Enhanced Heterosexual Transmission Hypothesis for the Origin of Pandemic HIV-1

Abstract
:
1. Introduction
2. Study of Risk Factors
2.1. The Patchy Prevalence of SIVcpz and SIVgor
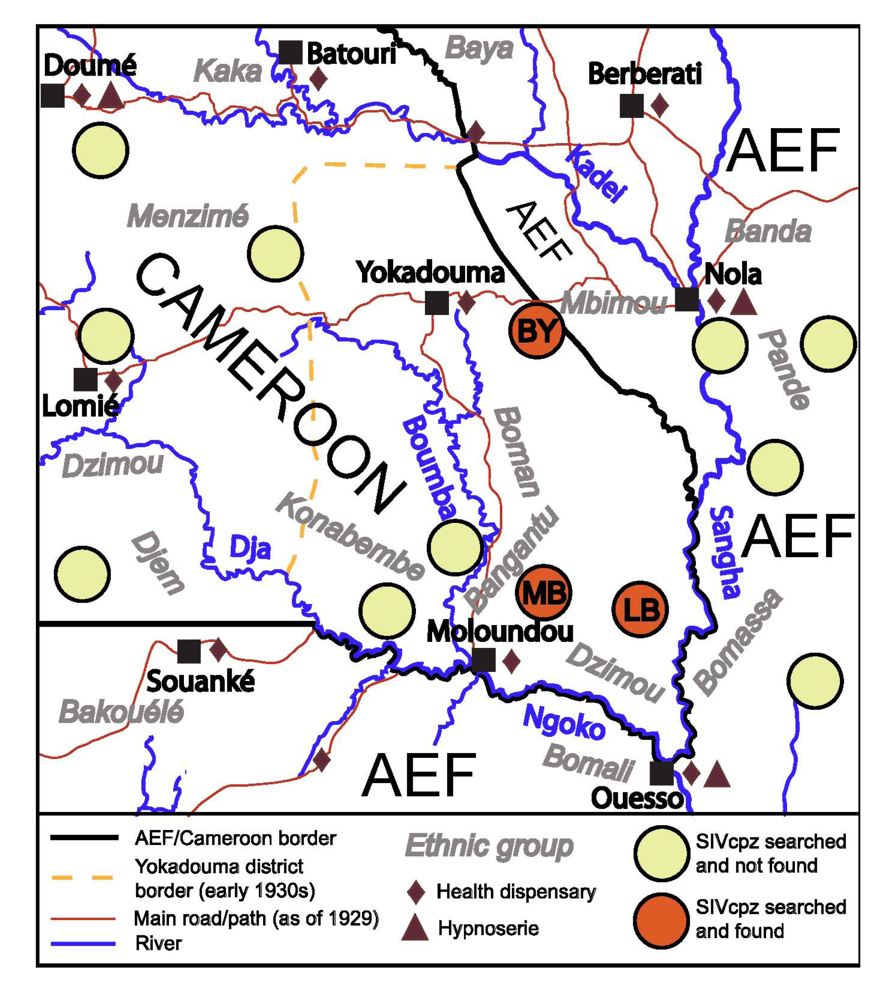
2.2. The Region under Study
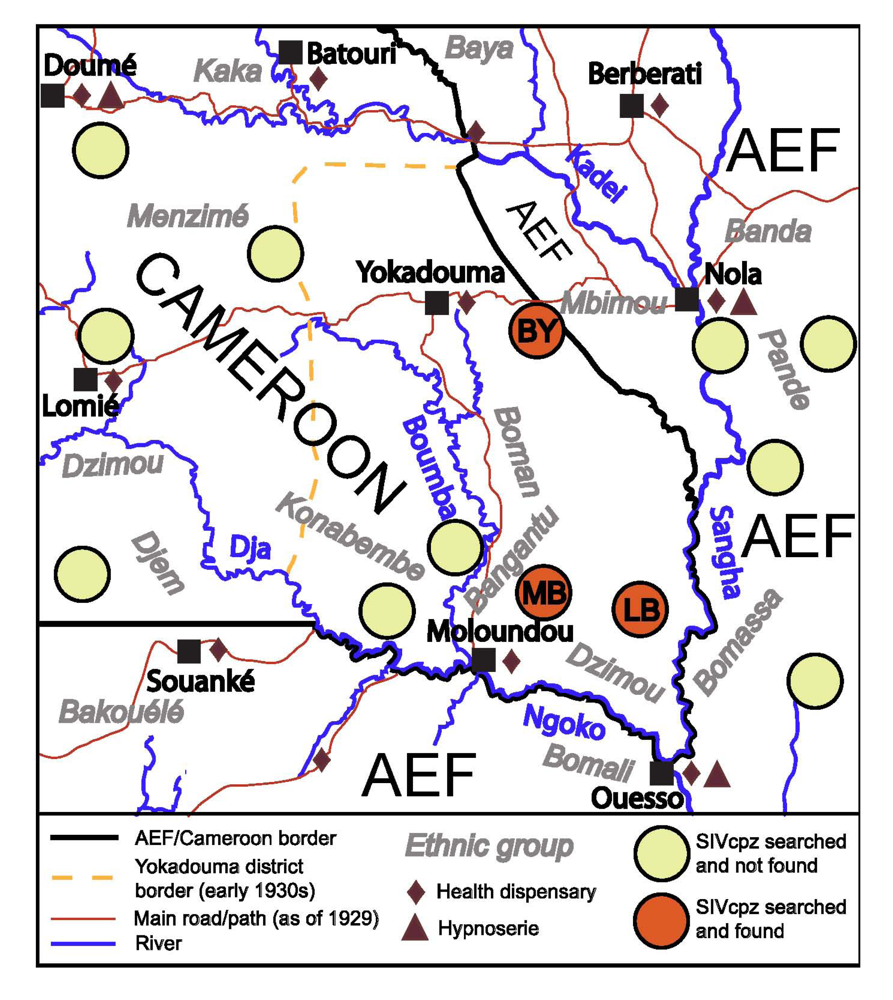
2.3. Evolution of Ape Hunting Practices over Time
2.4. Nosocomial Risks
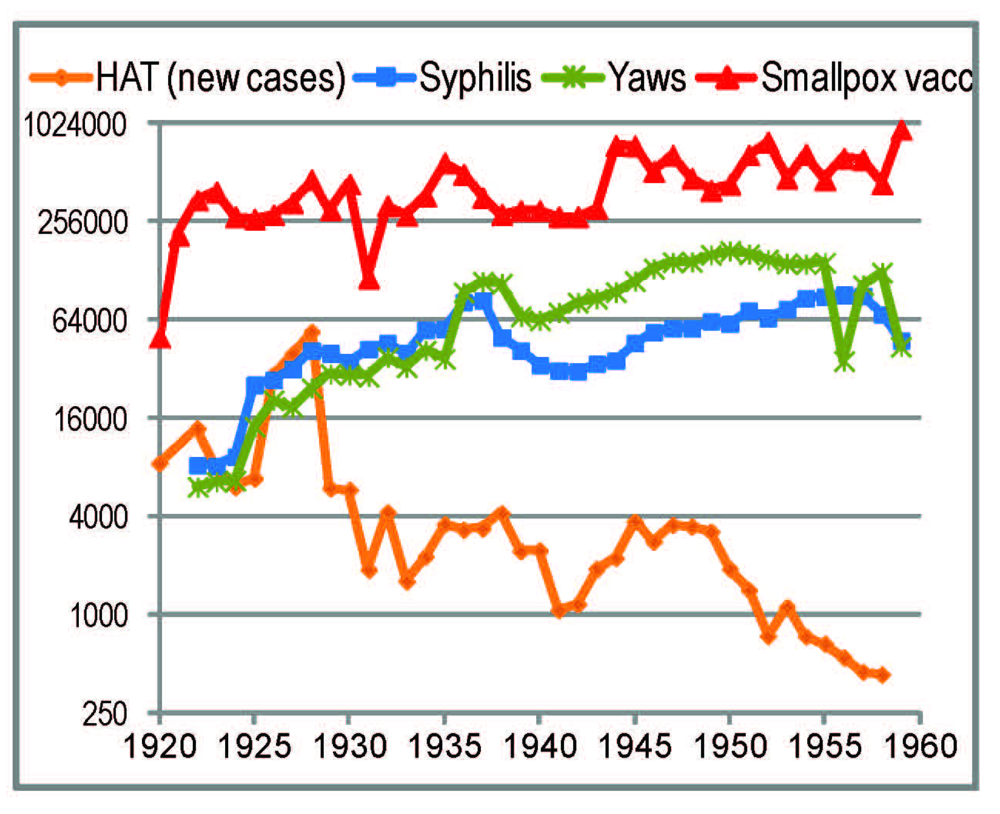
2.4.1. General Trends

2.4.2. Smallpox Vaccinations
2.4.3. Yaws and Syphilis Treatments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | # of treponematoses cases | # of injections | # of injections/patient/year | ||||
|---|---|---|---|---|---|---|---|
| Syphilis | Yaws | Neosalvarsan | Mercury salts | Sulfarsenol | Neosalvarsan | Others | |
| 1922 | 8,285 | 6,129 | 27,800 | NA | NA | 1.93 | NA |
| 1923 | 8,237 | 6,552 | 20,050 | NA | NA | 1.36 | NA |
| 1927 | 32,303 | 18,930 | 59,200 | 15,274 | 22,310 | 1.16 | 0.73 |
| 1928 | 42,015 | 24,718 | 76,270 | 18,158 | 59,450 | 1.14 | 1.16 |
| 1929 | 40,341 | 30,031 | 75,567 | 20,128 | 4,731 | 1.07 | 0.35 |
| 1930 | 35,693 | 29,701 | 77,728 | 23,085 | 15,591 | 1.19 | 0.59 |
| 1931 | 42,730 | 29,086 | 106,669 | 27,800 | 32,680 | 1.49 | 0.84 |
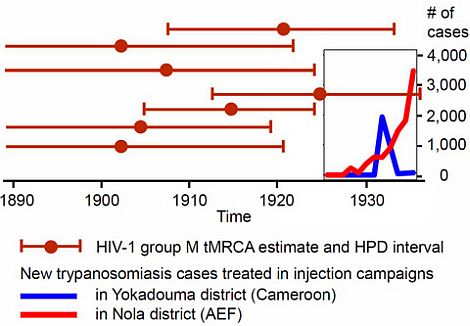
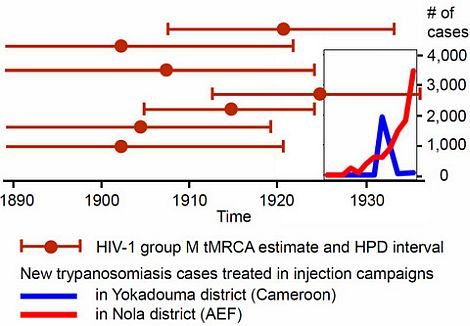
2.4.4. The HAT Campaigns
| Year | # of surveyed HAT cases | # of injections performed | # of injections per patient per year | |||
|---|---|---|---|---|---|---|
| New cases | Old cases | All surveyed cases | New cases (all treated) | Old cases | ||
| 1930 | 12,649 | 34,611 | 144,168 | 3.05 | 6-8? | 1-2? |
| 1934 | 13,368 | 42,508 | 254,000 | 4.55 | ~12 | 2.20 |
| 1935 | 12,557 | 54,709 | 262,424 | 3.90 | ~12 | 2.75 |
| 1937 | 16,597 | 60,196 | 597,409 | 7.78 | ~12 | 6.62 |
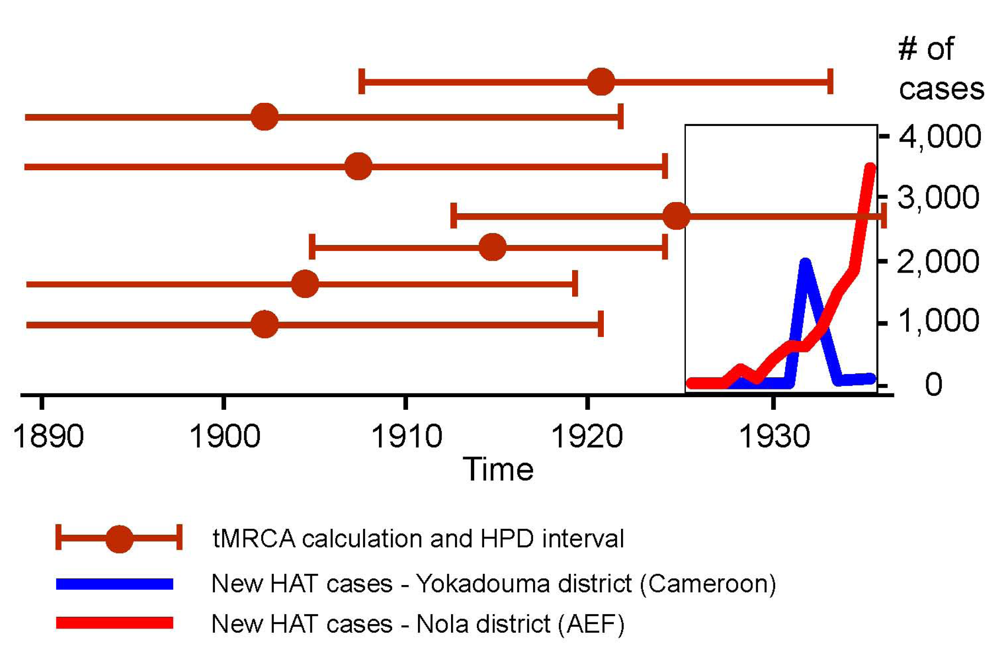
2.4.5. The Early 1930s: Intensification of Injection Treatments
| Year | Yokadouma sector a (Cameroon) | Nola sector (AEF) b | Ouesso sector (AEF) b | |||
|---|---|---|---|---|---|---|
