Host Restriction of Lentiviruses and Viral Countermeasures: APOBEC3 and Vif

{kind=link}
{kind=link}
Abstract
:1. Introduction
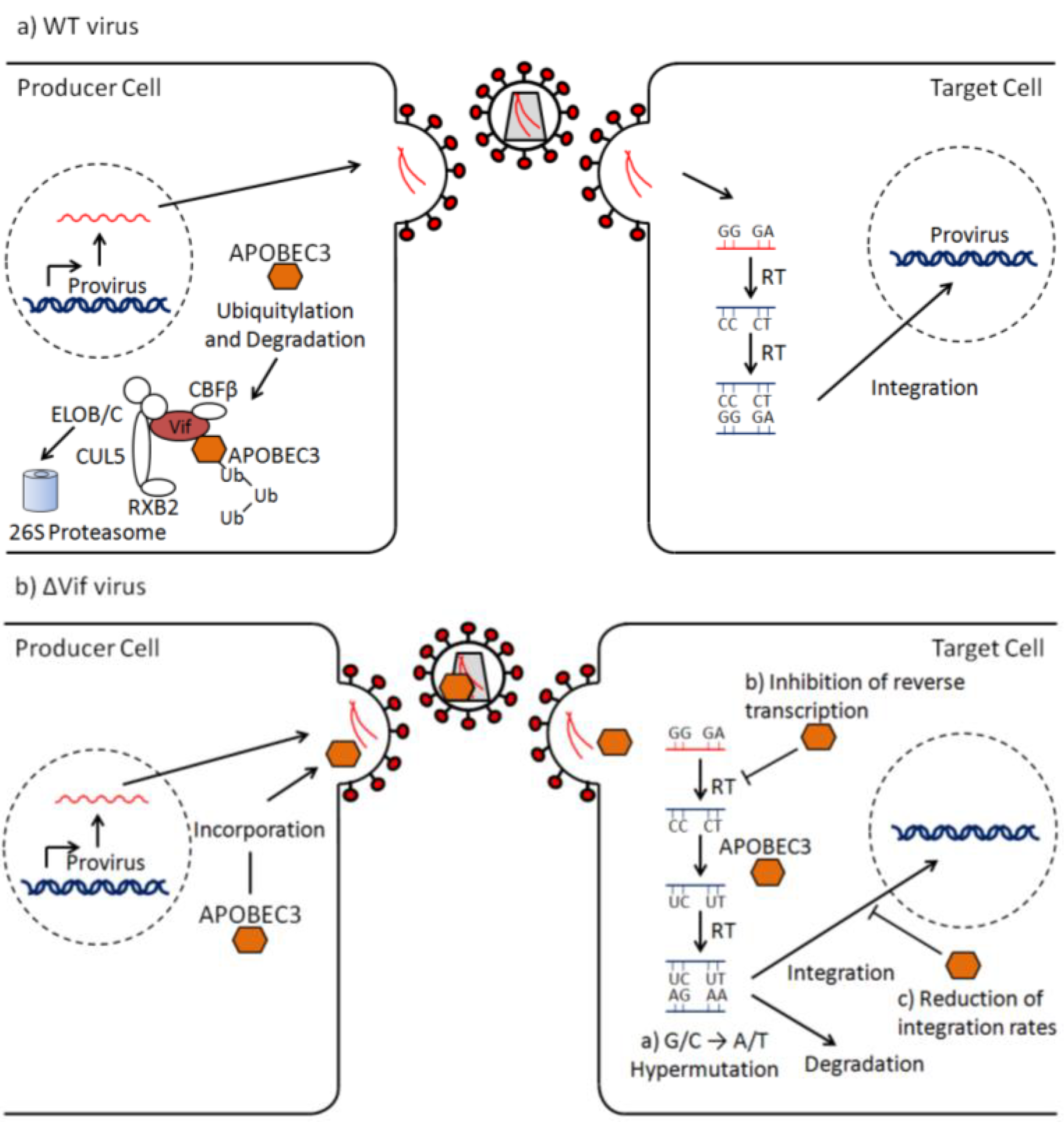
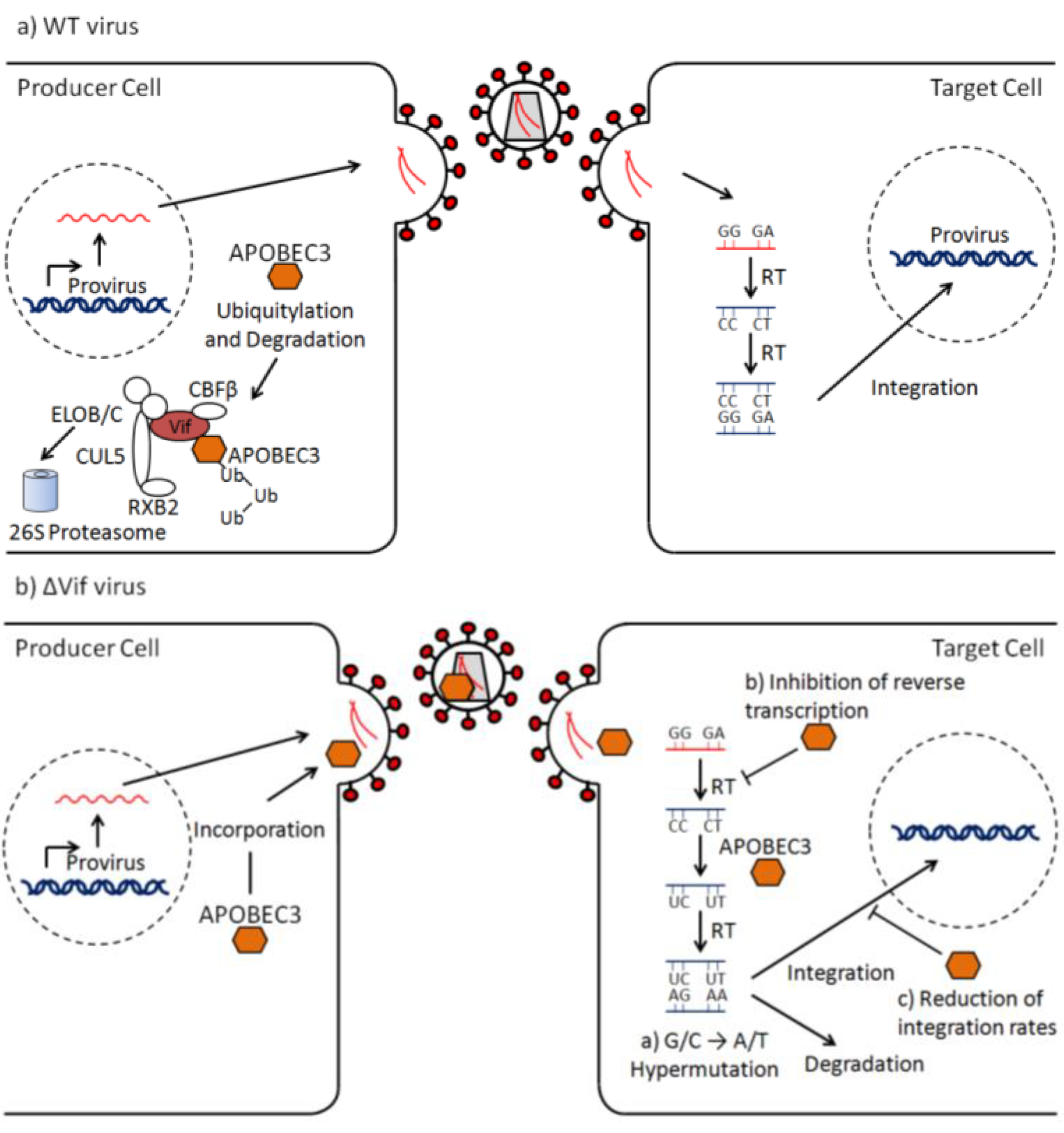
2. The ΔVif Phenotype
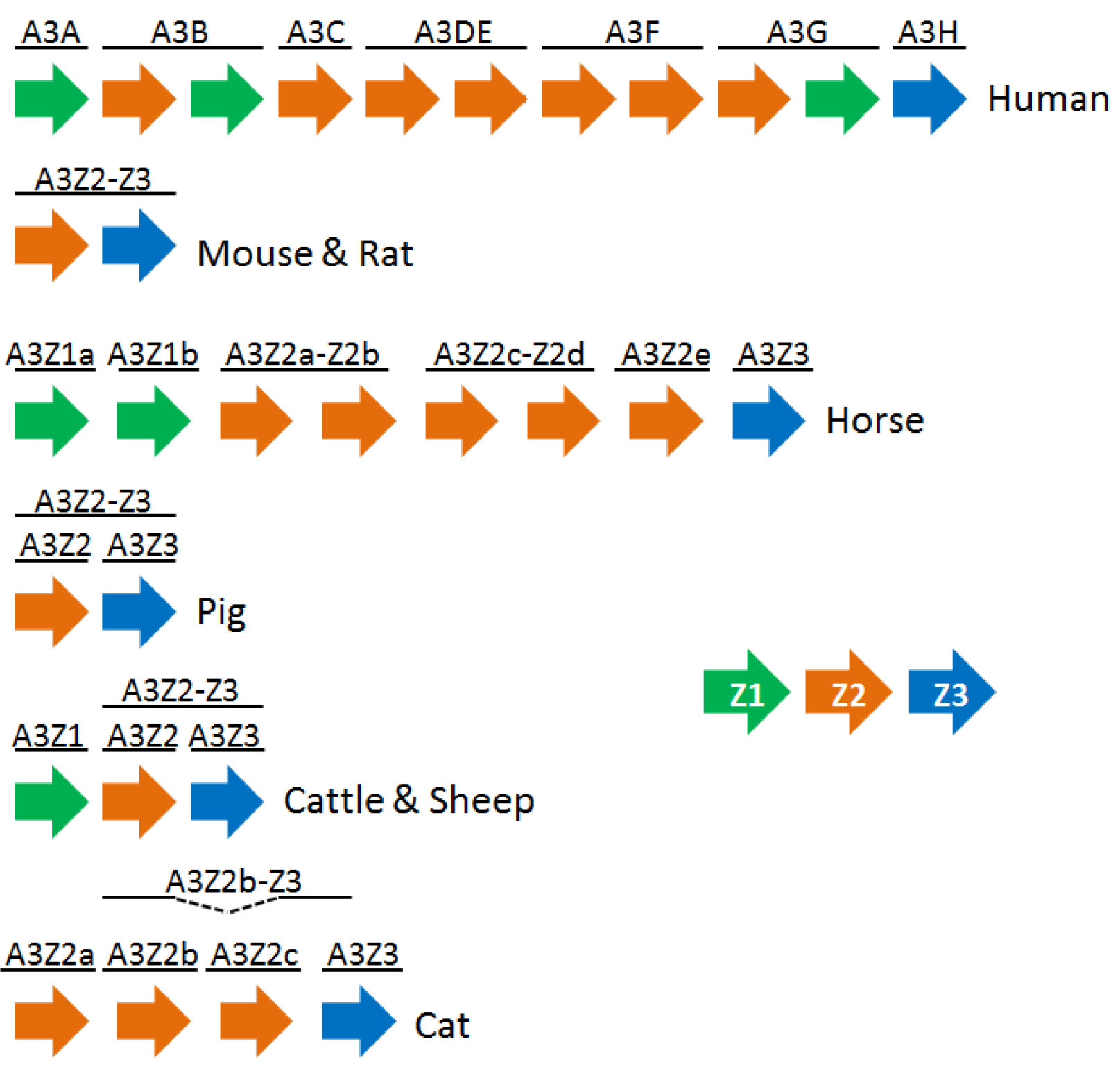
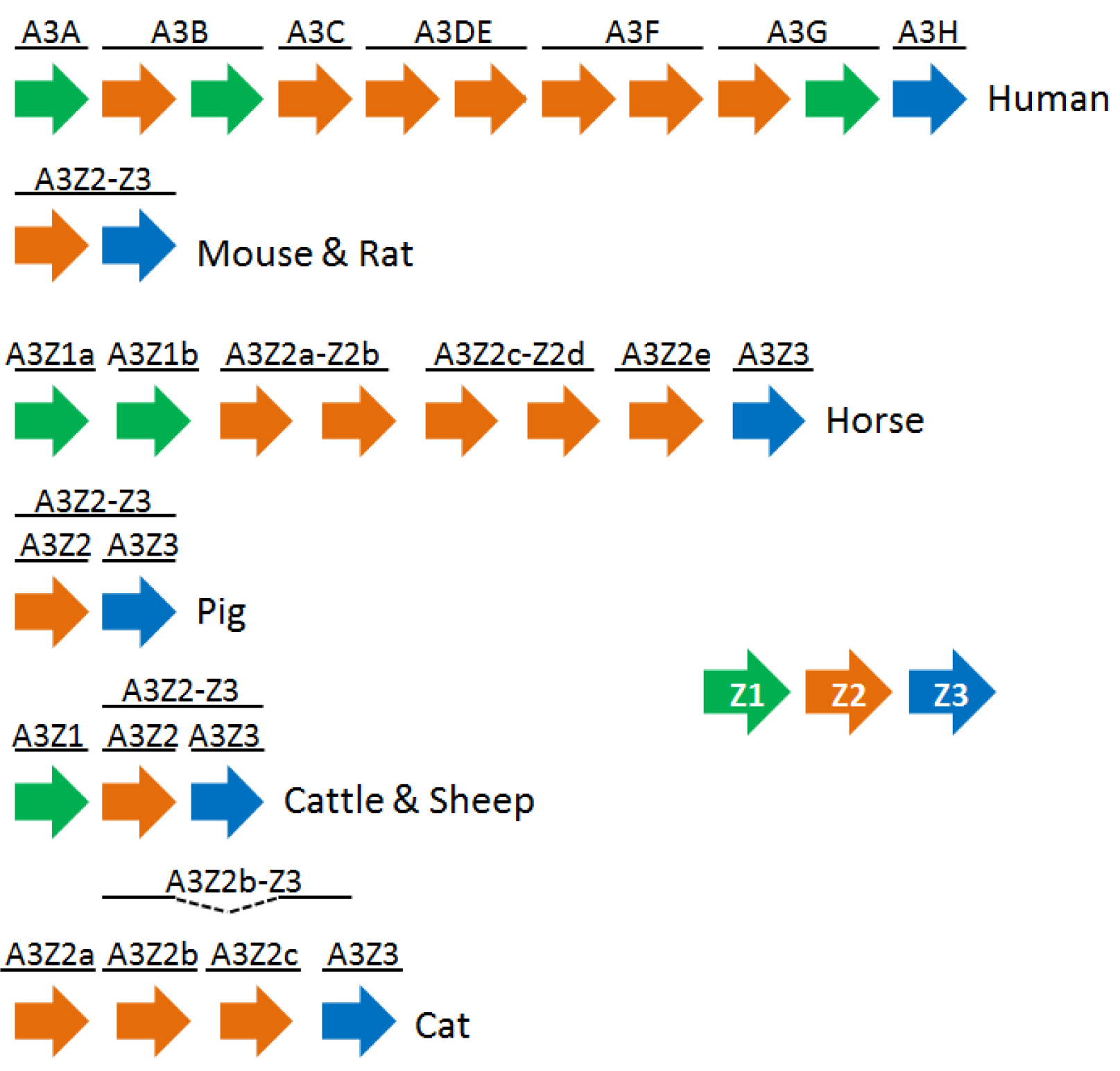
3. The APOBEC3 Family of Cytidine Deaminases
4. Inhibition of Viral Growth
5. Modes of Viral Escape
6. Ovine A3 Proteins and MVV Vif
7. Concluding Remarks
Acknowledgments
Conflict of Interest
References and Notes
- Malim, M.H.; Emerman, M. HIV-1 accessory proteins—Ensuring viral survival in a hostile environment. Cell Host Microbe 2008, 3, 388–398. [Google Scholar] [CrossRef]
- Laguette, N.; Sobhian, B.; Casartelli, N.; Ringeard, M.; Chable-Bessia, C.; Segeral, E.; Yatim, A.; Emiliani, S.; Schwartz, O.; Benkirane, M. SAMHD1 is the dendritic- and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature 2011, 474, 654–657. [Google Scholar] [CrossRef]
- Neil, S.J.; Zang, T.; Bieniasz, P.D. Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature 2008, 451, 425–430. [Google Scholar] [CrossRef]
- Harris, R.S.; Hultquist, J.F.; Evans, D.T. The restriction factors of human immunodeficiency virus. J. Biol. Chem. 2012, 287, 40875–40883. [Google Scholar]
- Audoly, G.; Sauze, N.; Harkiss, G.; Vitu, C.; Russo, P.; Querat, G.; Suzan, M.; Vigne, R. Identification and subcellular localization of the Q gene product of visna virus. Virology 1992, 189, 734–739. [Google Scholar] [CrossRef]
- Lee, T.H.; Coligan, J.E.; Allan, J.S.; McLane, M.F.; Groopman, J.E.; Essex, M. A new HTLV-III/LAV protein encoded by a gene found in cytopathic retroviruses. Science 1986, 231, 1546–1549. [Google Scholar]
- Oberste, M.S.; Gonda, M.A. Conservation of amino-acid sequence motifs in lentivirus Vif proteins. Virus Genes 1992, 6, 95–102. [Google Scholar] [CrossRef]
- Gabuzda, D.H.; Lawrence, K.; Langhoff, E.; Terwilliger, E.; Dorfman, T.; Haseltine, W.A.; Sodroski, J. Role of vif in replication of human immunodeficiency virus type 1 in CD4+ T lymphocytes. J. Virol. 1992, 66, 6489–6495. [Google Scholar]
- Strebel, K.; Daugherty, D.; Clouse, K.; Cohen, D.; Folks, T.; Martin, M.A. The HIV 'A' (sor) gene product is essential for virus infectivity. Nature 1987, 328, 728–730. [Google Scholar] [CrossRef]
- Fan, L.; Peden, K. Cell-free transmission of Vif mutants of HIV-1. Virology 1992, 190, 19–29. [Google Scholar] [CrossRef]
- Reddy, T.R.; Kraus, G.; Suhasini, M.; Leavitt, M.C.; Wong-Staal, F. Identification and mapping of inhibitory sequences in the human immunodeficiency virus type 2 vif gene. J. Virol. 1995, 69, 5167–5170. [Google Scholar]
- Desrosiers, R.C.; Lifson, J.D.; Gibbs, J.S.; Czajak, S.C.; Howe, A.Y.; Arthur, L.O.; Johnson, R.P. Identification of highly attenuated mutants of simian immunodeficiency virus. J. Virol. 1998, 72, 1431–1437. [Google Scholar]
- Lockridge, K.M.; Himathongkham, S.; Sawai, E.T.; Chienand, M.; Sparger, E.E. The feline immunodeficiency virus vif gene is required for productive infection of feline peripheral blood mononuclear cells and monocyte-derived macrophages. Virology 1999, 261, 25–30. [Google Scholar] [CrossRef]
