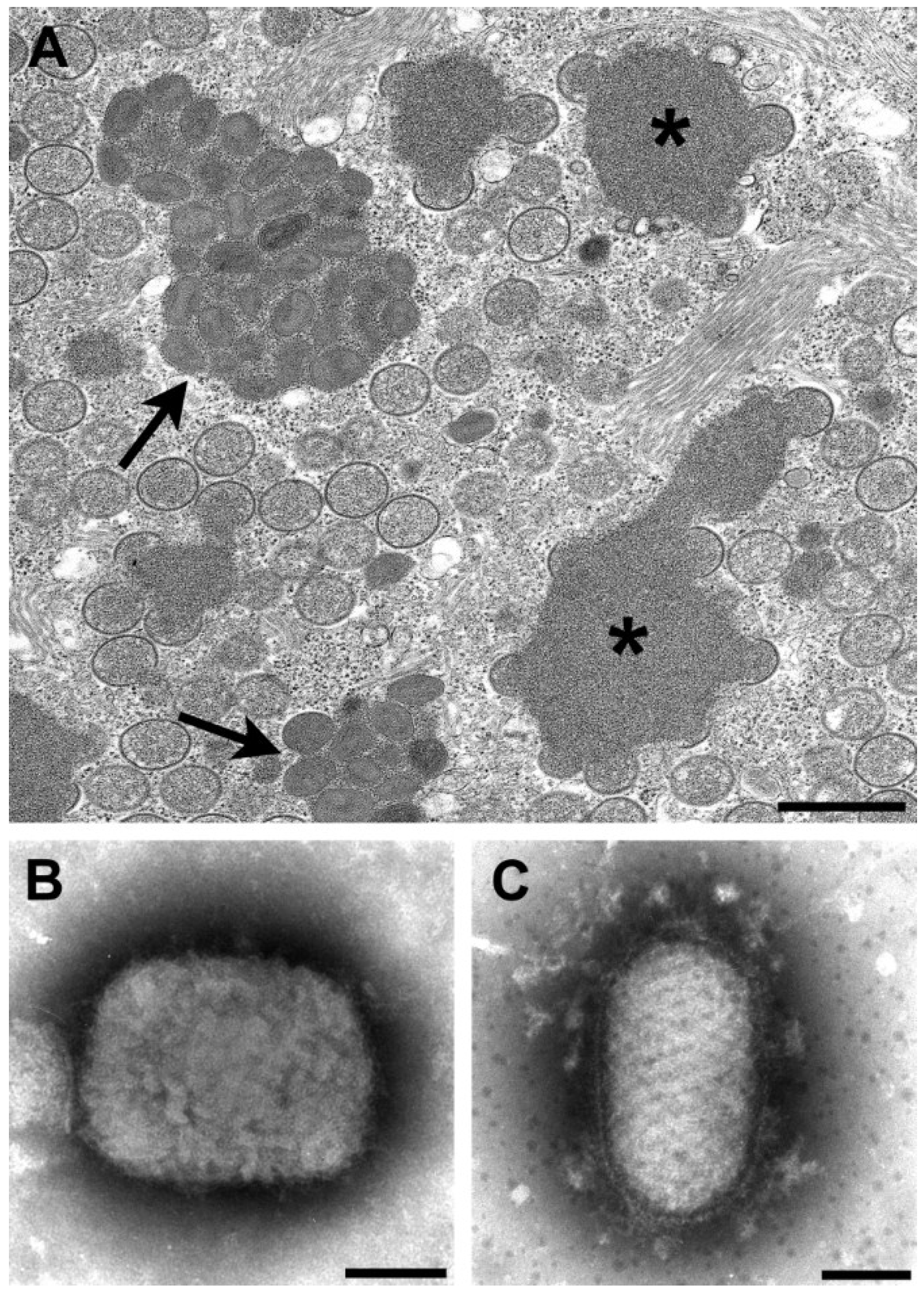
Figure 5.
Negative stain images of Marburg virus (A) and Ebola virus (B), illustrating that Ebola virus has a longer length than Marburg virus. Bars, 100 nm; (C) Thin section image of a large inclusion in an Ebola virus-infected cell. Bar, 500 nm; (D) Intermediate inclusion in the cytoplasm of a Marburg virus-infected cell. Bar, 580 nm. (Figure A, courtesy of Russell Regnery, Centers for Disease Control and Prevention; Figure D, courtesy of Thomas Geisbert, United States Army Medical Research Institute of Infectious Diseases, Frederick, MD, USA.)
