Evidence of Epstein-Barr Virus Association with Gastric Cancer and Non-Atrophic Gastritis


Abstract
:1. Introduction
2. Results and Discussion
2.1. Sample Description
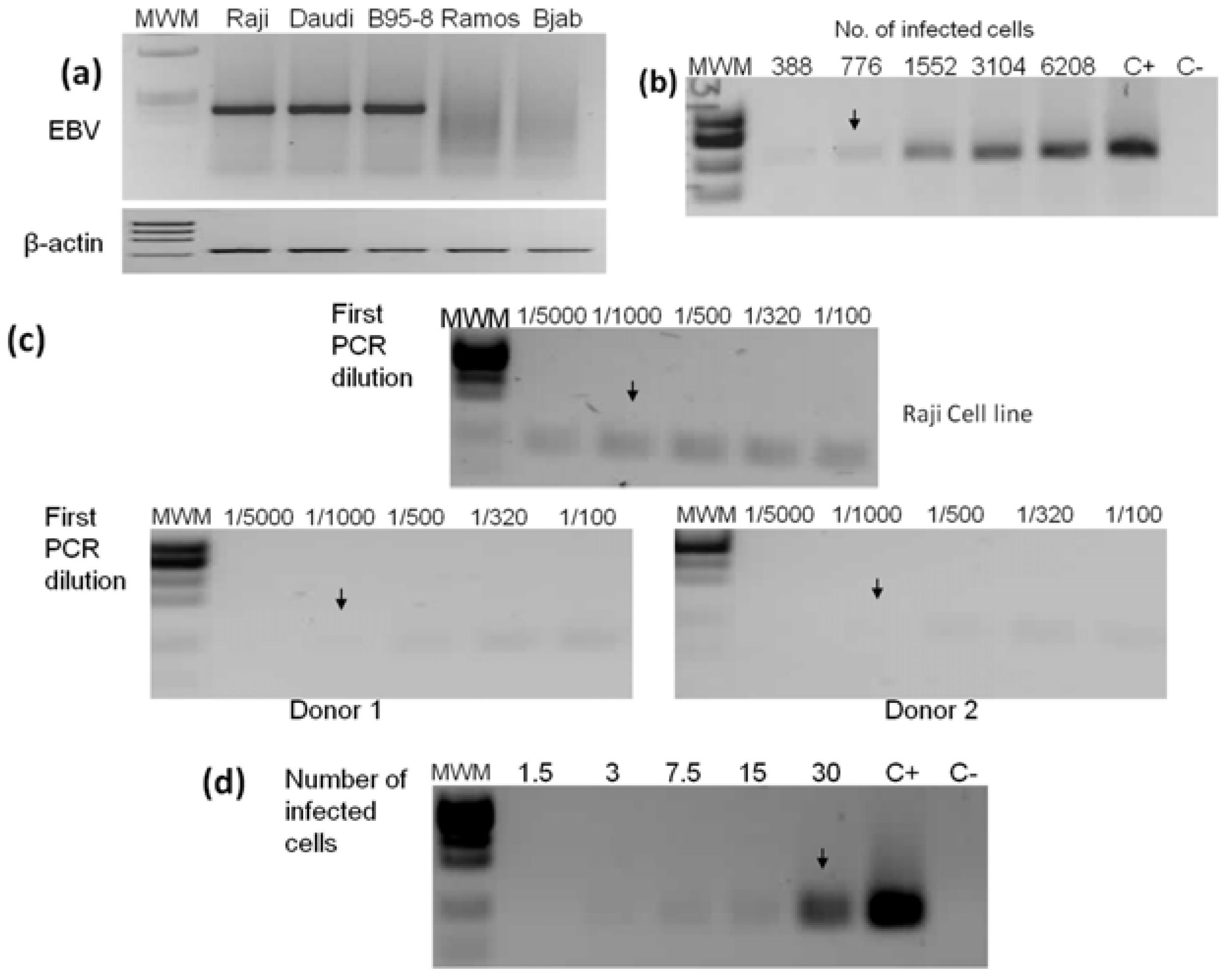
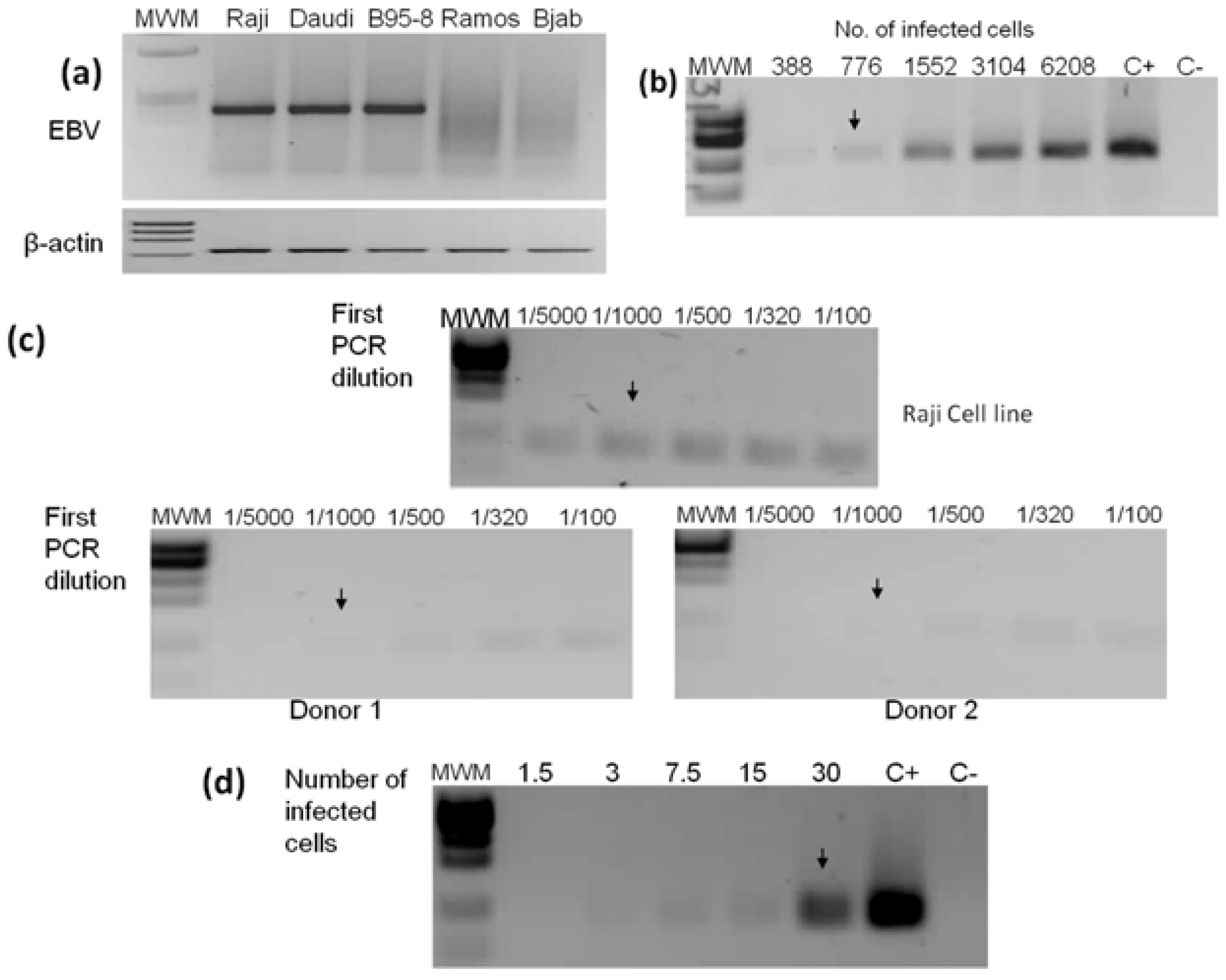
2.2. PCR Detection Limits
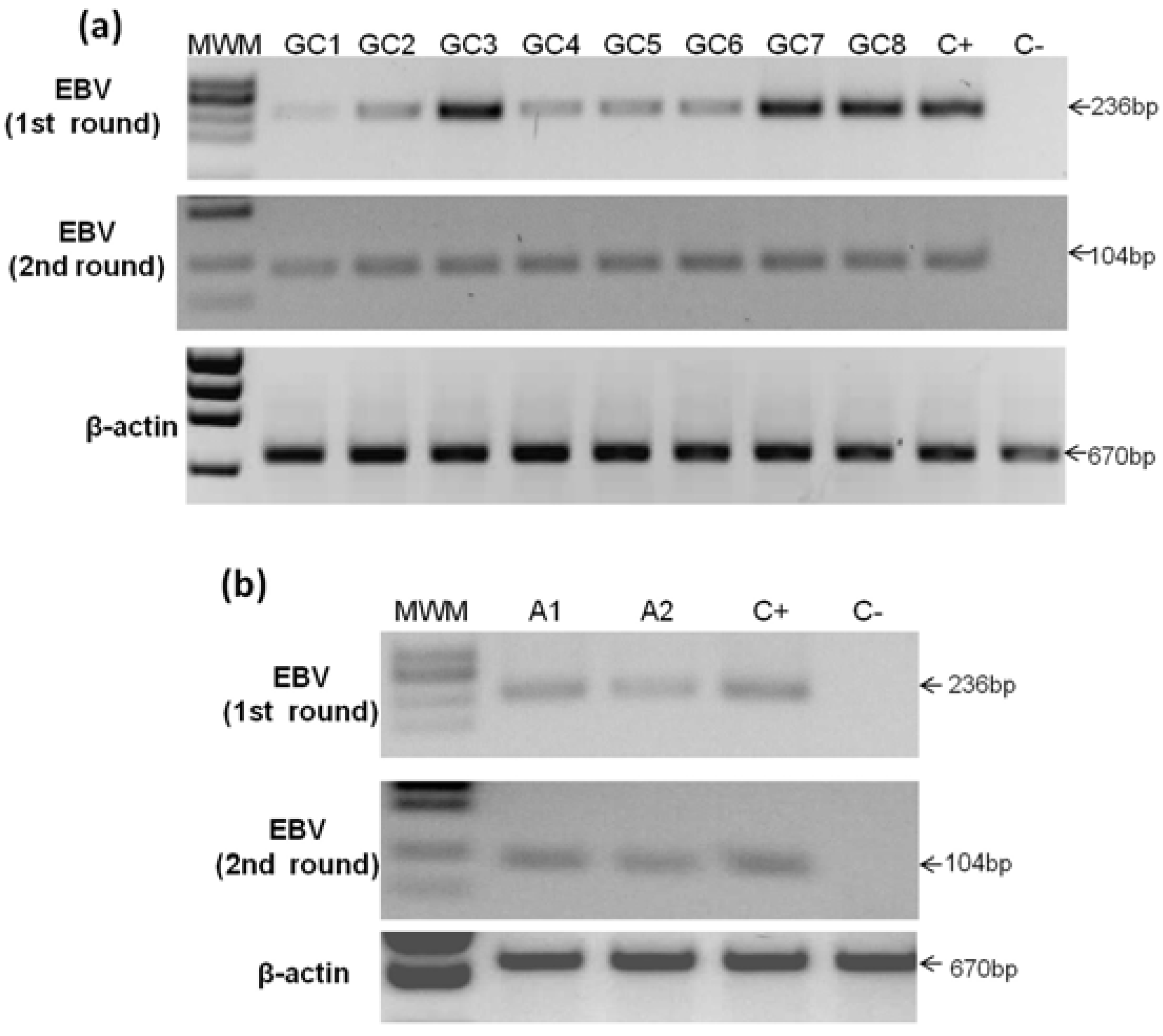
2.3. Detection of EBV in Gastric Cancer
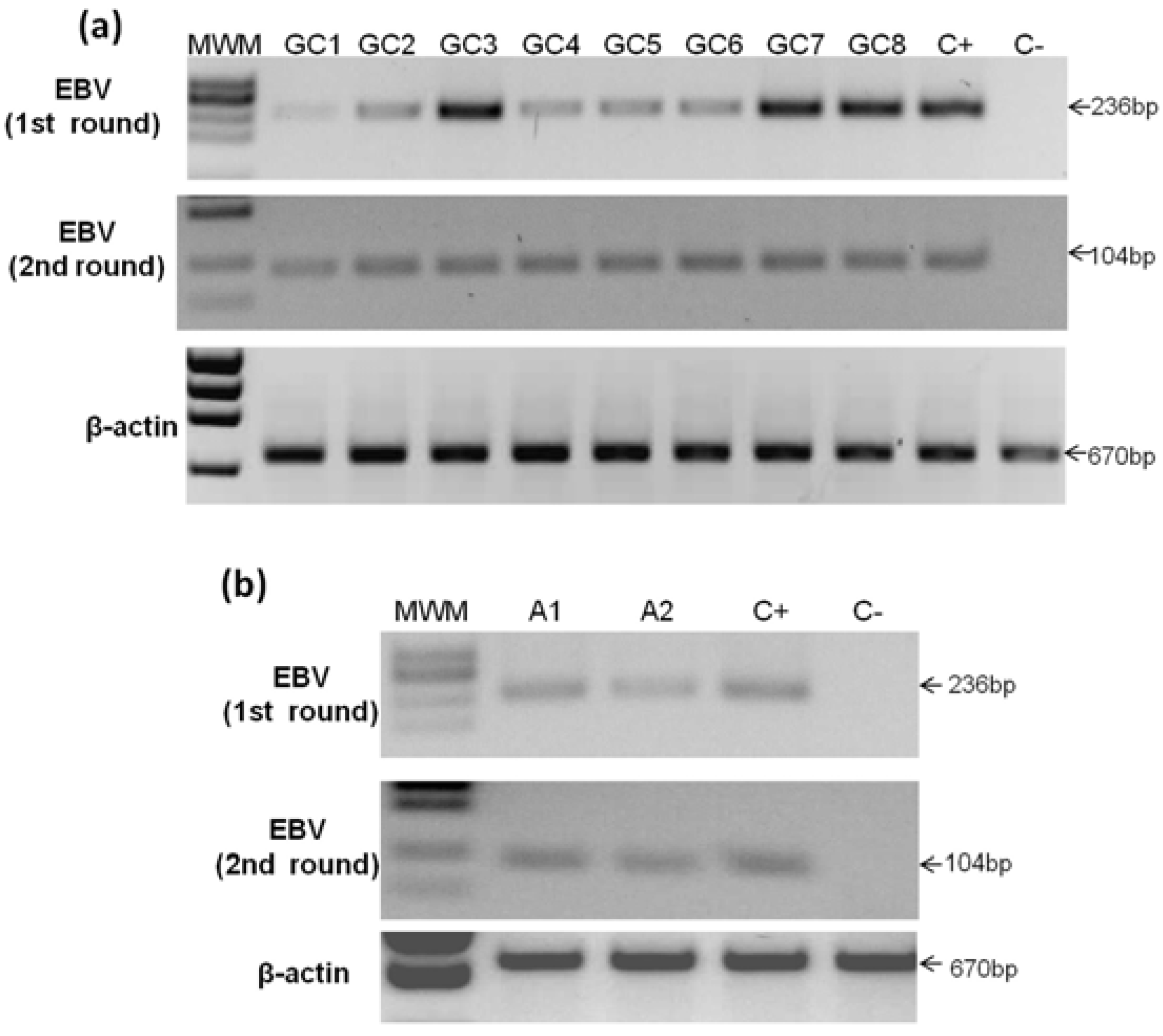

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No sample | Lauren | WHO | Other histological patterns | Lymphocyte infiltrate −/+++ | Age | Gender | % of tumoral cells | Site |
|---|---|---|---|---|---|---|---|---|
| GC1 | Mixed (Diffuse and intestinal) | Tubular Mucinous | Lace pattern with clear cells (LEL-like) | +++ | 87 | Male | 80 | Body |
| GC2 | Intestinal | Tubular | Cribriform | + | 78 | Female | 10 | Body and Antrum |
| GC3 | Diffuse | Signet ring cells | None | ++ | 38 | Male | 30 | Body and Antrum |
| GC4 | Intestinal | Tubular | Cribriform with clear cells | + | 72 | Female | 60 | Body |
| GC5 | Diffuse | Signet ring cells | None | + | 74 | Female | 70 | Antrum |
| GC6 | Intestinal | Tubular | None | + | 72 | Male | 25 | Body |
| GC7 | Intestinal | Tubular | Cribriform | ++ | 52 | Female | 60 | Body and Antrum |
| GC8 | Intestinal | Tubular | Lace pattern | + | 57 | Male | 20 | Antrum |
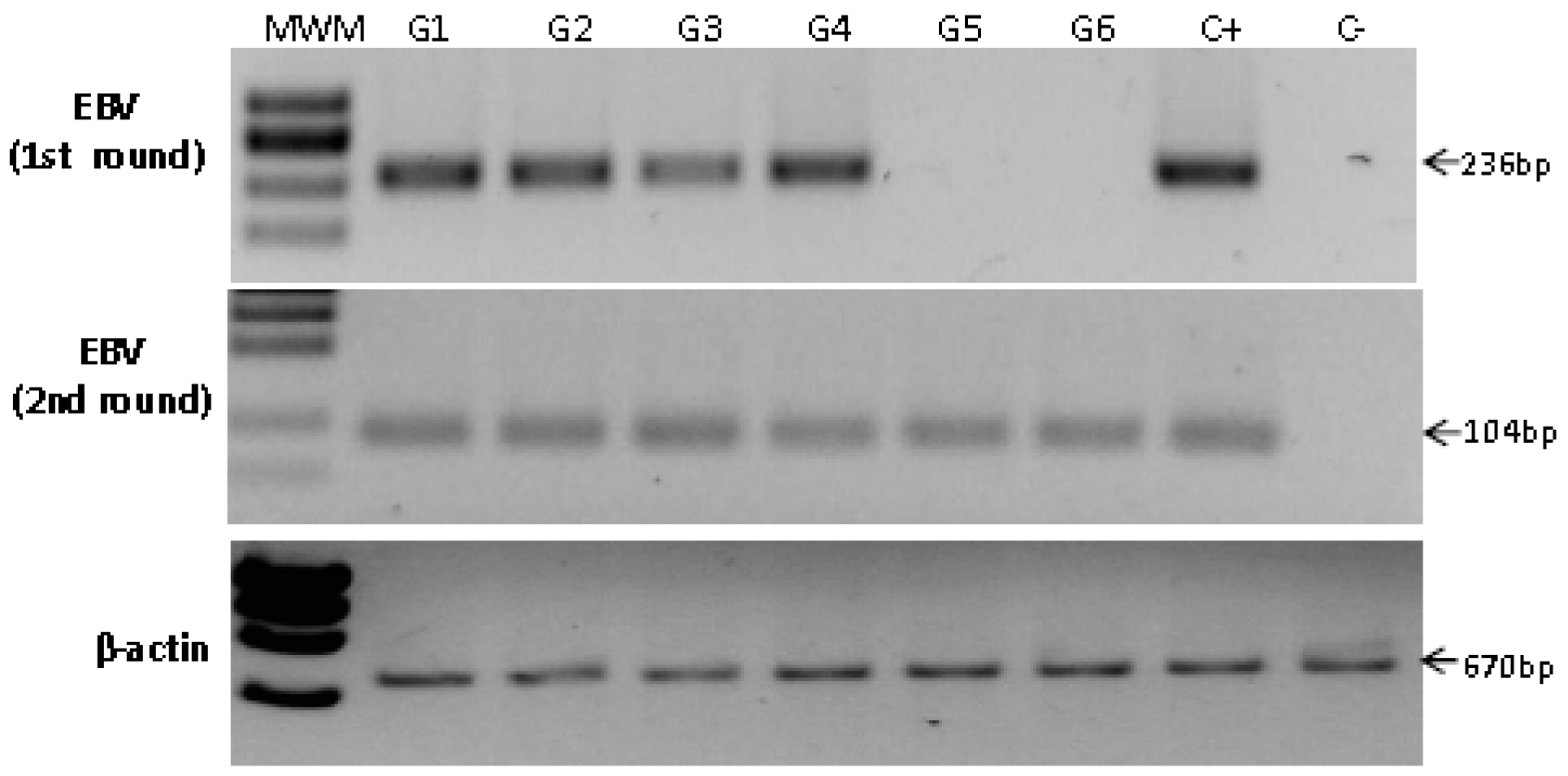
2.4. Detection of EBV in Non-Atrophic Gastritis
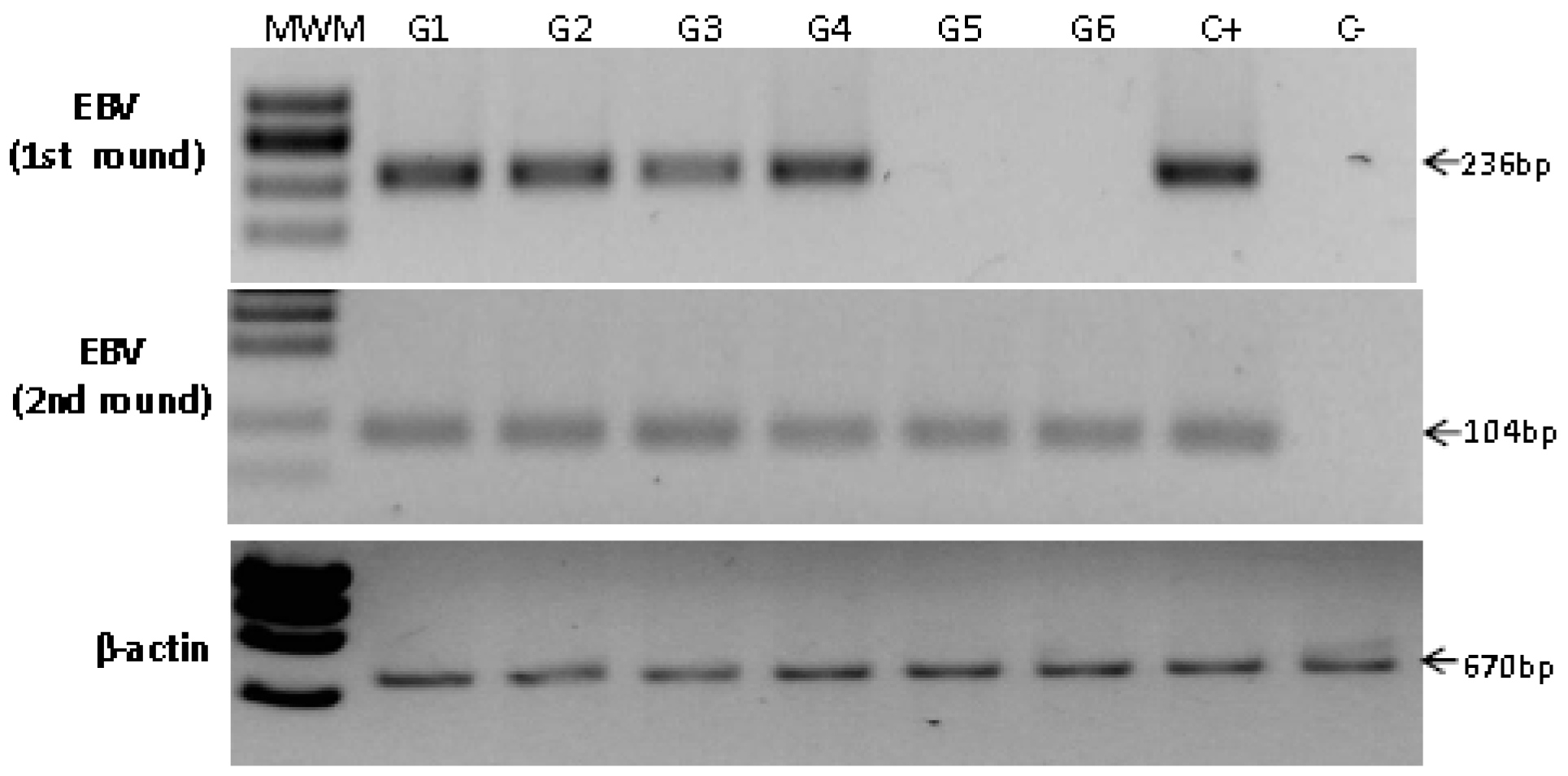
| Sample | Age | Gender | Inflammation −/+++ | Activity −/+++ | Site | Hp | PCR | ISH signal |
|---|---|---|---|---|---|---|---|---|
| G1 | 35 | Male | ++ | ++ | Body | − | First | Lymphoid infiltrate |
| G2 | 25 | Male | +++ | ++ | Body | + | First | Lymphoid infiltrate |
| G3 | 64 | Male | ++ | ++ | Body | − | First | Epithelial cell |
| G4 | 61 | Female | +++ | − | Antrum | + | First | Lymphoid infiltrate |
| G5 | 44 | Female | +++ | − | Antrum | NA | Nested | Epithelial cell & Lymphoid infiltrate |
| G6 | 23 | Male | +++ | − | Antrum | + | Nested | Lymphoid infiltrate |