- Tomonaga, K.; Norimine, J.; Shin, Y.S.; Fukasawa, M.; Miyazawa, T.; Adachi, A.; Toyosaki, T.; Kawaguchi, Y.; Kai, C.; Mikami, T. Identification of a feline immunodeficiency virus gene which is essential for cell-free virus infectivity. J. Virol. 1992, 66, 6181–6185. [Google Scholar]
- Harmache, A.; Bouyac, M.; Audoly, G.; Hieblot, C.; Peveri, P.; Vigne, R.; Suzan, M. The vif gene is essential for efficient replication of caprine arthritis encephalitis virus in goat synovial membrane cells and affects the late steps of the virus replication cycle. J. Virol. 1995, 69, 3247–3257. [Google Scholar]
- Harmache, A.; Russo, P.; Guiguen, F.; Vitu, C.; Vignoni, M.; Bouyac, M.; Hieblot, C.; Pepin, M.; Vigne, R.; Suzan, M. Requirement of caprine arthritis encephalitis virus vif gene for in vivo replication. Virology 1996, 224, 246–255. [Google Scholar] [CrossRef]
- Kristbjornsdottir, H.B.; Andresdottir, V.; Svansson, V.; Torsteinsdottir, S.; Matthiasdottir, S.; Andresson, O.S. The vif gene of maedi-visna virus is essential for infectivity in vivo and in vitro. Virology 2004, 318, 350–359. [Google Scholar] [CrossRef]
- Bouyac, M.; Rey, F.; Nascimbeni, M.; Courcoul, M.; Sire, J.; Blanc, D.; Clavel, F.; Vigne, R.; Spire, B. Phenotypically Vif- human immunodeficiency virus type 1 is produced by chronically infected restrictive cells. J. Virol. 1997, 71, 2473–2477. [Google Scholar]
- Chowdhury, I.H.; Chao, W.; Potash, M.J.; Sova, P.; Gendelman, H.E.; Volsky, D.J. Vif-negative human immunodeficiency virus type 1 persistently replicates in primary macrophages, producing attenuated progeny virus. J. Virol. 1996, 70, 5336–5345. [Google Scholar]
- Courcoul, M.; Patience, C.; Rey, F.; Blanc, D.; Harmache, A.; Sire, J.; Vigne, R.; Spire, B. Peripheral blood mononuclear cells produce normal amounts of defective Vif- human immunodeficiency virus type 1 particles which are restricted for the preretrotranscription steps. J. Virol. 1995, 69, 2068–2074. [Google Scholar]
- Ochsenbauer, C.; Wilk, T.; Bosch, V. Analysis of vif-defective human immunodeficiency virus type 1 (HIV-1) virions synthesized in 'non-permissive' T lymphoid cells stably infected with selectable HIV-1. J. Gen. Virol. 1997, 78, 627–635. [Google Scholar]
- Von Schwedler, U.; Song, J.; Aiken, C.; Trono, D. Vif is crucial for human immunodeficiency virus type 1 proviral DNA synthesis in infected cells. J. Virol. 1993, 67, 4945–4955. [Google Scholar]
- Madani, N.; Kabat, D. An endogenous inhibitor of human immunodeficiency virus in human lymphocytes is overcome by the viral Vif protein. J. Virol. 1998, 72, 10251–10255. [Google Scholar]
- Simon, J.H.; Gaddis, N.C.; Fouchier, R.A.; Malim, M.H. Evidence for a newly discovered cellular anti-HIV-1 phenotype. Nat. Med. 1998, 4, 1397–1400. [Google Scholar] [CrossRef]
- Sheehy, A.M.; Gaddis, N.C.; Choi, J.D.; Malim, M.H. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature 2002, 418, 646–650. [Google Scholar] [CrossRef]