Figure 5.
Negative stain images of Marburg virus (A) and Ebola virus (B), illustrating that Ebola virus has a longer length than Marburg virus. Bars, 100 nm; (C) Thin section image of a large inclusion in an Ebola virus-infected cell. Bar, 500 nm; (D) Intermediate inclusion in the cytoplasm of a Marburg virus-infected cell. Bar, 580 nm. (Figure A, courtesy of Russell Regnery, Centers for Disease Control and Prevention; Figure D, courtesy of Thomas Geisbert, United States Army Medical Research Institute of Infectious Diseases, Frederick, MD, USA.)
2.6. Bunyaviridae
The family Bunyaviridae contains four genera that can infect humans—Orthobunyavirus, Nairovirus, Phlebovirus, and Hantavirus. Viruses contain single-stranded RNA with three RNA segments that are negative sense, with the exception of phleboviruses which have one ambisense segment. These are zoonotic viruses, where each virus is associated with a specific vector or natural reservoir, including mosquitoes, ticks, sand flies, and rodents. Patients will usually have a hemorrhagic syndrome which is characterized by fever, increased capillary permeability, leukopenia and thrombocytopenia.
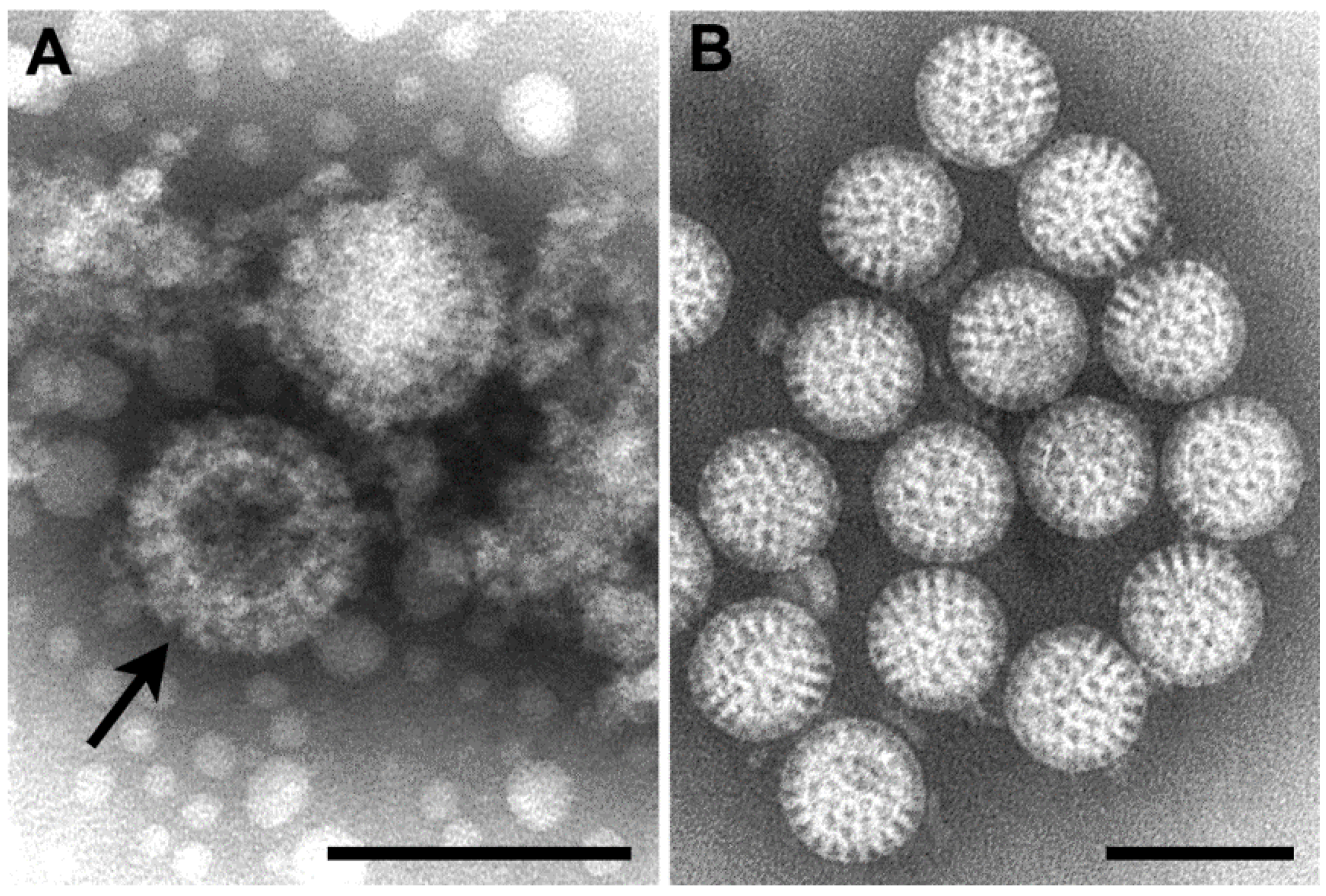
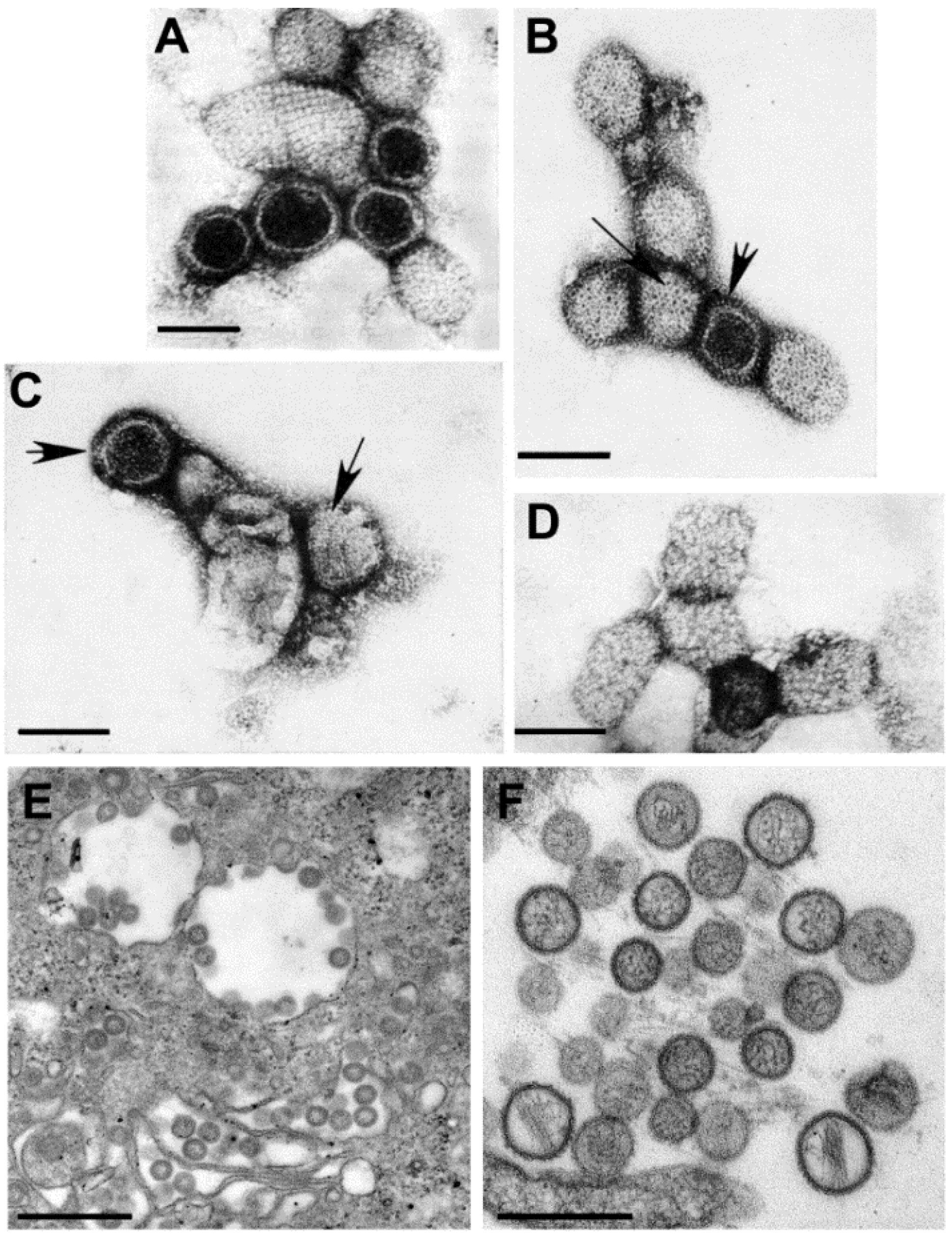
Figure 6.