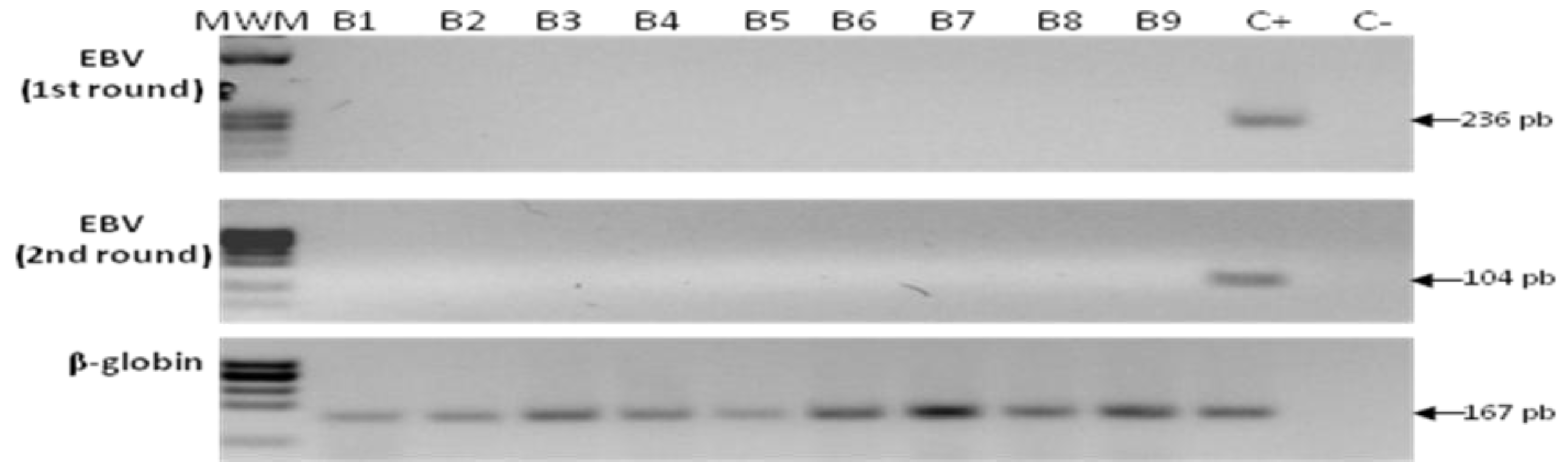
2.5. Detection of EBV in Samples from Gastric Bypass Surgeries
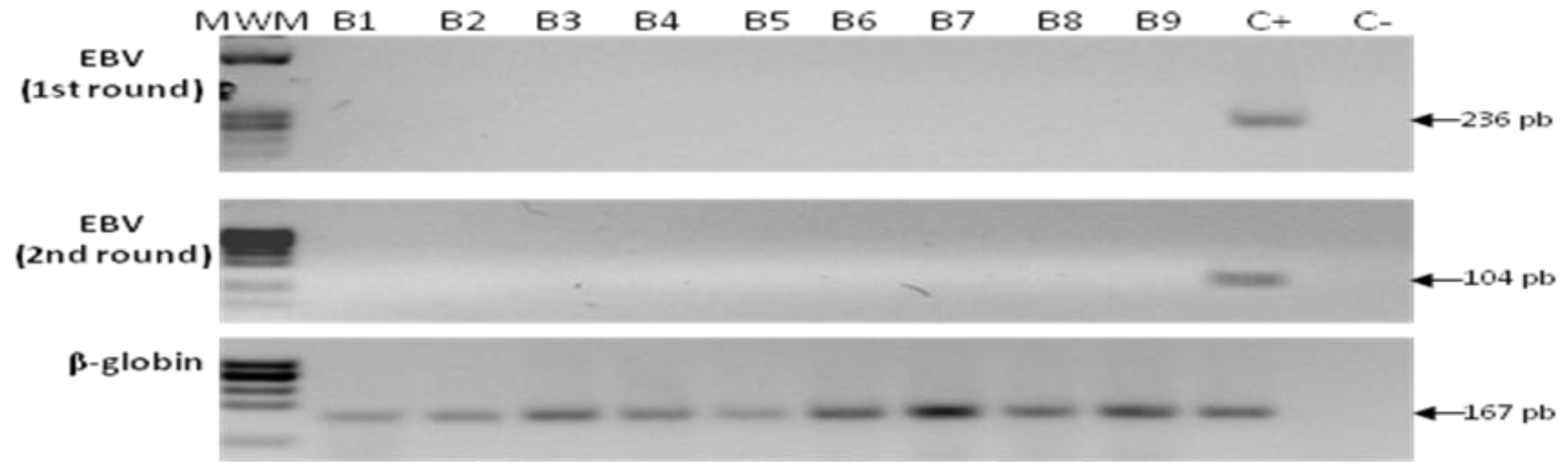
2.6. Sequencing
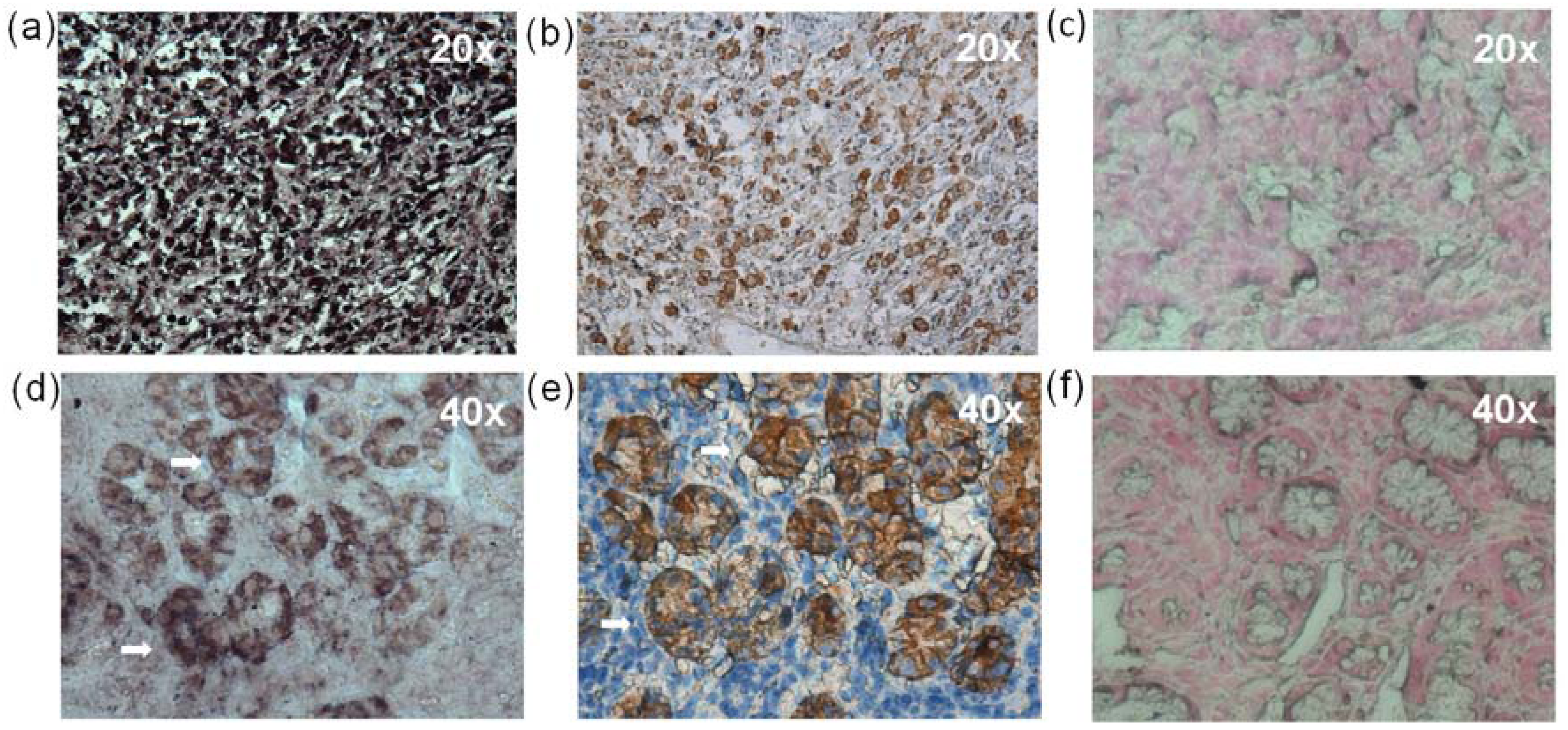
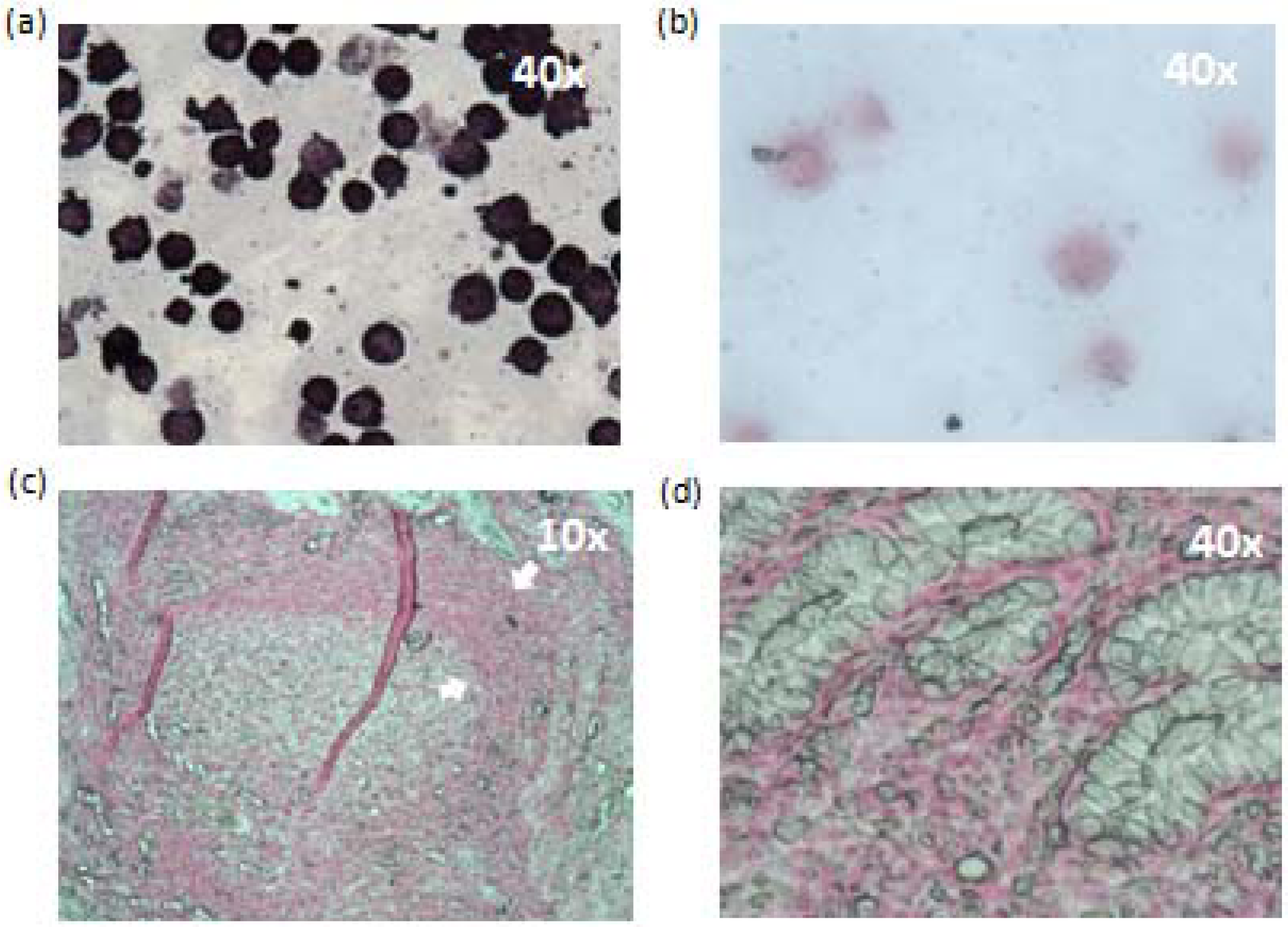
2.7. In situ Hybridization (ISH) and Immunohistochemistry (IHC)
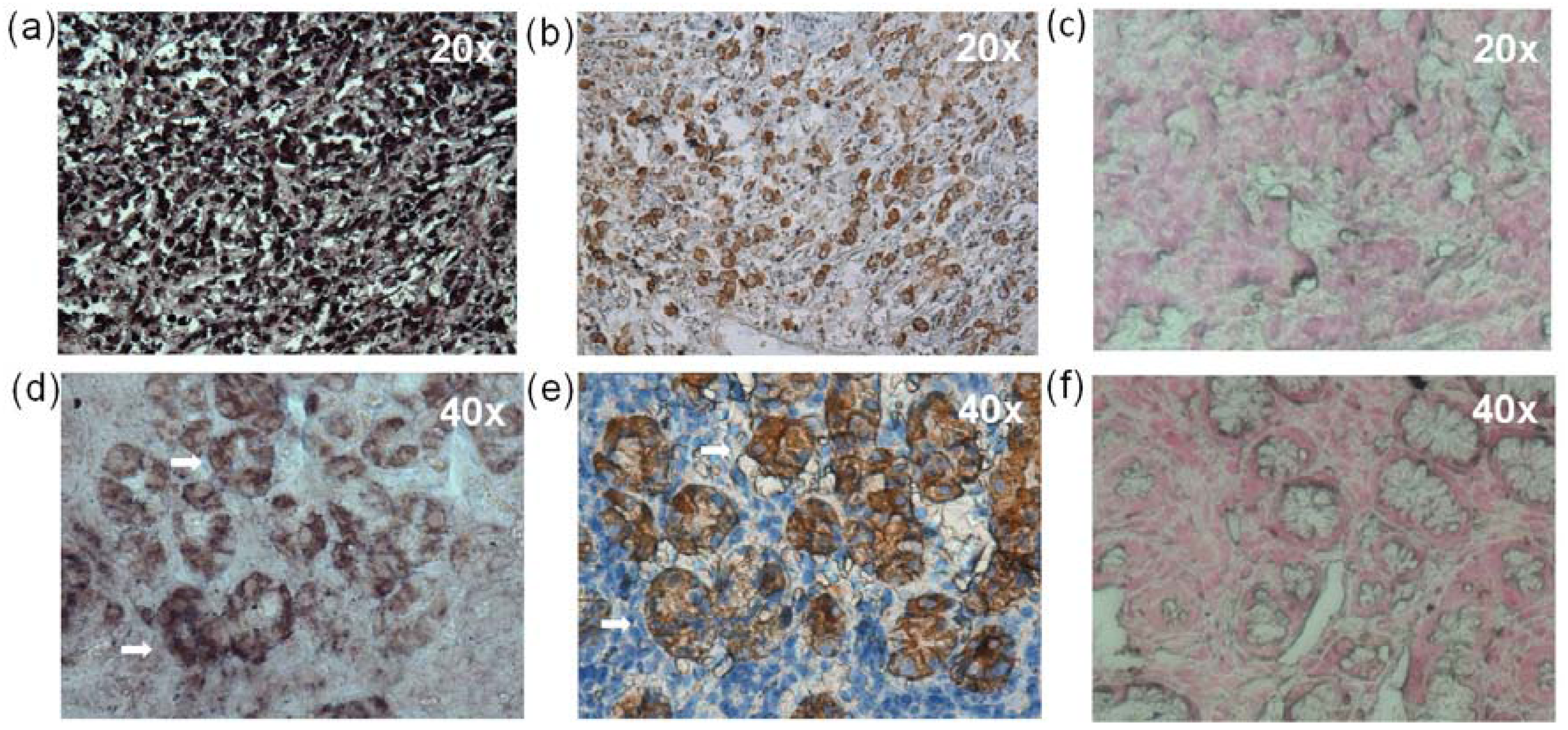
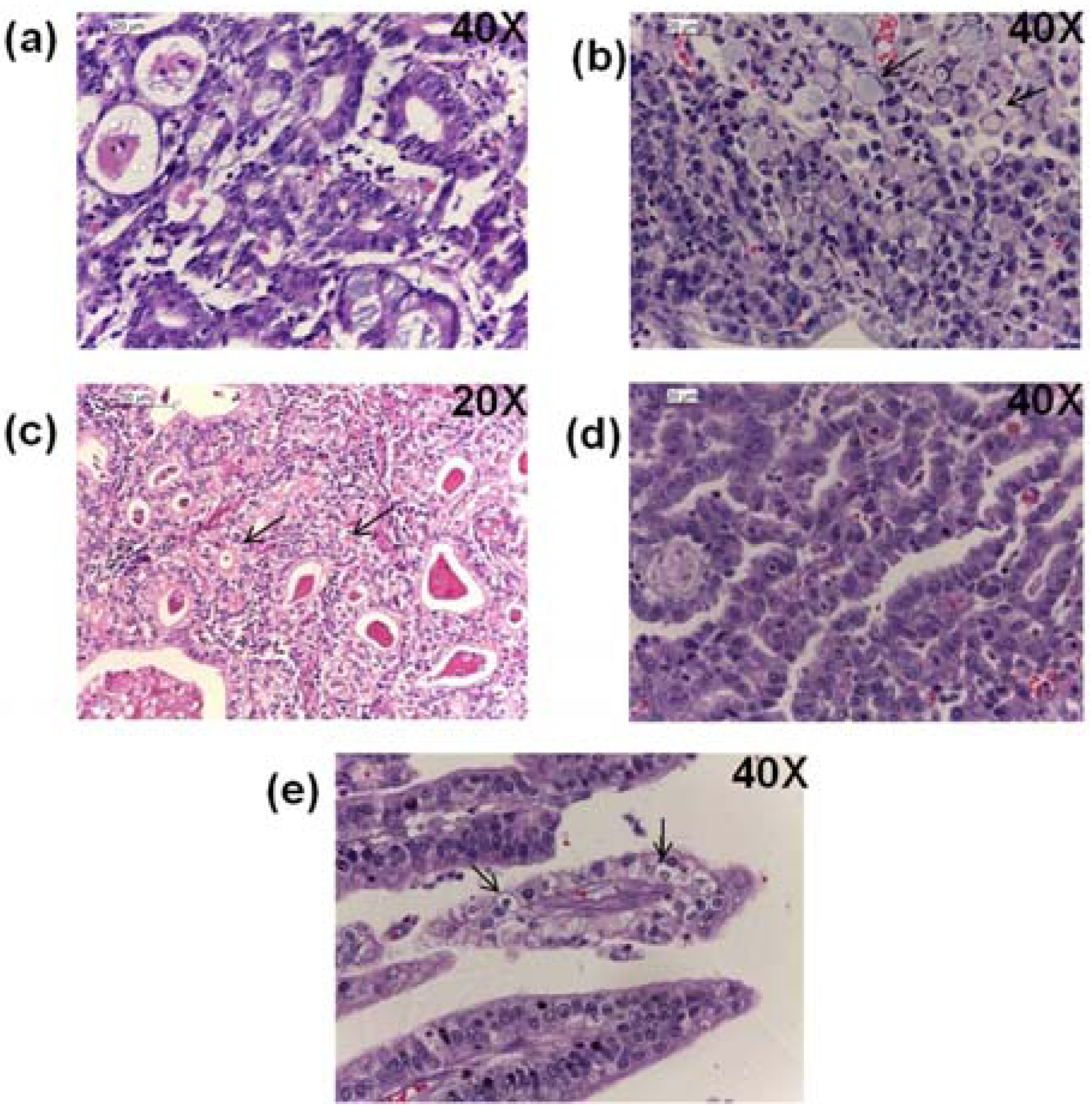
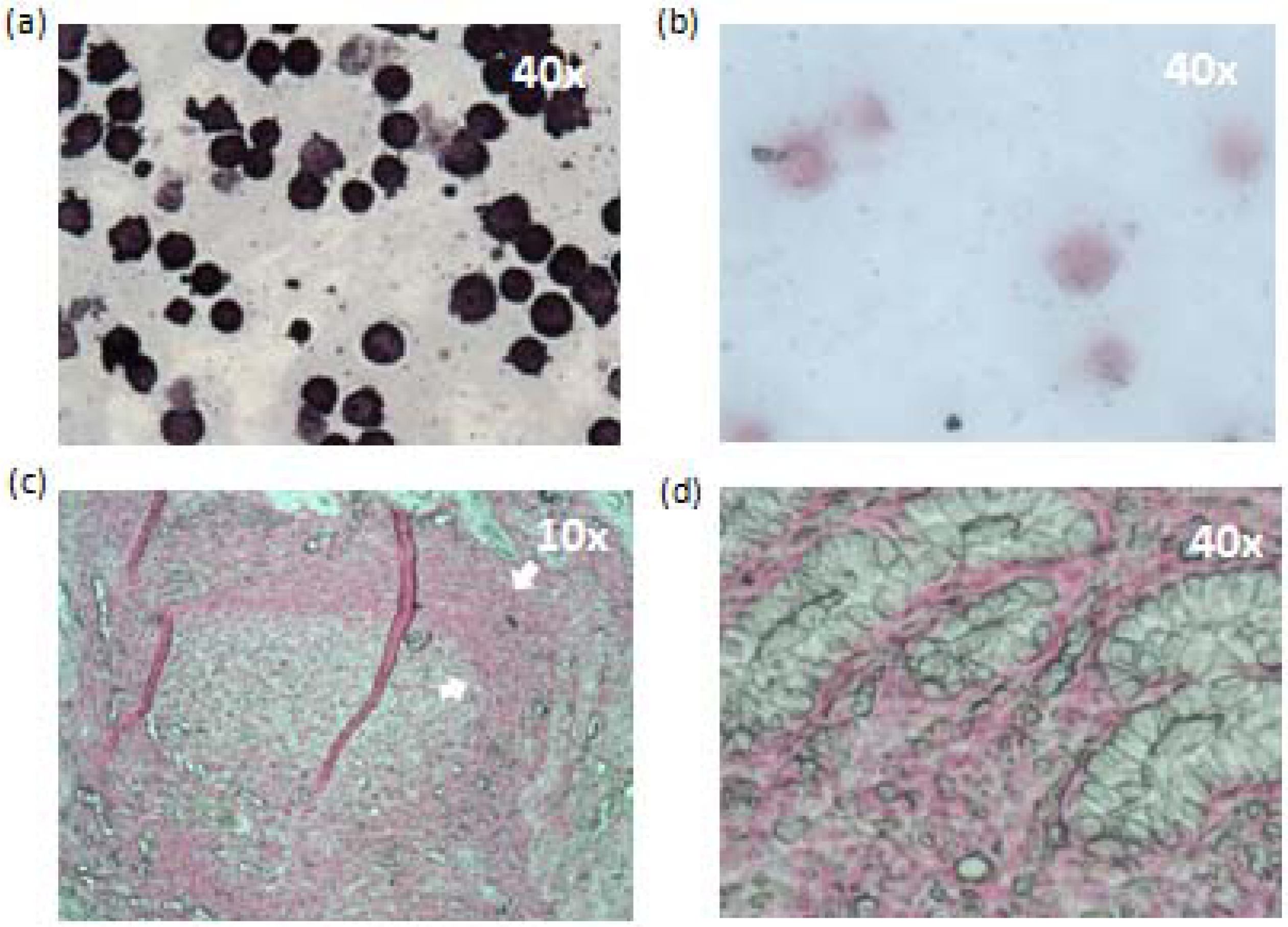
2.8. Histopathological Findings
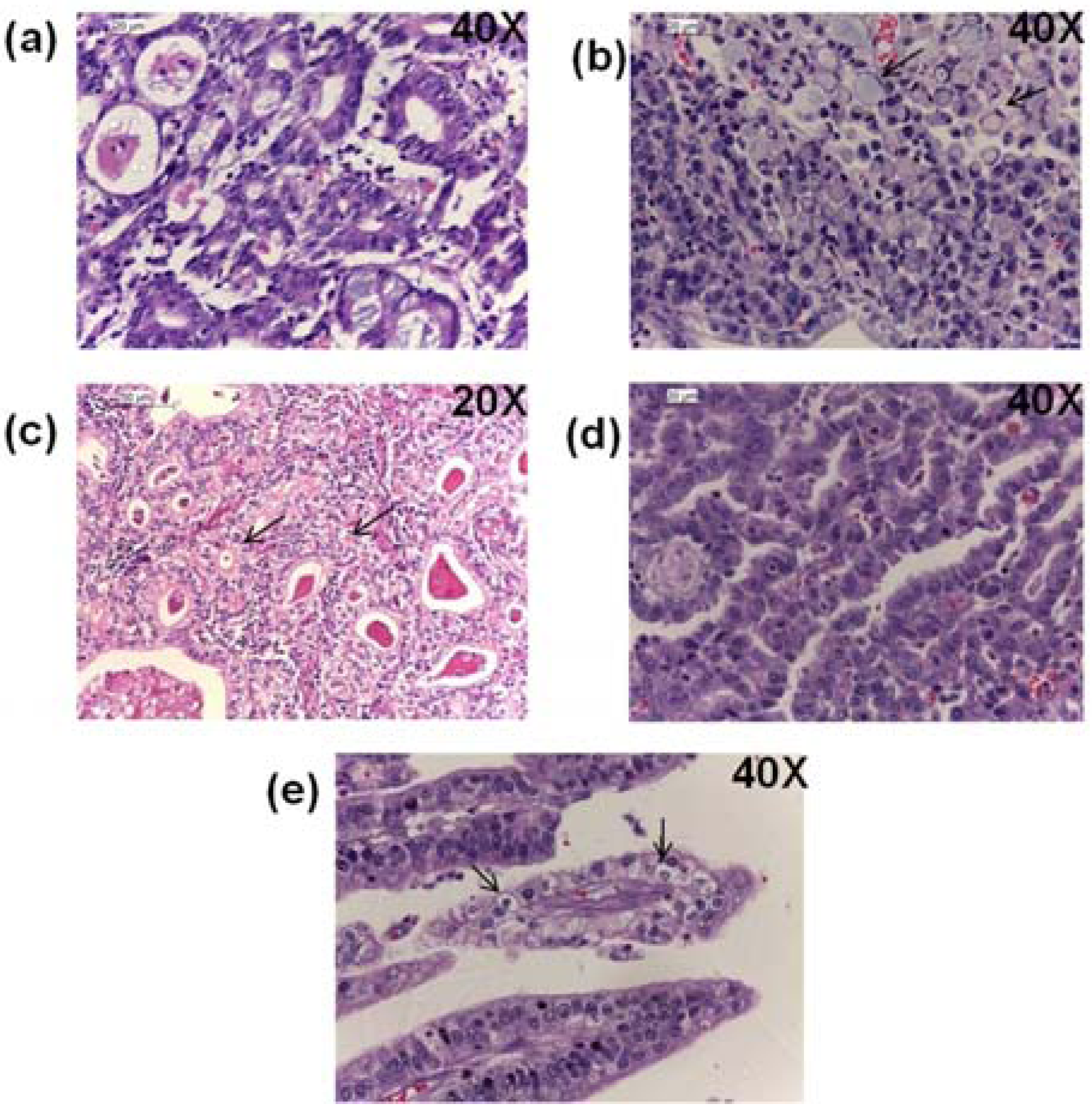
2.9. Discussion
3. Experimental Section
3.1. Cell Lines
3.2. Patients and Samples
| Hospital | Gastric Sample | Gender | Mean Age | Condition | |
|---|---|---|---|---|---|
| Male * | Female * | ||||
| GC Mexico City: Oncology Hospital, General Hospital and National Institute of Cancer | 34 Diffuse | 23 | 31 | 61 | Fresh and frozen |
| 17 Intestinal | |||||
| 4 Mixed | |||||
| 44 Non-tumor controls | |||||
| GC Merida, Yucatan: Ignacio Garcia Tellez Hospital. | 10 Diffuse | 9 | 11 | 62.6 | Paraffin embedded |
| 10 Intestinal | |||||
| Non-tumor controls, Mexico City: Bernardo Sepulveda Specialties Hospital, Oncology Hospital, General Hospital, National Institute of Cancer and Carlos McGregor Hospital | 99 Non-tumor controls | 43 | 43 | 61.6 | Frozen |
| Non-tumor controls Merida Yucatan: Ignacio García Tellez, Hospital | 4 Non-tumor controls | 2 | 2 | 71.5 | Paraffin embedded |
| Non-atrophic gastritis, México City: Bernardo Sepulveda Specialties Hospital and Carlos McGregor Hospital. | 49 NAG | 26 | 23 | 49.5 | Frozen |
| Non-atrophic gastritis, Merida Yucatan: Ignacio Garcia Tellez Hospital | 27 NAG | 10 | 17 | 52.6 | Paraffin embedded |
3.3. Histopathological Examination
3.4. DNA Purification
3.5. EBV Detection
3.6. Sequencing
3.7. In Situ Hybridization
3.8. Immunohistochemistry (IHC)
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Appendices
| Muestra | H.pilory | PMN cells | MN cells | Atrophic Gastritis | Metaplasia | Dysplasia | Additional observations |
|---|---|---|---|---|---|---|---|
| SGB 001 | + | 0 | ++ | 0 | 0 | 0 | Follicular gastritis |