- Jarmuz, A.; Chester, A.; Bayliss, J.; Gisbourne, J.; Dunham, I.; Scott, J.; Navaratnam, N. An anthropoid-specific locus of orphan C to U RNA-editing enzymes on chromosome 22. Genomics 2002, 79, 285–296. [Google Scholar] [CrossRef]
- Conticello, S.G.; Thomas, C.J.; Petersen-Mahrt, S.K.; Neuberger, M.S. Evolution of the AID/APOBEC family of polynucleotide (deoxy)cytidine deaminases. Mol. Biol. Evol. 2005, 22, 367–377. [Google Scholar]
- Rogozin, I.B.; Basu, M.K.; Jordan, I.K.; Pavlov, Y.I.; Koonin, E.V. APOBEC4, a new member of the AID/APOBEC family of polynucleotide (deoxy)cytidine deaminases predicted by computational analysis. Cell Cycle 2005, 4, 1281–1285. [Google Scholar] [CrossRef]
- Teng, B.; Burant, C.F.; Davidson, N.O. Molecular cloning of an apolipoprotein B messenger RNA editing protein. Science 1993, 260, 1816–1819. [Google Scholar]
- Yamanaka, S.; Balestra, M.E.; Ferrell, L.D.; Fan, J.; Arnold, K.S.; Taylor, S.; Taylor, J.M.; Innerarity, T.L. Apolipoprotein B mRNA-editing protein induces hepatocellular carcinoma and dysplasia in transgenic animals. Proc. Natl. Acad. Sci. USA 1995, 92, 8483–8487. [Google Scholar] [CrossRef]
- Harris, R.S.; Bishop, K.N.; Sheehy, A.M.; Craig, H.M.; Petersen-Mahrt, S.K.; Watt, I.N.; Neuberger, M.S.; Malim, M.H. DNA deamination mediates innate immunity to retroviral infection. Cell 2003, 113, 803–809. [Google Scholar] [CrossRef]
- Mangeat, B.; Turelli, P.; Caron, G.; Friedli, M.; Perrin, L.; Trono, D. Broad antiretroviral defence by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature 2003, 424, 99–103. [Google Scholar] [CrossRef]
- Mariani, R.; Chen, D.; Schrofelbauer, B.; Navarro, F.; Konig, R.; Bollman, B.; Munk, C.; Nymark-McMahon, H.; Landau, N.R. Species-specific exclusion of APOBEC3G from HIV-1 virions by Vif. Cell 2003, 114, 21–31. [Google Scholar] [CrossRef]
- Wedekind, J.E.; Dance, G.S.; Sowden, M.P.; Smith, H.C. Messenger RNA editing in mammals: New members of the APOBEC family seeking roles in the family business. Trends Genet. 2003, 19, 207–216. [Google Scholar] [CrossRef]
- LaRue, R.S.; Andresdottir, V.; Blanchard, Y.; Conticello, S.G.; Derse, D.; Emerman, M.; Greene, W.C.; Jonsson, S.R.; Landau, N.R.; Lochelt, M.; et al. Guidelines for naming nonprimate APOBEC3 genes and proteins. J. Virol. 2009, 83, 494–497. [Google Scholar] [CrossRef]
- LaRue, R.S.; Jonsson, S.R.; Silverstein, K.A.; Lajoie, M.; Bertrand, D.; El-Mabrouk, N.; Hotzel, I.; Andresdottir, V.; Smith, T.P.; Harris, R.S. The artiodactyl APOBEC3 innate immune repertoire shows evidence for a multi-functional domain organization that existed in the ancestor of placental mammals. BMC Mol. Biol. 2008, 9, 104. [Google Scholar] [CrossRef]
- Doehle, B.P.; Schafer, A.; Cullen, B.R. Human APOBEC3B is a potent inhibitor of HIV-1 infectivity and is resistant to HIV-1 Vif. Virology 2005, 339, 281–288. [Google Scholar] [CrossRef]
- Bishop, K.N.; Holmes, R.K.; Sheehy, A.M.; Malim, M.H. APOBEC-mediated editing of viral RNA. Science 2004, 305, 645. [Google Scholar] [CrossRef]