| New cases | Surveyed old cases | New cases | Surveyed old cases | New cases | Surveyed old cases | |
| 1927 | – | – | – | – | – | – |
| 1928 | – | – | 228 | 222 | 108 | ? |
| 1929 | – | – | 77 | 176 | 212 | 242 |
| 1930 | – | – | 380 | ? | 154 | 356 |
| 1931 | – | – | 594 | 2,371 | 116 | 457 |
| 1932 | 2,928 | – | 590 | 1,812 | 130 | 957 |
| 1933 | ? | ? | 885 | 3,409 | 177 | 1,632 |
| 1934 | 43 | 2,928 | 1,463 | 2,516 | 184 | 818 |
| 1935 | ? | ? | 1,816 | 7,342 | b | b |
| 1936 | 74 | 1,036 | 3,464 | 3,721 | b | b |

2.4.6. Health Facilities
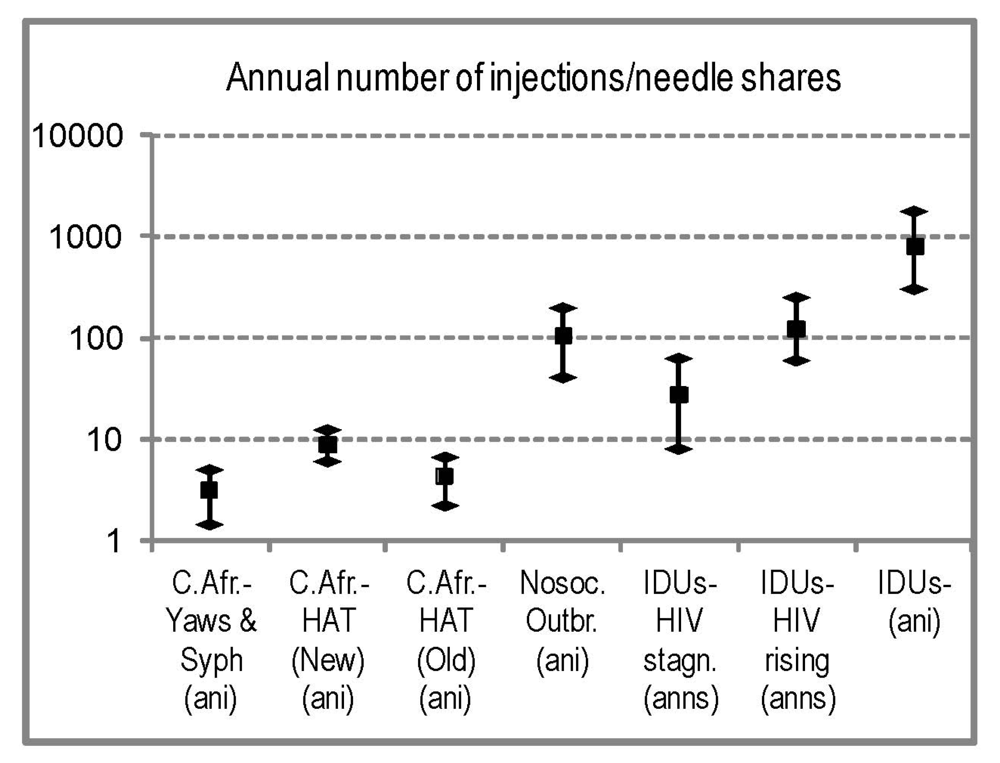
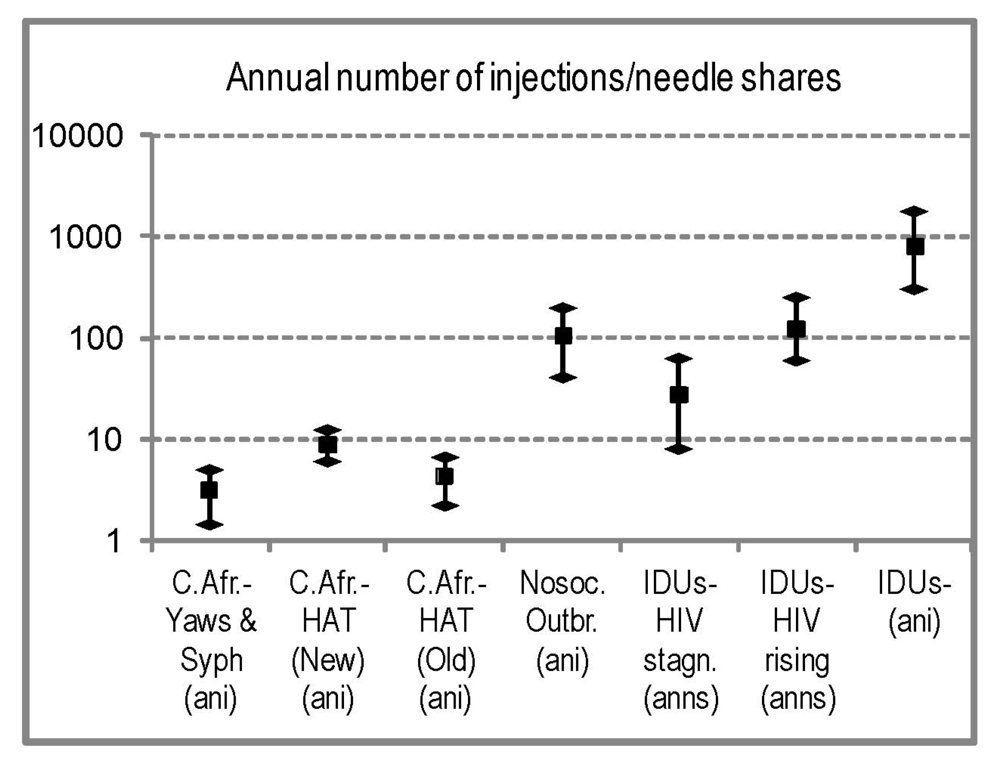
2.4.7. Comparison with Modern HIV Parenteral Outbreaks
2.5. Sexual Promiscuity and Sexually Transmitted Diseases
2.6. Travelling and Migration of People to/from the Stanley Pool Cities
3. Discussion
4. Sources Used
5. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Van Heuverswyn, F.; Li, Y.; Neel, C.; Bailes, E.; Keele, B.F.; Liu, W.; Loul, S.; Butel, C.; Liegeois, F.; Bienvenue, Y.; et al. SIV infection in wild gorillas. Nature 2006, 444, 164. [Google Scholar] [CrossRef]
- Keele, B.F.; Van Heuverswyn, F.; Li, Y.; Bailes, E.; Takehisa, J.; Santiago, M.L.; Bibollet-Ruche, F.; Chen, Y.; Wain, L.V.; Liegeois, F.; et al. Chimpanzee Reservoirs of Pandemic and Nonpandemic HIV-1. Science 2006, 313, 523–526. [Google Scholar]
- Hahn, B.H.; Shaw, G.M.; De Cock, K.M.; Sharp, P.M. AIDS as a zoonosis: Scientific and public health implications. Science 2000, 287, 607–614. [Google Scholar] [CrossRef]
- Marx, P.A.; Alcabes, P.G.; Drucker, E. Serial human passage of simian immunodeficiency virus by unsterile injections and the emergence of epidemic human immunodeficiency virus in Africa. Phil. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 911–920. [Google Scholar] [CrossRef]
- Chitnis, A.; Rawls, D.; Moore, J. Origin of HIV-1 in colonial French Equatorial Africa? AIDS Res. Hum. Retrovir. 2000, 16, 5–8. [Google Scholar] [CrossRef]