| SGB 002 | + | 0 | ++ | 0 | 0 | 0 | Follicular gastritis |
| SGB 003 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 004 | + | 0 | ++ | 0 | 0 | 0 | None |
| SGB 005 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 006 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 007 | + | + | ++ | 0 | 0 | 0 | None |
| SGB 008 | 0 | 0 | 0 | 0 | 0 | 0 | None |
| SGB 009 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 010 | 0 | 0 | ++ | 0 | 0 | 0 | None |
| SGB 011 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 012 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 013 | 0 | + | ++ | 0 | 0 | 0 | None |
| SGB 014 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 015 | 0 | ++ | +++ | +++ | ++ | 0 | Neuroendocrine cell hyperplasia |
| SGB 016 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 017 | 0 | 0 | + | 0 | 0 | 0 | None |
| SGB 018 | 0 | 0 | ++ | 0 | 0 | 0 | None |
| SGB 019 | 0 | 0 | 0 | 0 | 0 | 0 | None |
Authors’ Contribution
Conflicts of Interest
References and Notes
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef]
- Lauren, P. The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma. An attempt at a histo-clinical classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar]
- Correa, P.; Piazuelo, M.B.; Camargo, M.C. Etiopathogenesis of gastric cancer. Scand. J. Surg. 2006, 95, 218–224. [Google Scholar]
- Crew, K.D.; Neugut, A.I. Epidemiology of gastric cancer. World J. Gastroenterol. 2006, 12, 354–362. [Google Scholar]
- You, W.C.; Brown, L.M.; Zhang, L.; Li, J.Y.; Jin, M.L.; Chang, Y.S.; Ma, J.L.; Pan, K.F.; Liu, W.D.; Hu, Y.; et al. Randomized double-blind factorial trial of three treatments to reduce the prevalence of precancerous gastric lesions. J. Natl. Cancer Inst. 2006, 98, 974–983. [Google Scholar] [CrossRef]
- Crawford, D.H. Biology and disease associations of Epstein-Barr virus. Philos. Trans. R Soc. Lond. B Biol. Sci. 2001, 356, 461–473. [Google Scholar] [CrossRef]