- Liddament, M.T.; Brown, W.L.; Schumacher, A.J.; Harris, R.S. APOBEC3F properties and hypermutation preferences indicate activity against HIV-1 in vivo. Curr. Biol. 2004, 14, 1385–1391. [Google Scholar] [CrossRef]
- Albin, J.S.; Harris, R.S. Interactions of host APOBEC3 restriction factors with HIV-1 in vivo: Implications for therapeutics. Expert Rev. Mol. Med. 2010, 12, e4. [Google Scholar] [CrossRef]
- Esnault, C.; Heidmann, O.; Delebecque, F.; Dewannieux, M.; Ribet, D.; Hance, A.J.; Heidmann, T.; Schwartz, O. APOBEC3G cytidine deaminase inhibits retrotransposition of endogenous retroviruses. Nature 2005, 433, 430–433. [Google Scholar] [CrossRef]
- Jonsson, S.R.; Hache, G.; Stenglein, M.D.; Fahrenkrug, S.C.; Andresdottir, V.; Harris, R.S. Evolutionarily conserved and non-conserved retrovirus restriction activities of artiodactyl APOBEC3F proteins. Nucleic Acids Res. 2006, 34, 5683–5694. [Google Scholar] [CrossRef]
- Dutko, J.A.; Schafer, A.; Kenny, A.E.; Cullen, B.R.; Curcio, M.J. Inhibition of a yeast LTR retrotransposon by human APOBEC3 cytidine deaminases. Curr. Biol. 2005, 15, 661–666. [Google Scholar] [CrossRef]
- Schumacher, A.J.; Nissley, D.V.; Harris, R.S. APOBEC3G hypermutates genomic DNA and inhibits Ty1 retrotransposition in yeast. Proc. Natl. Acad. Sci. USA 2005, 102, 9854–9859. [Google Scholar] [CrossRef]
- Bogerd, H.P.; Cullen, B.R. Single-stranded RNA facilitates nucleocapsid: APOBEC3G complex formation. RNA 2008, 14, 1228–1236. [Google Scholar] [CrossRef]
- Khan, M.A.; Kao, S.; Miyagi, E.; Takeuchi, H.; Goila-Gaur, R.; Opi, S.; Gipson, C.L.; Parslow, T.G.; Ly, H.; Strebel, K. Viral RNA is required for the association of APOBEC3G with human immunodeficiency virus type 1 nucleoprotein complexes. J. Virol. 2005, 79, 5870–5874. [Google Scholar] [CrossRef]
- Luo, K.; Liu, B.; Xiao, Z.; Yu, Y.; Yu, X.; Gorelick, R.; Yu, X.F. Amino-terminal region of the human immunodeficiency virus type 1 nucleocapsid is required for human APOBEC3G packaging. J. Virol. 2004, 78, 11841–11852. [Google Scholar] [CrossRef]
- Schafer, A.; Bogerd, H.P.; Cullen, B.R. Specific packaging of APOBEC3G into HIV-1 virions is mediated by the nucleocapsid domain of the gag polyprotein precursor. Virology 2004, 328, 163–168. [Google Scholar] [CrossRef]
- Svarovskaia, E.S.; Xu, H.; Mbisa, J.L.; Barr, R.; Gorelick, R.J.; Ono, A.; Freed, E.O.; Hu, W.S.; Pathak, V.K. Human apolipoprotein B mRNA-editing enzyme-catalytic polypeptide-like 3G (APOBEC3G) is incorporated into HIV-1 virions through interactions with viral and nonviral RNAs. J. Biol. Chem. 2004, 279, 35822–35828. [Google Scholar] [CrossRef]
- Zennou, V.; Perez-Caballero, D.; Gottlinger, H.; Bieniasz, P.D. APOBEC3G incorporation into human immunodeficiency virus type 1 particles. J. Virol. 2004, 78, 12058–12061. [Google Scholar] [CrossRef]
- Newman, E.N.; Holmes, R.K.; Craig, H.M.; Klein, K.C.; Lingappa, J.R.; Malim, M.H.; Sheehy, A.M. Antiviral function of APOBEC3G can be dissociated from cytidine deaminase activity. Curr. Biol. 2005, 15, 166–170. [Google Scholar] [CrossRef]