- Lemey, P.; Pybus, O.G.; Wang, B.; Saksena, N.K.; Salemi, M.; Vandamme, A.-M. Tracing the origin and history of the HIV-2 epidemic. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 6588–6592. [Google Scholar]
- Yamaguchi, J.; Devare, S.G.; Brennan, C.A. Identification of a new HIV-2 subtype based on phylogenetic analysis of full-length genomic sequence. AIDS Res. Hum. Retrovir. 2000, 16, 925–930. [Google Scholar]
- Damond, F.; Worobey, M.; Campa, P.; Farfara, I.; Colin, G.; Matheron, S.; Brun-Vézinet, F.; Robertson, D.L.; Simon, F. Identification of a highly divergent HIV type 2 and proposal for a change in HIV type 2 classification. AIDS Res. Hum. Retrovir. 2004, 20, 666–672. [Google Scholar]
- Santiago, M.L.; Range, F.; Keele, B.F.; Li, Y.; Bailes, E.; Bibollet-Ruche, F.; Fruteau, C.; Noë, R.; Peeters, M.; Brookfield, J.F.Y.; et al. Smian Immunodeficiency Virus Infection in Free-Ranging Sooty Mangabeys (Cercocebus atys atys) from the Tai Forest, Côte d’Ivoire: Implications for the Origin of Epidemic Human Immunodeficiency Virus Type 2. J. Virol. 2005, 79, 12515–12527. [Google Scholar]
- Plantier, J.C.; Leoz, M.; Dickerson, J.E.; De Oliveira, F.; Cordonnier, F.; Lemée, V.; Damond, F.; Robertson, D.L.; Simon, F. A new human immunodeficiency virus derived from gorillas. Nat. Med. 2009, 15, 871–872. [Google Scholar]
- Vallari, A.; Holzmayer, V.; Harris, B.; Yamaguchi, J.; Ngansop, C.; Makamche, F.; Mbanya, D.; Kaptué, L.; Ndembi, N.; Gürtler, L.; et al. Confirmation of putative HIV-1 group P in Cameroon. J. Virol. 2011, 85, 1403–1407. [Google Scholar]
- Ayouba, A.; Akoua-Koffi, C.; Adjogoua, E.; Calvignac, S.; Esteban, A.; Locatelli, S.; Delaporte, E.; Leendertz, F.; Peeters, M. Identification of a new HIV-2 lineage in rural Côte d’Ivoire: An additional cross-species transmission from SIVsmm from sooty mangabeys to humans. In Proceedings of the 19th Conference on Retroviruses and Opportunistic Infections, Seattle, WA, USA, 5–8 March 2012. Oral presentation 62LB.