- Camargo, M.C.; Murphy, G.; Koriyama, C.; Pfeiffer, R.M.; Kim, W.H.; Herrera-Goepfert, R.; Corvalan, A.H.; Carrascal, E.; Abdirad, A.; Anwar, M.; et al. Determinants of Epstein-Barr virus-positive gastric cancer: An international pooled analysis. Br. J. Cancer 2011, 105, 38–43. [Google Scholar] [CrossRef]
- Murphy, G.; Pfeiffer, R.; Camargo, M.C.; Rabkin, C.S. Meta-analysis shows that prevalence of Epstein-Barr virus-positive gastric cancer differs based on sex and anatomic location. Gastroenterology 2009, 137, 824–833. [Google Scholar] [CrossRef]
- Cardenas-Mondragon, M.G.; Carreon-Talavera, R.; Camorlinga-Ponce, M.; Gomez-Delgado, A.; Torres, J.; Fuentes-Panana, E.M. Epstein Barr virus and Helicobacter pylori co-infection are positively associated with severe gastritis in pediatric patients. PLoS One 2013, 8, e62850. [Google Scholar]
- Ryan, J.L.; Morgan, D.R.; Dominguez, R.L.; Thorne, L.B.; Elmore, S.H.; Mino-Kenudson, M.; Lauwers, G.Y.; Booker, J.K.; Gulley, M.L. High levels of Epstein-Barr virus DNA in latently infected gastric adenocarcinoma. Lab. Invest. 2009, 89, 80–90. [Google Scholar] [CrossRef]
- Camorlinga-Ponce, M.; Flores-Luna, L.; Lazcano-Ponce, E.; Herrero, R.; Bernal-Sahagun, F.; Abdo-Francis, J.M.; Aguirre-Garcia, J.; Munoz, N.; Torres, J. Age and severity of mucosal lesions influence the performance of serologic markers in Helicobacter pylori-associated gastroduodenal pathologies. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2498–2504. [Google Scholar] [CrossRef]
- Adams, A. Replication of latent Epstein-Barr virus genomes in Raji cells. J. Virol. 1987, 61, 1743–1746. [Google Scholar]
- Gulley, M.L.; Eagan, P.A.; Quintanilla-Martinez, L.; Picado, A.L.; Smir, B.N.; Childs, C.; Dunn, C.D.; Craig, F.E.; Williams, J.W., Jr.; Banks, P.M. Epstein-Barr virus DNA is abundant and monoclonal in the Reed-Sternberg cells of Hodgkin’s disease: Association with mixed cellularity subtype and Hispanic American ethnicity. Blood 1994, 83, 1595–1602. [Google Scholar]