- Holmes, R.K.; Koning, F.A.; Bishop, K.N.; Malim, M.H. APOBEC3F can inhibit the accumulation of HIV-1 reverse transcription products in the absence of hypermutation. Comparisons with APOBEC3G. J. Biol. Chem. 2007, 282, 2587–2595. [Google Scholar]
- Bishop, K.N.; Verma, M.; Kim, E.Y.; Wolinsky, S.M.; Malim, M.H. APOBEC3G inhibits elongation of HIV-1 reverse transcripts. PLoS Pathog. 2008, 4, e1000231. [Google Scholar] [CrossRef]
- Iwatani, Y.; Chan, D.S.; Wang, F.; Maynard, K.S.; Sugiura, W.; Gronenborn, A.M.; Rouzina, I.; Williams, M.C.; Musier-Forsyth, K.; Levin, J.G. Deaminase-independent inhibition of HIV-1 reverse transcription by APOBEC3G. Nucleic Acids Res. 2007, 35, 7096–7108. [Google Scholar] [CrossRef]
- Guo, F.; Cen, S.; Niu, M.; Saadatmand, J.; Kleiman, L. Inhibition of formula-primed reverse transcription by human APOBEC3G during human immunodeficiency virus type 1 replication. J. Virol. 2006, 80, 11710–11722. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, F.; Cen, S.; Kleiman, L. Inhibition of initiation of reverse transcription in HIV-1 by human APOBEC3F. Virology 2007, 365, 92–100. [Google Scholar] [CrossRef]
- Li, X.Y.; Guo, F.; Zhang, L.; Kleiman, L.; Cen, S. APOBEC3G inhibits DNA strand transfer during HIV-1 reverse transcription. J. Biol. Chem. 2007, 282, 32065–32074. [Google Scholar] [CrossRef]
- Mbisa, J.L.; Barr, R.; Thomas, J.A.; Vandegraaff, N.; Dorweiler, I.J.; Svarovskaia, E.S.; Brown, W.L.; Mansky, L.M.; Gorelick, R.J.; Harris, R.S.; et al. Human immunodeficiency virus type 1 cDNAs produced in the presence of APOBEC3G exhibit defects in plus-strand DNA transfer and integration. J. Virol. 2007, 81, 7099–7110. [Google Scholar] [CrossRef]
- Okeoma, C.M.; Lovsin, N.; Peterlin, B.M.; Ross, S.R. APOBEC3 inhibits mouse mammary tumour virus replication in vivo. Nature 2007, 445, 927–930. [Google Scholar] [CrossRef]
- Sanchez-Martinez, S.; Aloia, A.L.; Harvin, D.; Mirro, J.; Gorelick, R.J.; Jern, P.; Coffin, J.M.; Rein, A. Studies on the restriction of murine leukemia viruses by mouse APOBEC3. PLoS One 2012, 7, e38190. [Google Scholar] [CrossRef]
- Refsland, E.W.; Hultquist, J.F.; Harris, R.S. Endogenous origins of HIV-1 G-to-A hypermutation and restriction in the nonpermissive T cell line CEM2n. PLoS Pathog. 2012, 8, e1002800. [Google Scholar] [CrossRef]
- Chaipan, C.; Smith, J.L.; Hu, W.S.; Pathak, V.K. APOBEC3G restricts HIV-1 to a greater extent than APOBEC3F and APOBEC3DE in human primary CD4+ T cells and macrophages. J. Virol. 2013, 87, 444–453. [Google Scholar] [CrossRef]
- Zhen, A.; Du, J.; Zhou, X.; Xiong, Y.; Yu, X.F. Reduced APOBEC3H variant anti-viral activities are associated with altered RNA binding activities. PLoS One 2012, 7, e38771. [Google Scholar]
- OhAinle, M.; Kerns, J.A.; Malik, H.S.; Emerman, M. Adaptive evolution and antiviral activity of the conserved mammalian cytidine deaminase APOBEC3H. J. Virol. 2006, 80, 3853–3862. [Google Scholar] [CrossRef]