- Neel, C.; Etienne, L.; Li, Y.; Takehisa, J.; Rudicell, R.S.; Ndong Bass, I.; Moudindo, J.; Mebenga, A.; Esteban, A.; Van Heuverswyn, F.; et al. Molecular epidemiology of simian immunodeficiency virus infection in wild-living gorillas. J. Virol. 2010, 84, 1464–1476. [Google Scholar] [CrossRef]
- Mauclère, P.; Loussert-Ajaka, I.; Damond, F.; Fagot, P.; Souquières, S.; Lobe, M.M.; Mbopi Keou, F.-X.; Barré-Sinoussi, F.; Saragosti, S.; Brun-Vézinet, F.; et al. Serological and virological characterization of HIV-1 group O infection in Cameroon. AIDS 1997, 11, 445–453. [Google Scholar] [CrossRef]
- Sharp, P.M.; Hahn, B.H. Origins of HIV and the AIDS pandemic. Cold Spring Harb. Perspect. Med. 2011, 1, a006841. [Google Scholar]
- Sousa, J.D.; Müller, V.; Lemey, P.; Vandamme, A.-M. High GUD incidence in the early 20th century created a particularly permissive time window for the origin and initial spread of epidemic HIV strains. PLoS One 2010, 5, e9936. [Google Scholar]
- Pépin, J.; Labbé, A.C. Noble goals, unforeseen consequences: Control of tropical diseases in colonial Central Africa and the iatrogenic transmission of blood-borne viruses. Trop. Med. Int. Health 2008, 13, 1–10. [Google Scholar]
- Pépin, J. The Origins of AIDS; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Kalish, M.L.; Wolfe, N.D.; Ndongmo, C.B.; McNicholl, J.; Robbins, K.E.; Aidoo, M.; Fonjungo, P.N.; Alemnji, G.; Zeh, C.; Djoko, C.F.; et al. Central African hunters exposed to simian immunodeficiency virus. Emerg. Infect. Dis. 2005, 11, 1928–1930. [Google Scholar] [CrossRef]
- Wolfe, N.D.; Switzer, W.M.; Carr, J.K.; Bhullar, V.B.; Shanmugam, V.; Tamoufe, U.; Prosser, A.T.; Torimiro, J.N.; Wright, A.; Mpoudi-Ngole, E.; et al. Naturally acquired simian retrovirus infections in central African hunters. Lancet 2004, 363, 932–937. [Google Scholar]
- Wolfe, N.D.; Heneine, W.; Carr, J.K.; Garcia, A.D.; Shanmugam, V.; Tamoufe, U.; Torimiro, J.N.; Prosser, A.T.; LeBreton, M.; Mpoudi-Ngole, E.; et al. Emergence of unique primate T-lymphotropic viruses among central African bushmeat hunters. Proc. Nat. Acad. Sci. U. S. A. 2005, 102, 7994–7999. [Google Scholar]
- Salemi, M.; Van Dooren, S.; Vandamme, A.-M. Origin and evolution of human and simian T-cell lymphotropic viruses. AIDS Rev. 1999, 1, 131–139. [Google Scholar]
- Van Dooren, S.; Salemi, M.; Vandamme, A.-M. Dating the origin of the african human T-cell lymphotropic virus type-I (HTLV-I) subtypes. Mol. Biol. Evol. 2001, 18, 661–671. [Google Scholar]
- Salemi, M.; Desmyter, J.; Vandamme, A.-M. Tempo and mode of human and simian T-lymphotropic virus (HTLV/STLV) evolution revealed by analyses of full-genome sequences. Mol. Biol. Evol. 2000, 17, 374–386. [Google Scholar] [CrossRef]
- Salemi, M.; Strimmer, K.; Hall, W.W.; Duffy, M.; Delaporte, E.; Mboup, S.; Peeters, M.; Vandamme, A.-M. Dating the radiation of HIV-1 group M in 1930s using a new method to uncover clock-like molecular evolution. FASEB J. Print 2001, 15, 276–278. [Google Scholar]
- Korber, B.; Muldoon, M.; Theiler, J.; Gao, F.; Gupta, R.; Lapedes, A.; Hahn, B.H.; Wolinsky, S.; Bhattacharya, T. Timing the ancestor of the HIV-1 pandemic strains. Science 2000, 288, 1789–1796. [Google Scholar]
- Worobey, M.; Gemmel, M.; Teuwen, D.E.; Haselkorn, T.; Kunstman, K.; Bunce, M.; Muyembe, J.-J.; Kabongo, J.M.M.; Kalengayi, R.M.; Van Marck, E.; et al. Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960. Nature 2008, 455, 661–665. [Google Scholar]
- Lemey, P.; Pybus, O.G.; Rambaut, A.; Drummond, A.J.; Robertson, D.L.; Roques, P.; Worobey, M.; Vandamme, A.-M. The molecular population genetics of HIV-1 group O. Genetics 2004, 167, 1059–1068. [Google Scholar]
- Wertheim, J.O.; Worobey, M. Dating the age of the SIV lineages that gave rise to HIV-1 and HIV-2. PLoS Comput. Biol. 2009, 5, e1000377. [Google Scholar]
- Wertheim, J.O; Fourment, M.; Kosakovsky Pond, S.L. Inconsistencies in estimating the age of HIV-1 subtypes due to heterotachy. Mol. Biol. Evol. 2012, 29, 451–456. [Google Scholar] [CrossRef]
- Faria, N.R.; Hodges-Mameletzis, I.; Silva, J.C.; Rodés, B.; Erasmus, S.; Paolucci, S.; Ruelle, J.; Pieniazek, D.; Taveira, N.; Treviño, A.; et al. Pylogeographical footprint of colonial history in the global dispersal of human immunodeficiency virus type 2 group A. J. Gen. Virol. 2012, 93, 889–899. [Google Scholar] [CrossRef]
- Drucker, E.; Alcabes, P.G.; Marx, P.A. The injection century: Massive unsterile injections and the emergence of human pathogens. Lancet 2001, 358, 1989–1992. [Google Scholar]
- Verdraeger, J. SIDA: Hypothèses sur l'origine et l'émergence du VIH (in French). Bull. Soc. Pathol. Exótique 1995, 88, 54–60. [Google Scholar]
- Gisselquist, D. Emergence of HIV type 1 epidemic in the twentieth century: Comparing hypotheses to evidence. AIDS Res. Hum. Retrovir. 2003, 19, 1071–1078. [Google Scholar]
- Pépin, J.; Annie-Claude Labbé, A.C.; Mamadou-Yaya, F.; Mbélesso, P.; Mbadingaï, S.; Deslandes, S.; Locas, M.-C.; Frost, E. Iatrogenic transmission of human T cell lymphotropic virus type 1 and hepatitis C virus through parenteral treatment and chemoprophylaxis of sleeping sickness in Colonial Equatorial Africa. Clin. Inf. Dis. 2010, 51, 777–784. [Google Scholar]
- Cameron, D.W.; Simonsen, J.N.; D'Costa, L.J.; Ronald, A.R.; Maitha, G.M.; Gakinya, M.N.; Cheang, M.; Ndinya-Achola, J.O.; Piot, P.; Brunham, R.C.; et al. Female to male transmission of human immunodeficiency virus type 1: Risk factors for seroconversion in men. Lancet 1989, 2, 403–407. [Google Scholar]
- Hayes, R.J.; Schulz, K.F.; Plummer, F.A. The cofactor effect of genital ulcers on the per-exposure risk of HIV transmission in sub-Saharan Africa. J. Trop. Med. Hyg. 1995, 98, 1–8. [Google Scholar]
- Rémy, G. Image geographique de l’infection à VIH-1 en Afrique Centrale: Des descontinuités remarquables (in French). Ann. Soc. Belge Méd. Trop. 1993, 73, 127–142. [Google Scholar]
- Hooper, E. The River: A Journey to the Source of HIV and AIDS; Little, Brown, and Company: London, UK, 2000. [Google Scholar]
- Hooper, E. Dephlogistication, imperial display, apes, angels, and the return of Monsieur Émile Zola. Atti dei Convegni Lincei 2003, 187, 27–230. [Google Scholar]
- Vidal, N.; Peeters, M.; Mulanga-Kabeya, C.; Nzilambi, N.; Robertson, D.; Ilunga, W.; Sema, H.; Tshimanga, K.; Bongo, B.; Delaporte, E. Unprecedented degree of human immunodeficiency virus type 1 (HIV-1) group M genetic diversity in the Democratic Republic of Congo suggests that the HIV-1 pandemic originated in Central Africa. J. Virol. 2000, 74, 10498–10507. [Google Scholar]
- Vidal, N.; Mulanga, C.; Bazepeo, S.E.; Mwamba, J.K.; Tchimpaka, J.W.; Kashi, M.; Mama, N.; Laurent, C.; Lepira, F.; Delaporte, E.; et al. Distribution of HIV-1 variants in the Democratic Republic of Congo suggests increase of subtype C in Kinshasa between 1997 and 2002. J. Acquir. Immune Defic. Syndr. 2005, 40, 456–462. [Google Scholar] [CrossRef]
- Lemey, P. Early spread of HIV-1 M in the DRC. In Proceedings of the International Symposium on Simian Viruses and Emerging Diseases in Humans, Paris, France, 9–10 June 2010. Oral presentation.