(A–D) Negative stain EM preparations. (A) Hantaan virus particles (genus Hantavirus) showing a grid-like surface pattern. Bar, 100 nm; (B) Rift Valley fever virus (genus Phlebovirus) exhibiting subunits with a central hole (long arrow) and showing the regularly spaced subunits (short arrow). Bar, 100 nm; (C) Crimean-Congo hemorrhagic fever virus (genus Nairovirus) having small surface subunits (long arrow) which appear as a peripheral fringe (short arrow). Bar, 100 nm; (D) Anhemi virus (genus Orthobunyavirus) with knob-like surface structures. Bar, 100 nm; (E,F) Thin section EM preparations; (E) Virions have moderately dense centers and accumulate in the cisternae of the Golgi complex of a Rift Valley fever virus-infected cell. Bar, 500 nm; (F) Extracellular Sin Nombre virus particles (genus Hantavirus) have a variety of sizes and internal cores composed of thin thread-like material. Bar, 500 nm. (Figures A–D, courtesy of Mary Lane Martin, Centers for Disease Control and Prevention; Figure E, courtesy of Frederick A. Murphy, Centers for Disease Control and Prevention.)
Figure 6.
(A–D) Negative stain EM preparations. (A) Hantaan virus particles (genus Hantavirus) showing a grid-like surface pattern. Bar, 100 nm; (B) Rift Valley fever virus (genus Phlebovirus) exhibiting subunits with a central hole (long arrow) and showing the regularly spaced subunits (short arrow). Bar, 100 nm; (C) Crimean-Congo hemorrhagic fever virus (genus Nairovirus) having small surface subunits (long arrow) which appear as a peripheral fringe (short arrow). Bar, 100 nm; (D) Anhemi virus (genus Orthobunyavirus) with knob-like surface structures. Bar, 100 nm; (E,F) Thin section EM preparations; (E) Virions have moderately dense centers and accumulate in the cisternae of the Golgi complex of a Rift Valley fever virus-infected cell. Bar, 500 nm; (F) Extracellular Sin Nombre virus particles (genus Hantavirus) have a variety of sizes and internal cores composed of thin thread-like material. Bar, 500 nm. (Figures A–D, courtesy of Mary Lane Martin, Centers for Disease Control and Prevention; Figure E, courtesy of Frederick A. Murphy, Centers for Disease Control and Prevention.)
![Viruses 06 04902 g006]()
In negative stain preparations, Martin,
et al. [
25] were able to distinguish among the genera of the family, according to the surface arrangement of structural units. Hantaviruses have the most distinct structure, where the surface units are arranged in a square, grid-like pattern (
Figure 6A). Phleboviruses, which have recently incorporated the genus
Uukuvirus, have a surface structure formed of subunits with a distinct central hole (
Figure 6B). The surface of nairoviruses have very small morphologic units (
Figure 6C), and orthobunyaviruses exhibit either knob-like surface units or are indistinct (
Figure 6D).
By thin section EM, the virions of most bunyaviruses have a fairly homogeneous core and small spikes on the virus surface are visible in some preparations. The viruses mature by budding upon the membranes of the Golgi complex and can accumulate in the Golgi cisternae or extracellularly [
26] (
Figure 6E). The viruses in the genus
Hantavirus, however, have a core consisting of thin threads of ribonucleoproteins (
Figure 6F). In addition, in cell cultures hantaviruses can mature by budding from the plasma membranes of infected cells, and are associated with tubular projections [
27,
28].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}