- Hultquist, J.F.; Lengyel, J.A.; Refsland, E.W.; LaRue, R.S.; Lackey, L.; Brown, W.L.; Harris, R.S. Human and rhesus APOBEC3D, APOBEC3F, APOBEC3G, and APOBEC3H demonstrate a conserved capacity to restrict Vif-deficient HIV-1. J. Virol. 2011, 85, 11220–11234. [Google Scholar] [CrossRef]
- Koning, F.A.; Newman, E.N.; Kim, E.Y.; Kunstman, K.J.; Wolinsky, S.M.; Malim, M.H. Defining APOBEC3 expression patterns in human tissues and hematopoietic cell subsets. J. Virol. 2009, 83, 9474–9485. [Google Scholar]
- Refsland, E.W.; Stenglein, M.D.; Shindo, K.; Albin, J.S.; Brown, W.L.; Harris, R.S. Quantitative profiling of the full APOBEC3 mRNA repertoire in lymphocytes and tissues: Implications for HIV-1 restriction. Nucleic Acids Res. 2010, 38, 4274–4284. [Google Scholar] [CrossRef]
- Stenglein, M.D.; Burns, M.B.; Li, M.; Lengyel, J.; Harris, R.S. APOBEC3 proteins mediate the clearance of foreign DNA from human cells. Nat. Struct. Mol. Biol. 2010, 17, 222–229. [Google Scholar] [CrossRef]
- Berger, G.; Durand, S.; Fargier, G.; Nguyen, X.N.; Cordeil, S.; Bouaziz, S.; Muriaux, D.; Darlix, J.L.; Cimarelli, A. APOBEC3A is a specific inhibitor of the early phases of HIV-1 infection in myeloid cells. PLoS Pathog. 2011, 7, e1002221. [Google Scholar] [CrossRef]
- Koning, F.A.; Goujon, C.; Bauby, H.; Malim, M.H. Target cell-mediated editing of HIV-1 cDNA by APOBEC3 proteins in human macrophages. J. Virol. 2011, 85, 13448–13452. [Google Scholar] [CrossRef]
- Landry, S.; Narvaiza, I.; Linfesty, D.C.; Weitzman, M.D. APOBEC3A can activate the DNA damage response and cause cell-cycle arrest. EMBO Rep. 2011, 12, 444–450. [Google Scholar] [CrossRef]
- Jamieson, A.M.; Diefenbach, A.; McMahon, C.W.; Xiong, N.; Carlyle, J.R.; Raulet, D.H. The role of the NKG2D immunoreceptor in immune cell activation and natural killing. Immunity 2002, 17, 19–29. [Google Scholar] [CrossRef]
- Norman, J.M.; Mashiba, M.; McNamara, L.A.; Onafuwa-Nuga, A.; Chiari-Fort, E.; Shen, W.; Collins, K.L. The antiviral factor APOBEC3G enhances the recognition of HIV-infected primary T cells by natural killer cells. Nat. Immunol. 2011, 12, 975–983. [Google Scholar] [CrossRef]
- Sheehy, A.M.; Erthal, J. APOBEC3 versus retroviruses, immunity versus invasion: Clash of the titans. Mol. Biol. Int. 2012, 974924. [Google Scholar]
- Harris, R.S. Enhancing immunity to HIV through APOBEC. Nat. Biotechnol. 2008, 26, 1089–1090. [Google Scholar] [CrossRef]
- Simon, V.; Zennou, V.; Murray, D.; Huang, Y.; Ho, D.D.; Bieniasz, P.D. Natural variation in Vif: Differential impact on APOBEC3G/3F and a potential role in HIV-1 diversification. PLoS Pathog. 2005, 1, e6. [Google Scholar] [CrossRef]
- Kim, E.Y.; Bhattacharya, T.; Kunstman, K.; Swantek, P.; Koning, F.A.; Malim, M.H.; Wolinsky, S.M. Human APOBEC3G-mediated editing can promote HIV-1 sequence diversification and accelerate adaptation to selective pressure. J. Virol. 2010, 84, 10402–10405. [Google Scholar] [CrossRef]