- Van Heuverswyn, F.; Li, Y.; Bailes, E.; Neel, C.; Lafay, B.; Keele, B.F.; Shaw, K.S.; Takehisa, J.; Kraus, M.H.; Loul, S.; et al. Genetic diversity and phylogeographic clustering of SIVcpzPtt in wild chimpanzees in Cameroon. Virology 2007, 368, 155–171. [Google Scholar] [CrossRef]
- Li, Y.; Ndjango, J.-B.; Learn, G.; Robertson, J.; Takehisa, J.; Bibollet-Ruche, F.; Sharp, P.; Worobey, M.; Shaw, G.; Hahn, B. Molecular epidemiology of simian immunodeficiency virus in Eastern Chimpanzees and Gorillas. In Proceedings of the 17th Conference on Retroviruses and Opportunistic Infections, San Francisco, NC, USA, 16–19 February 2010. Poster presentation B-108.
- Peterson, D. Eating Apes; University of California Press: Berkeley, CA, USA, 2003. [Google Scholar]
- Merfield, F.G. Gorillas Were My Neighbours; The Company Book Club: London, UK, 1957. [Google Scholar]
- Meder, A. Gorillas in African culture and medicine. Gorilla J. 1999, 18, 11–15. [Google Scholar]
- Florian Liégeois, F.; Boué, V.; Locatelli, S.; Butel, C.; Mouinga-Ondeme, A.; Delaporte, E.; Gonzalez, J.-P.; Peeters, M.; Rouet, F. Identification of new divergent SIVcpz Ptt strains in wild living chimpanzees in Gabon. In Proceedings of the 19th Conference on Retroviruses and Opportunistic Infections, Seattle, WA, USA, 5–8 March 2012. Poster presentation B-109.
- Cameroon, Rapport annuel adressé par le Gouvernement Français au Conseil de la Société des Nations sur l'administration sous mandat du territoire du Cameroun pour l'année[1921 to 1939] (in French); Imprimerie Générale Lahure: Paris, France, 1921.
- French Congo, Rapport annuel du Chef du Service de Santé, Colonie du Moyen-Congo, Afrique Équatoriale Française [1930 through 1934] (in French); Service de Santé: Brazzaville, French Congo, 1930; IMTSSA, Box 117.
- Pollacchi, P.; Lyautey, H. Atlas colonial français: colonies, protectorats, et pays sous mandat (in French); L'Illustration: Paris, France, 1929. [Google Scholar]
- Bruel, G. La France Équatoriale Africaine: le pays, les habitants, la colonization, les pouvoirs publics (in French); Larose Éditeur: Paris, France, 1935. [Google Scholar]
- Koch, C.W.H. Die Stämme des Bezirks Molundu: in Sprachlicher, Geschichtlicher und Völkerkundlicher Beziehung (in German). Bässler-Archiv: Beiträge zu Völkerkunde 1913, 3, 257–312. [Google Scholar]
- Giles-Vernick, T. We wander like birds: Migration, indigeneity, and the fabrication of frontiers in the Sangha River Basin of Equatorial Africa. Environ. Hist. 1999, 4, 168–197. [Google Scholar] [CrossRef]
- Giles-Vernick, T. Cutting the Vines of the Past: Environmental Histories of the Central African Rain Forest; University Press of Virginia: Charlottesville, VA, USA, 2002. [Google Scholar]
- Despois, J. Les genres de vie des populations de la forêt dans le Cameroun oriental (in French). Ann. Géographie 1946, 55, 19–38. [Google Scholar] [CrossRef]
- Hattori, S. Nature conservation and hunter-gatherer’s life in Cameroonian rainforest. Afr. Study Monogr. 2005, 29, 41–51. [Google Scholar]
- Köhler, A. Of Apes and men: Baka and Bantu attitudes to wildlife and the making of eco-goodies and baddies. Conservat. Soc. 2005, 3, 407–435. [Google Scholar]
- Allys, P. Monographie de la tribu des Dzems (Ngoko-Sangha) (in French). Bull. Soc. Rech. Congolaises 1930, 11, 3–21. [Google Scholar]
- Ballif, N. Les Pygmées de la Grande Forêt (in French); L’Harmattan: Paris, France, 1992. [Google Scholar]
- Darré, E.; Le Bourhis, A. Notes sur la tribu Bomitaba (in French). Bull. Soc. Rech. Congolaises 1925, 5, 15–38. [Google Scholar]
- Blake, S. A reconnaissance survey in the Likouala swamps of northern Congo and its implications for conservation.
- Kano, T.; Asato, R. Hunting pressure on chimpanzees and gorillas in the Motaba river area, northeastern Congo. Afr. Study Monogr. 1994, 15, 143–162. [Google Scholar]
- Lewis, J. Forest hunther-gatherers and their world: A study of the Mbendjele Yaka pygmies of Congo-Brazzaville and their secular and religious activities and representations.
- Lalouel, J. Les Babinga du Bas-Oubangui. Contribution à l'étude ethnographique des Négrilles, Bakà et Bayakà (in French). Bull. et Mémoires de la Soc. d'Anthrop. de Paris 1950, 1, 175–211. [Google Scholar] [CrossRef]
- Giles-Vernick, T.; Rupp, S. Visions of apes, reflections on change: Telling tales of great apes in Equatorial Africa. Afr. Studies Rev. 2006, 49, 51–73. [Google Scholar] [CrossRef]
- Giles-Vernick, T. Apes and Humans in the Sangha basin, 1890–1920. In Proceedings of the International Symposium on Simian Viruses and Emerging Diseases in Humans, Paris, France, 9–10 June 2010. Oral presentation.
- Coquery-Vidrovitch, C. La Haute-Sangha au Temps des Compagnies Concessionnaires (in French). Yale F&ES Bull. 1999, 102, 78–90. [Google Scholar]
- Timberg, C.; Halperin, D. Tinderbox: How the West Sparked the AIDS Epidemic and How the World Can Finally Overcome It; The Penguin Press: New York, NY, USA, 2012. [Google Scholar]
- Fargeot, C. La Chasse Commerciale et le Négoce de la Venaison en Afrique Centrale Forestière (in French). Game Wildl. Sci. 2004, 21, 817–833. [Google Scholar]
- Headrick, R. Colonialism, Health and Illness in French Equatorial Africa, 1885–1935; African Studies Associated Press: Atlanta, GA, USA, 1994. [Google Scholar]
- Société d'Éditions Géographiques, Maritimes et Coloniales. In Guide des Colonies Françaises: Afrique Française: Afrique Occidentale Française, Afrique Équatoriale Française, Togo et Cameroun (in French); Société d'Éditions Géographiques, Maritimes et Coloniales: Paris, France, 1931.