- Herrera-Goepfert, R.; Akiba, S.; Koriyama, C.; Ding, S.; Reyes, E.; Itoh, T.; Minakami, Y.; Eizuru, Y. Epstein-Barr virus-associated gastric carcinoma: Evidence of age-dependence among a Mexican population. World J. Gastroenterol. 2005, 11, 6096–6103. [Google Scholar]
- Herrera-Goepfert, R.; Reyes, E.; Hernandez-Avila, M.; Mohar, A.; Shinkura, R.; Fujiyama, C.; Akiba, S.; Eizuru, Y.; Harada, Y.; Tokunaga, M. Epstein-Barr virus-associated gastric carcinoma in Mexico: Analysis of 135 consecutive gastrectomies in two hospitals. Mod. Pathol. 1999, 12, 873–878. [Google Scholar]
- Corvalan, A.; Koriyama, C.; Akiba, S.; Eizuru, Y.; Backhouse, C.; Palma, M.; Argandona, J.; Tokunaga, M. Epstein-Barr virus in gastric carcinoma is associated with location in the cardia and with a diffuse histology: A study in one area of Chile. Int. J. Cancer 2001, 94, 527–530. [Google Scholar] [CrossRef]
- Carrascal, E.; Koriyama, C.; Akiba, S.; Tamayo, O.; Itoh, T.; Eizuru, Y.; Garcia, F.; Sera, M.; Carrasquilla, G.; Piazuelo, M.B.; et al. Epstein-Barr virus-associated gastric carcinoma in Cali, Colombia. Oncol. Rep. 2003, 10, 1059–1062. [Google Scholar]
- Yoshiwara, E.; Koriyama, C.; Akiba, S.; Itoh, T.; Minakami, Y.; Chirinos, J.L.; Watanabe, J.; Takano, J.; Miyagui, J.; Hidalgo, H.; et al. Epstein-Barr virus-associated gastric carcinoma in Lima, Peru. J. Exp. Clin. Cancer Res. 2005, 24, 49–54. [Google Scholar]
- Lopes, L.F.; Bacchi, M.M.; Elgui-de-Oliveira, D.; Zanati, S.G.; Alvarenga, M.; Bacchi, C.E. Epstein-Barr virus infection and gastric carcinoma in Sao Paulo State, Brazil. Braz. J. Med. Biol. Res. 2004, 37, 1707–1712. [Google Scholar] [CrossRef]
- Hayashi, K.; Chen, W.G.; Chen, Y.Y.; Murakami, I.; Chen, H.L.; Ohara, N.; Nose, S.; Hamaya, K.; Matsui, S.; Bacchi, M.M.; et al. Deletion of Epstein-Barr virus latent membrane protein 1 gene in Japanese and Brazilian gastric carcinomas, metastatic lesions, and reactive lymphocytes. Am. J. Pathol. 1998, 152, 191–198. [Google Scholar]
- Shousha, S.; Luqmani, Y.A. Epstein-Barr virus in gastric carcinoma and adjacent normal gastric and duodenal mucosa. J. Clin. Pathol. 1994, 47, 695–698. [Google Scholar] [CrossRef]