- Jern, P.; Russell, R.A.; Pathak, V.K.; Coffin, J.M. Likely role of APOBEC3G-mediated G-to-A mutations in HIV-1 evolution and drug resistance. PLoS Pathog. 2009, 5, e1000367. [Google Scholar] [CrossRef]
- Hultquist, J.F.; Harris, R.S. Leveraging APOBEC3 proteins to alter the HIV mutation rate and combat AIDS. Future Virol. 2009, 4, 605. [Google Scholar] [CrossRef]
- Berkhout, B.; de Ronde, A. APOBEC3G versus reverse transcriptase in the generation of HIV-1 drug-resistance mutations. AIDS 2004, 18, 1861–1863. [Google Scholar] [CrossRef]
- Yan, N.; O'Day, E.; Wheeler, L.A.; Engelman, A.; Lieberman, J. HIV DNA is heavily uracilated, which protects it from autointegration. Proc. Natl. Acad. Sci. USA 2011, 108, 9244–9249. [Google Scholar] [CrossRef]
- Conticello, S.G.; Harris, R.S.; Neuberger, M.S. The Vif protein of HIV triggers degradation of the human antiretroviral DNA deaminase APOBEC3G. Curr. Biol. 2003, 13, 2009–2013. [Google Scholar] [CrossRef]
- Sheehy, A.M.; Gaddis, N.C.; Malim, M.H. The antiretroviral enzyme APOBEC3G is degraded by the proteasome in response to HIV-1 Vif. Nat. Med. 2003, 9, 1404–1407. [Google Scholar] [CrossRef]
- Yu, X.; Yu, Y.; Liu, B.; Luo, K.; Kong, W.; Mao, P.; Yu, X.F. Induction of APOBEC3G ubiquitination and degradation by an HIV-1 Vif-Cul5-SCF complex. Science 2003, 302, 1056–1060. [Google Scholar] [CrossRef]
- Yu, Y.; Xiao, Z.; Ehrlich, E.S.; Yu, X.; Yu, X.F. Selective assembly of HIV-1 Vif-Cul5-ElonginB-ElonginC E3 ubiquitin ligase complex through a novel SOCS box and upstream cysteines. Genes Dev. 2004, 18, 2867–2872. [Google Scholar] [CrossRef]
- Stanley, B.J.; Ehrlich, E.S.; Short, L.; Yu, Y.; Xiao, Z.; Yu, X.F.; Xiong, Y. Structural insight into the human immunodeficiency virus Vif SOCS box and its role in human E3 ubiquitin ligase assembly. J. Virol. 2008, 82, 8656–8663. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, G.; Niewiadomska, A.M.; Xu, R.; Yu, X.F. Distinct determinants in HIV-1 Vif and human APOBEC3 proteins are required for the suppression of diverse host anti-viral proteins. PLoS One 2008, 3, e3963. [Google Scholar]
- Jager, S.; Kim, D.Y.; Hultquist, J.F.; Shindo, K.; LaRue, R.S.; Kwon, E.; Li, M.; Anderson, B.D.; Yen, L.; Stanley, D.; et al. Vif hijacks CBF-beta to degrade APOBEC3G and promote HIV-1 infection. Nature 2012, 481, 371–375. [Google Scholar]
- Zhang, W.; Du, J.; Evans, S.L.; Yu, Y.; Yu, X.F. T-cell differentiation factor CBF-beta regulates HIV-1 Vif-mediated evasion of host restriction. Nature 2012, 481, 376–379. [Google Scholar]
- Hultquist, J.F.; Binka, M.; LaRue, R.S.; Simon, V.; Harris, R.S. Vif proteins of human and simian immunodeficiency viruses require cellular CBFbeta to degrade APOBEC3 restriction factors. J. Virol. 2012, 86, 2874–2877. [Google Scholar] [CrossRef]
- Li, M.M.; Emerman, M. Polymorphism in human APOBEC3H affects a phenotype dominant for subcellular localization and antiviral activity. J. Virol. 2011, 85, 8197–8207. [Google Scholar] [CrossRef]