- Frechkop, S. Mammifères et oiseaux protégés au Congo Belge (in French); Institut des Parcs Nationaux du Congo Belge: Brussels, Belgium, 1936. [Google Scholar]
- Frechkop, S. Animaux Protégés au Congo Belge et dans le Territoire sous Mandat du Ruanda-Urundi (in French); Institut des Parcs Nationaux du Congo Belge: Brussels, Belgium, 1941. [Google Scholar]
- Gagneux, P.; Gonder, M.K.; Goldberg, T.L.; Morin, P.A. Gene flow in wild chimpanzee populations: what genetic data tell us about chimpanzee movement over space and time. Phil. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 889–897. [Google Scholar] [CrossRef]
- Walsh, P.D.; Abernethy, K.A.; Bermejo, M.; Beyersk, R.; De Wachter, P.; Akou, M.E.; Huijbregts, B.; Mambounga, D.I.; Toham, A.K.; Kilbourn, A.M.; et al. Catastrophic ape decline in western equatorial Africa. Nature 2003, 422, 611–614. [Google Scholar] [CrossRef]
- Bailey, N.D. Bushmeat Crisis Task Force, Bushmeat Crisis Task Force. Effects of Bushmeat Hunting on Populations of African Great Apes: Summary Document; Bushmeat Crisis Task Force: Silver Spring, MD, USA, 2001. [Google Scholar]
- Butynsky, T.M. The robust chimpanzee Pan troglodytes: Taxonomy, distribution, abundance and conservation status. In Status Survey and Conservation Action Plan: West African Chimpanzees; Kormos, R., Boesch, C., Bakarr, M.I., Butynski, T.M., Eds.; IUCN/SSC Primate Specialist Group: Gland, Switzerland and Cambridge, UK, 2003; pp. 5–12. [Google Scholar]
- De Goyon, J. Variole et vaccinations dans le Haut Oubangui (in French). Ann. Hyg. Méd. Colon. 1996, 9, 117–125. [Google Scholar]
- Jouveau-Dubreil, H. Service de la vaccine à l'Institut Pasteur de Tchen-Tou (in French). Ann. Méd. Pharm. Colon. 1921, 19, 64–77. [Google Scholar]
- Legendre, J. Les épingles pour la vaccination antivariolique en série (in French). Ann. Méd. Pharm. Colon. 1924, 22, 412–413. [Google Scholar]
- Aubert, P. Mission dans la Haute-Sangha: étude de la dysenterie, de la trypanosomiase humaine, etc. (in French). Ann. Hyg. Méd. Colon. 1911, 14, 782–816. [Google Scholar]
- Heckenroth, F.F. La vaccine dans la Haute-Sangha (Congo Français) (in French). Ann. Hyg. Méd. Colon. 1909, 12, 402–414. [Google Scholar]
- Bouillez, M. Vaccine et vaccination au Tchad, secteur médical de Bokoro (27 février–28 novembre 1907) (in French). Ann. Hyg. Méd. Colon. 1908, 11, 475–487. [Google Scholar]
- Lecompte, A. Rapport sur les opérations de vaccination pratiquées dans l'Ogooué (Congo Français) en 1896–1897 (in French). Ann. Hyg. Méd. Colon. 1899, 2, 98–103. [Google Scholar]
- Rousseau, P. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l’année 1927 (in French). Ann. Méd. Pharm. Colon. 1929, 27, 145–246. [Google Scholar]
- Hermant, P. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l’année 1928 (in French). Ann. Méd. Pharm. Colon. 1931, 29, 5–138. [Google Scholar]
- Ledentu, G. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l’année 1929 (in French). Ann. Méd. Pharm. Colon. 1931, 29, 661–849. [Google Scholar]
- Lefèvre, R. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l’année 1930 (in French). Ann. Méd. Pharm. 1932, 30, 296–405 and 546–569. [Google Scholar]
- Marque, E. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l'année 1931 (in French). Ann. Méd. Pharm. Colon. 1933, 31, 123–323. [Google Scholar]
- Grosfillez, J. Les principales maladies observées dans les colonies françaises et territoires sous mandat en 1932 (in French). Ann. Méd. Pharm. Colon. 1934, 32, 153–268. [Google Scholar]
- Ledentu, G. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l’année 1933 (in French). Ann. Méd. Pharm. Colon. 1935, 33, 552–816. [Google Scholar]
- Ledentu, G. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l'année 1934 (in French). Ann. Méd. Pharm. Colon. 1936, 34, 474–749. [Google Scholar]
- Ledentu, G.; Peltier, M. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l’année 1935 (in French). Ann. Méd. Pharm. Colon. 1937, 35, 748–928 and 1207–1335. [Google Scholar]
- Vogel, E.; Le Rouzic, J. Les maladies transmissibles observées dans les colonies françaises et territoires sous mandat pendant l'année 1936 (in French). Ann. Méd. Pharm. Colon. 1938, 36, 352–520 and 633–725. [Google Scholar]
- Vogel, E.; Riou, M. Les maladies épidémiques, endémiques, et sociales dans les colonies françaises pendant l’année 1937 (in French). Ann. Méd. Pharm. Colon. 1939, 37, 257–551. [Google Scholar]
- Clapier, N. L'endémie pianique sur le Bas-Oubangui: essai de lutte anti-pianique, 1920 (in French). Ann. Méd. Pharm. Colon. 1921, 19, 319–334. [Google Scholar]
- Cameroon, Rapport du Service de Santé, Division du Wouri, Territoire du Cameroun (in French); Service de Santé: Douala, Cameroon, 1935; IMTSSA, Box 481.
- Dupuy, L. Rapport sur le Service de l'Assistance Médicale aux Indigènes du Foréami pour l'exercice 1931 (in French); Foréami: Brussels, Belgium, 1932. [Google Scholar]
- Croix Rouge du Congo, Rapport annuel [1928 through 1954] (in French); Croix Rouge de Belgique: Brussels, Belgium; pp. 1928–1954.