- Nishikawa, J.; Yanai, H.; Hirano, A.; Okamoto, T.; Nakamura, H.; Matsusaki, K.; Kawano, T.; Miura, O.; Okita, K. High prevalence of Epstein-Barr virus in gastric remnant carcinoma after Billroth-II reconstruction. Scand. J. Gastroenterol. 2002, 37, 825–829. [Google Scholar]
- Yamamoto, N.; Tokunaga, M.; Uemura, Y.; Tanaka, S.; Shirahama, H.; Nakamura, T.; Land, C.E.; Sato, E. Epstein-Barr virus and gastric remnant cancer. Cancer 1994, 74, 805–809. [Google Scholar] [CrossRef]
- Herath, C.H.; Chetty, R. Epstein-Barr virus-associated lymphoepithelioma-like gastric carcinoma. Arch. Pathol. Lab. Med. 2008, 132, 706–709. [Google Scholar]
- Shibata, D.; Tokunaga, M.; Uemura, Y.; Sato, E.; Tanaka, S.; Weiss, L.M. Association of Epstein-Barr virus with undifferentiated gastric carcinomas with intense lymphoid infiltration. Lymphoepithelioma-like carcinoma. Am. J. Pathol. 1991, 139, 469–474. [Google Scholar]
- Uemura, Y.; Tokunaga, M.; Arikawa, J.; Yamamoto, N.; Hamasaki, Y.; Tanaka, S.; Sato, E.; Land, C.E. A unique morphology of Epstein-Barr virus-related early gastric carcinoma. Cancer Epidemiol. Biomarkers Prev. 1994, 3, 607–611. [Google Scholar]
- Lino-Silva, L.S.; Salcedo Hernandez, R.A.; Molina-Frias, E. Mixed gastric carcinoma with intestinal and cribriform patterns: A distinctive pathologic appearance associated with poor prognosis in advanced stages and a potential mimicker of metastatic breast carcinoma. Int. J. Surg. Pathol. 2013, 21, 6–14. [Google Scholar] [CrossRef]
- Shibata, D.; Weiss, L.M. Epstein-Barr virus-associated gastric adenocarcinoma. Am. J. Pathol. 1992, 140, 769–774. [Google Scholar]
- Zur Hausen, A.; van Rees, B.P.; van Beek, J.; Craanen, M.E.; Bloemena, E.; Offerhaus, G.J.; Meijer, C.J.; van den Brule, A.J. Epstein-Barr virus in gastric carcinomas and gastric stump carcinomas: A late event in gastric carcinogenesis. J. Clin. Pathol. 2004, 57, 487–491. [Google Scholar] [CrossRef]