- Larue, R.S.; Lengyel, J.; Jonsson, S.R.; Andresdottir, V.; Harris, R.S. Lentiviral Vif degrades the APOBEC3Z3/APOBEC3H protein of its mammalian host and is capable of cross-species activity. J. Virol. 2010, 84, 8193–8201. [Google Scholar] [CrossRef]
- Franzdóttir, S.R.; Ólafsdóttir, K.; Jonsson, S.R.; Andresdottir, V.; University of Iceland, Reykjavik, Iceland. To be submitted. 2013.
- Mahrour, N.; Redwine, W.B.; Florens, L.; Swanson, S.K.; Martin-Brown, S.; Bradford, W.D.; Staehling-Hampton, K.; Washburn, M.P.; Conaway, R.C.; Conaway, J.W. Characterization of Cullin-box sequences that direct recruitment of Cul2-Rbx1 and Cul5-Rbx2 modules to elongin BC-based ubiquitin ligases. J. Biol. Chem. 2008, 283, 8005–8013. [Google Scholar] [CrossRef]
- Luo, K.; Xiao, Z.; Ehrlich, E.; Yu, Y.; Liu, B.; Zheng, S.; Yu, X.F. Primate lentiviral virion infectivity factors are substrate receptors that assemble with cullin 5-E3 ligase through a HCCH motif to suppress APOBEC3G. Proc. Natl. Acad. Sci. USA 2005, 102, 11444–11449. [Google Scholar] [CrossRef]
- Björnsdóttir, H.L.; Andrésdóttir, V.; University of Iceland, Reykjavik, Iceland. To be submitted. 2013.
- Wang, J.; Zhang, W.; Lv, M.; Zuo, T.; Kong, W.; Yu, X. Identification of a Cullin5-ElonginB-ElonginC E3 complex in degradation of feline immunodeficiency virus Vif-mediated feline APOBEC3 proteins. J. Virol. 2011, 85, 12482–12491. [Google Scholar] [CrossRef]
- Gudmundsson, B.; Jonsson, S.R.; Olafsson, O.; Agnarsdottir, G.; Matthiasdottir, S.; Georgsson, G.; Torsteinsdottir, S.; Svansson, V.; Kristbjornsdottir, H.B.; Franzdottir, S.R.; et al. Simultaneous mutations in CA and Vif of maedi-visna virus cause attenuated replication in macrophages and reduced infectivity in vivo. J. Virol. 2005, 79, 15038–15042. [Google Scholar] [CrossRef]
- Borman, A.M.; Quillent, C.; Charneau, P.; Dauguet, C.; Clavel, F. Human immunodeficiency virus type 1 Vif- mutant particles from restrictive cells: Role of Vif in correct particle assembly and infectivity. J. Virol. 1995, 69, 2058–2067. [Google Scholar]
- Hoglund, S.; Ohagen, A.; Lawrence, K.; Gabuzda, D. Role of Vif during packing of the core of HIV-1. Virology 1994, 201, 349–355. [Google Scholar] [CrossRef]
- Ohagen, A.; Gabuzda, D. Role of Vif in stability of the human immunodeficiency virus type 1 core. J. Virol. 2000, 74, 11055–11066. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jónsson, S.R.; Andrésdóttir, V. Host Restriction of Lentiviruses and Viral Countermeasures: APOBEC3 and Vif. Viruses 2013, 5, 1934-1947. https://doi.org/10.3390/v5081934
Jónsson SR, Andrésdóttir V. Host Restriction of Lentiviruses and Viral Countermeasures: APOBEC3 and Vif. Viruses. 2013; 5(8):1934-1947. https://doi.org/10.3390/v5081934
Chicago/Turabian StyleJónsson, Stefán R., and Valgerdur Andrésdóttir. 2013. "Host Restriction of Lentiviruses and Viral Countermeasures: APOBEC3 and Vif" Viruses 5, no. 8: 1934-1947. https://doi.org/10.3390/v5081934
APA StyleJónsson, S. R., & Andrésdóttir, V. (2013). Host Restriction of Lentiviruses and Viral Countermeasures: APOBEC3 and Vif. Viruses, 5(8), 1934-1947. https://doi.org/10.3390/v5081934