- Janssens, P.G.; Yaws, M.A. Health in Central Africa since 1885: Past, Present and Future; Janssens, P.G., Kivits, M., Vuylsteke, J., Eds.; Fondation Roi Baudouin: Brussels, Belgium, 1997; pp. 1829–1839. [Google Scholar]
- Vernier, P. Épidemiologie Vénérienne en Afrique Noire Française (in French); Imprimerie Bière: Bordeaux, France, 1950. [Google Scholar]
- Tournier, E. Le traitement du pian par les injections sous-cutanées de novarsénobenzol (in French). Ann. Méd. Pharm. Colon. 1922, 20, 37–39. [Google Scholar]
- Letonturier, L.P. Rapport sur le fonctionnement du service de santé au Cameroun pendant l’année 1923 (in French). Ann. Méd. Pharm. Colon. 1924, 22, 396–408. [Google Scholar]
- Broden, A.; Rodhain, J. Rapport sur les Travaux Faits au Laboratoire de la Société Belge d'Études Coloniales à l'Hôpital des Noirs et au Lazaret pour Trypanosomiés (in French); Société Belge d'Études Coloniales: Brussels, Belgium, 1908. [Google Scholar]
- Bauche, M. Trypanosomen und Tinbeef—Medizinisches Wissen um Schlafkrankheit zwischen Kamerun und Deutschland, 1910–1914. In Beiträge zur I. Kölner Afrikawissenschaftlichen Nachwuchstagung (in German); Seifert, M., Egert, M., Heerbaart, F., Kolossa, K., Limanski, M., Mumin, M., Rodekuhr, P.A., Rous, S., Stankowski, S., Thanassoula, M., Eds.; Universität Köln: Köln, Germany, 2005; pp. 1–19. [Google Scholar]
- Kerandel, J. Note sur la mission du Haut Logone (in French). Ann. Hyg. Méd. Colon. 1909, 12, 107–114. [Google Scholar]
- Heckenroth, F.F. Résumé de 68 observations de trypanosomiase humaine dans la Haute-Sangha (Congo) (in French). Ann. Hyg. Méd. Colon. 1910, 13, 663–669. [Google Scholar]
- Heckenroth, F.F. Tournée médicale effectuée sur le Congo et l'Oubangui (in French). Ann. Hyg. Méd. Colon. 1913, 16, 104–144. [Google Scholar]
- Ouzilleau, F. Rapport sur la tournée médicale effectuée en pays Yakoma du 23 mars au 3 avril 1912 (in French). Ann. Hyg. Méd. Colon. 1912, 15, 819–827. [Google Scholar]
- Montfort, F. Contribution à l'étude de la répartition et de la prophylaxie de la trypanosomiase humaine dans la Haute Sangha (avril-mai 1911) (in French). Ann Hyg Méd. Colon. 1912, 15, 593–613. [Google Scholar]
- Kuhn, P. Die Geschichte der Schlafkrankheit in Kamerun und ihre Lehren (in German). Med. Microbiol. Immunol. 1916, 81, 69–137. [Google Scholar]
- Boyé, L. Fonctionnement technique des secteurs de prophylaxie de la maladie du sommeil dans l'Afrique Équatoriale Française pendant l'année 1921 (in French). Ann. Méd. Pharm. Colon. 1922, 20, 194–216. [Google Scholar]
- Jamot, E. La maladie du sommeil au Cameroun en janvier 1929 (in French). Bull. Soc. Pathol. Exótique 1929, 22, 473–496. [Google Scholar]
- Jamot, E. La Maladie du Sommeil au Cameroun (in French). Africa: J. Int. Afr. Institute 1930, 3, 161–177. [Google Scholar] [CrossRef]
- Ollivier, G.; Legros, D. Trypanosomiase humaine africaine: Historique de la thérapeutique et de ses échecs (in French). Trop. Med. Int. Health 2001, 6, 855–863. [Google Scholar] [CrossRef]
- Millous, P. La lutte contre les maladies sociales au Cameroun en 1934 (in French). Africa: J. Int. Afr. Institute 1935, 8, 171–182. [Google Scholar] [CrossRef]
- Muraz, G.; Vaisseau, G. De divers types d'arséno-résistance dans les traitements actuels (A.E.F.) de la trypanosomiase humaine (in French). Bull. Soc. Pathol. Exótique 1932, 25, 260–280. [Google Scholar]
- Vamos, S. Traitement de trypanosomés dans un secteur du Moyen-Chari (A.E.F.): étude de 3,705 observations (in French). Bull. Soc. Pathol. Exótique 1936, 29, 1015–1022. [Google Scholar]
- Lotte, A.J. Historique du foyer de trypanosomiase de Nola (in French). Bull. Soc. Pathol. Exótique 1953, 46, 374–386. [Google Scholar]
- Aymérich, J. La Conquête du Cameroun: 1er Août 1914 – 20 Février 1916 (in French); Payot: Paris, France, 1933. [Google Scholar]
- Gisselquist, D.P. Estimating HIV-1 transmission efficiency through unsafe medical injections. Int. J. Sex. Transm. Dis. AIDS 2002, 13, 152–159. [Google Scholar]
- Gisselquist, D.P.; Upham, G.; Potterat, J.J. Efficiency of human immunodeficiency virus transmission through injections and other medical procedures: Evidence, estimates, and unfinished busines. Infect. Control Hosp. Epidemiol. 2006, 27, 944–952. [Google Scholar]
- Kaplan, E.H.; Heimer, R. A model-based estimate of HIV infectivity via needle sharing. J. Acq. Imm. Deficien. Syndr. 1992, 5, 1116–1118. [Google Scholar]
- Vickerman, P.; Hickman, M.; Judd, A. Modelling the impact of hepatitis C transmission of reducing syringe sharing: London case study. Int. J. Epidemiol. 2007, 36, 396–405. [Google Scholar] [CrossRef]
- Vickerman, P.; Platt, L.; Hawkes, S. Modelling the transmission of HIV and HCV among injecting drug users in Rawalpindi, a low HCV prevalence setting in Pakistan. Sex. Transm. Inf. 2009, 85, ii23–ii30. [Google Scholar]
- Lott, D.C.; Strain, E.C.; Brooner, R.K.; Bigelow, G.E.; Johnson, R.E. HIV risk behaviors during pharmacologic treatment for opioid dependence: A comparison of levomethadyl acetate, buprenorphine, and methadone. J. Subst. Abuse Treat. 2006, 31, 187–194. [Google Scholar] [CrossRef]
- Kwon, J.A.; Iversen, J.; Maher, L.; Law, M.G.; Wilson, D.P. The impact of needle and syringe programs on HIV and HCV transmissions in injecting drug users in Australia: A model-based analysis. J. Acquir. Immune Defic. Syndr. 2009, 51, 462–469. [Google Scholar]
- Jutras-Aswad, D.; Zang, G.; Bruneau, J. Cannabis use correlates of syringe sharing among injection drug users. Amer. J. Addictions 2010, 19, 231–237. [Google Scholar]
- Kipke, M.D.; Unger, J.B.; Palmer, R.F.; Edgington, R. Drug use, needle sharing, and HIV risk among injection drug-using street youth. Subst. Use Misuse 1996, 31, 1167–1187. [Google Scholar] [CrossRef]
- Ball, J.C.; Lange, W.R.; Myers, C.P.; Friedman, S.R. Reducing the risk of AIDS through methadone maintenance treatment. J. Health Soc. Behav. 1988, 29, 214–226. [Google Scholar]
- Caussy, D.; Weiss, S.H.; Blattner, W.A.; French, J.; Cantor, K.P.; Ginzburg, H.; Altman, R.; Goedert, J.J. Exposure factors for HIV-1 infection among heterosexual drug abusers in New Jersey treatment programs. AIDS Res. Hum. Retrovir. 1990, 6, 1459–1467. [Google Scholar]
- Chaisson, R.E.; Moss, A.R.; Onishi, R.; Osmond, D.; Carlson, J.R. Human Immunodeficiency Virus Infection in Heterosexual Intravenous Drug Users in San Francisco. Amer. J. Public Health 1987, 77, 169–172. [Google Scholar]
- Kretzschmar, M.; Wiessing, L.G. Modelling the spread of HIV in social networks of injecting drug users. AIDS 1998, 12, 801–811. [Google Scholar] [CrossRef]
- Allen, D.M.; Onorato, I.M.; Green, T.A. HIV infection in intravenous drug users entering drug treatment, United States, 1988 to 1989. Amer. J. Public Health 1992, 82, 541–546. [Google Scholar] [CrossRef]
- De Irala, J.; Bigelow, C.; McCusker, J.; Hindin, R.; Zheng, U. Reliability of self-reported human immunodeficiency virus risk behaviors in a residential drug treatment population. Amer. J. Epidemiol. 1996, 143, 725–732. [Google Scholar]
- Peters, A.; Davies, T.; Richardson, A. Multi-site samples of injecting drug users in Edinburgh: prevalence and correlates of risky injecting practices. Addiction 1998, 93, 253–267. [Google Scholar]
- Raboud, J.M.; Boily, M.C.; Rajeswaran, J.; O’Shaughnessy, M.V.; Schechter, M.T. The impact of needle-exchange programs on the spread of HIV among injection drug users: A simulation study. J. Urban Health 2003, 80, 302–320. [Google Scholar]
- Vickerman, P.; Miners, A.; Williams, J. Assessing the cost-effectiveness of interventions linked to needle and syringe programmes for injecting drug users: An economic modelling report. National Institute for Health and Clinical Excellence. Report published online, 2008. Available online: http://www.nice.org.uk/nicemedia/pdf/NSPEconomicmodellingFullreportJune08.pdf (accessed on 14 June 2012).