- Hungermann, D.; Muller, S.; Spieker, T.; Lisner, R.; Niedobitek, G.; Herbst, H. Low prevalence of latently Epstein-Barr virus-infected cells in chronic gastritis. Microsc. Res. Tech. 2001, 53, 409–413. [Google Scholar] [CrossRef]
- Yanai, H.; Takada, K.; Shimizu, N.; Mizugaki, Y.; Tada, M.; Okita, K. Epstein-Barr virus infection in non-carcinomatous gastric epithelium. J. Pathol. 1997, 183, 293–298. [Google Scholar] [CrossRef]
- Hirano, A.; Yanai, H.; Shimizu, N.; Okamoto, T.; Matsubara, Y.; Yamamoto, K.; Okita, K. Evaluation of epstein-barr virus DNA load in gastric mucosa with chronic atrophic gastritis using a real-time quantitative PCR assay. Int. J. Gastrointest Cancer 2003, 34, 87–94. [Google Scholar] [CrossRef]
- Chen, Z.M.; Shah, R.; Zuckerman, G.R.; Wang, H.L. Epstein-Barr virus gastritis: An underrecognized form of severe gastritis simulating gastric lymphoma. Am. J. Surg. Pathol. 2007, 31, 1446–1451. [Google Scholar] [CrossRef]
- Au, W.Y.; Pang, A.; Chan, E.C.; Chu, K.M.; Shek, T.W.; Kwong, Y.L. Epstein-barr virus-related gastric adenocarcinoma: An early secondary cancer post hemopoietic stem cell transplantation. Gastroenterology 2005, 129, 2058–2063. [Google Scholar] [CrossRef]
- Ohnishi, N.; Yuasa, H.; Tanaka, S.; Sawa, H.; Miura, M.; Matsui, A.; Higashi, H.; Musashi, M.; Iwabuchi, K.; Suzuki, M.; et al. Transgenic expression of Helicobacter pylori CagA induces gastrointestinal and hematopoietic neoplasms in mouse. Proc. Natl. Acad. Sci. USA 2008, 105, 1003–1008. [Google Scholar]
- Miura, M.; Ohnishi, N.; Tanaka, S.; Yanagiya, K.; Hatakeyama, M. Differential oncogenic potential of geographically distinct Helicobacter pylori CagA isoforms in mice. Int. J. Cancer 2009, 125, 2497–2504. [Google Scholar] [CrossRef]
- Castello, G.; Scala, S.; Palmieri, G.; Curley, S.A.; Izzo, F. HCV-related hepatocellular carcinoma: From chronic inflammation to cancer. Clin. Immunol. 2010, 134, 237–250. [Google Scholar] [CrossRef]
- Chemin, I.; Zoulim, F. Hepatitis B virus induced hepatocellular carcinoma. Cancer Lett. 2009, 286, 52–59. [Google Scholar] [CrossRef]
- Morales-Sanchez, A.; Molina-Munoz, T.; Martinez-Lopez, J.L.; Hernandez-Sancen, P.; Mantilla, A.; Leal, Y.A.; Torres, J.; Fuentes-Panana, E.M. No association between Epstein-Barr virus and mouse mammary tumor virus with breast cancer in Mexican women. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef]
- Labrecque, L.G.; Barnes, D.M.; Fentiman, I.S.; Griffin, B.E. Epstein-Barr virus in epithelial cell tumors: A breast cancer study. Cancer Res. 1995, 55, 39–45. [Google Scholar]
- Jones, M.D.; Griffin, B.E. Clustered repeat sequences in the genome of Epstein Barr virus. Nucleic Acids Res. 1983, 11, 3919–3937. [Google Scholar] [CrossRef]
- Basic Local Alignment Tool. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi/ (accessed on 15 July 2013).
- Luo, B.; Wang, Y.; Wang, X.F.; Liang, H.; Yan, L.P.; Huang, B.H.; Zhao, P. Expression of Epstein-Barr virus genes in EBV-associated gastric carcinomas. World J. Gastroenterol. 2005, 11, 629–633. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Martínez-López, J.L.E.; Torres, J.; Camorlinga-Ponce, M.; Mantilla, A.; Leal, Y.A.; Fuentes-Pananá, E.M. Evidence of Epstein-Barr Virus Association with Gastric Cancer and Non-Atrophic Gastritis. Viruses 2014, 6, 301-318. https://doi.org/10.3390/v6010301
Martínez-López JLE, Torres J, Camorlinga-Ponce M, Mantilla A, Leal YA, Fuentes-Pananá EM. Evidence of Epstein-Barr Virus Association with Gastric Cancer and Non-Atrophic Gastritis. Viruses. 2014; 6(1):301-318. https://doi.org/10.3390/v6010301
Chicago/Turabian StyleMartínez-López, Juan L.E., Javier Torres, Margarita Camorlinga-Ponce, Alejandra Mantilla, Yelda A. Leal, and Ezequiel M. Fuentes-Pananá. 2014. "Evidence of Epstein-Barr Virus Association with Gastric Cancer and Non-Atrophic Gastritis" Viruses 6, no. 1: 301-318. https://doi.org/10.3390/v6010301
APA StyleMartínez-López, J. L. E., Torres, J., Camorlinga-Ponce, M., Mantilla, A., Leal, Y. A., & Fuentes-Pananá, E. M. (2014). Evidence of Epstein-Barr Virus Association with Gastric Cancer and Non-Atrophic Gastritis. Viruses, 6(1), 301-318. https://doi.org/10.3390/v6010301