- Retel-Laurentin, A. Un Pays à la Dérive: une Societé en Régression Démographique: les Nzakara de l’Est Centrafricain (in French); Jean-Pierre Delarge: Paris, France, 1979. [Google Scholar]
- Mouchet, R. Yaws and syphilis among natives of the Belgian Congo. Kenya Med. J. 1926, 3, 242–245. [Google Scholar]
- Kermorgant, A. Aperçu sur les maladies vénériennes dans les colonies françaises (in French). Ann. Hyg. Méd. Colon. 1903, 6, 428–460. [Google Scholar]
- Egerton, F.C. African Majesty: A Record of Refuge at the Court of the King of Bangangté in the French Cameroons; George Routledge & Sons: London, UK, 1938. [Google Scholar]
- Gray, C. Colonial Rule and Crisis in Equatorial Africa: Southern Gabon ca. 1850–1940; The University of Rochester Press: Rochester, NY, USA, 2002. [Google Scholar]
- Patterson, K.D. The vanishing Mpongwe: European contact and demographic change in the Gabon river. J. Afr. Hist. 1975, 16, 217–238. [Google Scholar] [CrossRef]
- Phillips, R. Heterogeneous imperialism and the regulation of sexuality in British West Africa. J. Hist. Sexuality 2005, 14, 291–315. [Google Scholar] [CrossRef]
- Van Wing, J. Études Bakongo: Sociologie, Religion et Magie (in French); Van Campenhout: Brussels, Belgium, 1938. [Google Scholar]
- Trezenem, E. Notes ethnographiques sur les tribus Fan du Moyen-Ogooué (in French). J. Soc. Africanistes 1936, 6, 65–93. [Google Scholar]
- Georgelin, D. Notes médicales sur le Gabon: les facteurs de dépopulation (in French). Ann. Méd. Pharm. Colon. 1920, 19, 58–64. [Google Scholar]
- Robineau, C. Contribution à l'histoire du Congo: la domination européenne et l'example de Souanké (1900-1960) (in French). Cahiers d'Études Africaines 1967, 7, 300–344. [Google Scholar] [CrossRef]
- Spire, D. Rapport médicale de la mission Fourneau (in French). Ann. Hyg. Méd. Colon. 1902, 5, 353–379. [Google Scholar]
- Berny, P. La lèpre en Haute-Sangha (in French). Bull. Soc. Rech. Congolaises 1933, 18, 37–55. [Google Scholar]
- Kermorgant, A. Maladies endémiques, épidémiques et contagieuses qui ont regné dans les colonies françaises en 1906 (in French). Ann. Hyg. Méd. Colon. 1908, 11, 334–380. [Google Scholar]
- Adler, M.; French, P. Syphilis-clinical features, diagnosis, and management. In ABC of Sexually Transmitted Infections, 5th; Adler, M., Cowan, F., French, P., Mitchell, H., Richens, J., Eds.; BMJ Publishing Group: London, UK, 2004; pp. 49–55. [Google Scholar]
- Coquery-Vidrovitch, C. Le Congo au temps des grandes compagnies concessionaires, 1898-1930 (in French); Mouton & Co et EPHE: Paris, France, 1972. [Google Scholar]
- Zimmermann, E. Neu-Kamerun: Reiseerlebnisse und wirtschaftspolitische Untersuchungen (in German); Ernst Siegfried Mittler und Sohn: Berlin, Germany, 1913. [Google Scholar]
- Belgian Congo. Letter from the Comissaire du District du Moyen Congo to the Chef du Service des Terres à Léopoldville, dated from November 12, 1918 (in French). District du Moyen Congo: Leopoldville, Belgian Congo, 1918. Afrika Archief (FO-BZBHO, Brussels), Series GG, Folder GG 16586.
- Samarin, W.J. Protestant missions and the history of Lingala. J. Relig. Af. 1986, 16, 138–163. [Google Scholar]
- Schulte-Varendorff, U. Krieg in Kamerun: Die deutsche Kolonie im Ersten Weltkrieg (in German); Berlin: Christoph Links Verlag: Berlin, Germany, 2011. [Google Scholar]
- Schulte-Varendorff, U. Universität Osnabrück, Osnabrück, Germany. Personal communication, 2012.
- Schler, L. Ambiguous spaces: The struggle over African identities and urban communities in colonial Douala, 1914–45. J. Afr. Hist. 2003, 44, 51–72. [Google Scholar]
- Khabbaz, R.M.; Heneine, W.; George, J.R.; Parekh, B.; Rowe, T.; Woods, T.; Switzer, W.M.; McClure, H.M.; Murphey-Corb, M.; Folks, T.M. Bief report: Infection of a laboratory worker with simian immunodeficiency virus. New Engl. J. Med. 1994, 330, 172–177. [Google Scholar] [CrossRef]
- Korenromp, E.L.; Bakker, R.; de Vlas, S.J.; Gray, R.H.; Wawer, M.J.; Serwadda, D.; Sewankambo, N.K.; Habbema, J.D.F. HIV dynamics and behaviour change as determinants of the impact of sexually transmitted disease treatment on HIV transmission in the context of the Rakai trial. AIDS 2002, 16, 2209–2218. [Google Scholar]
- Orroth, K.K.; Freeman, E.E.; Bakker, R.; Buvé, A.; Glynn, J.R.; Boily, M.-C.; White, R.G.; Habbema, J.D.F.; Hayes, R.J. Understanding the differences between contrasting HIV epidemics in east and west Africa: Results from a simulation model of the Four Cities Study. Sex. Transm. Infect. 2007, 83, i5–i16. [Google Scholar]
- Müller, V. Eötvös Loránd University, Budapest, Hungary. Unpublished computer simulations, 2012.
- Van den Branden, F.; Van Hoof, L. Rapport sur le fonctionnement du Laboratoire de Léopoldville pendant l'année 1923 (in French). Ann. Soc. Belge Méd. Trop. 1924, 4, 231–245. [Google Scholar]
- Schmid, G.P.; Buvé, A.; Mugyenyi, P.; Garnett, G.P.; Hayes, R.J.; Williams, B.G.; Calleja, J.G.; De Cock, K.M.; Whitworth, J.A.; Kapiga, S.H.; et al. Transmission of HIV infection in sub-Saharan Africa and effect of elimination of unsafe injections. Lancet 2004, 363, 482–488. [Google Scholar]
- Njouom, R.; Nerrienet, E.; Dubois, M.; Lachenal, G.; Rousset, D.; Vessière, A.; Ayouba, A.; Pasquier, C.; Pouillot, R. The hepatitis C virus epidemic in Cameroon: Genetic evidence for rapid transmission between 1920 and 1960. Inf. Gen. Evol. 2007, 7, 361–367. [Google Scholar] [CrossRef]
- Frank, C.; Mohamed, M.K.; Strickland, G.T.; Lavanchy, D.; Arthur, R.R.; Magder, L.S.; El Khoby, T.; Abdel-Wahab, Y.; Ohn, E.S.A.; Anwar, W.; et al. The role of parenteral antischistosomal therapy in the spread of hepatitis C virus in Egypt. Lancet 2000, 355, 887–891. [Google Scholar]
- Abdala, N.; Gleghorn, A.A.; Carney, J.M.; Heimer, R. Can HIV-1-contaminated syringes be disinfected? J. Acquir. Immune Defic. Syndr. 2001, 28, 487–494. [Google Scholar]
- Abecasis, A.B.; Vandamme, A.-M.; Lemey, P. Quantifying differences in the tempo of human immunodeficiency virus type 1 subtype evolution. J. Virol. 2009, 83, 12917–12924. [Google Scholar] [CrossRef]
- Buvé, A.; Caraël, M.; Hayes, R.J.; Auvert, B.; Ferry, B.; Robinson, N.J.; Anagonou, S.; Kanhonou, L.; Laourou, M.; Abega, S.; et al. The multicentre study on factors determining the differential spread of HIV in four African cities: Summary and conclusions. AIDS 2001, 15, S127–S131. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sousa, J.D.d.; Alvarez, C.; Vandamme, A.-M.; Müller, V. Enhanced Heterosexual Transmission Hypothesis for the Origin of Pandemic HIV-1. Viruses 2012, 4, 1950-1983. https://doi.org/10.3390/v4101950
Sousa JDd, Alvarez C, Vandamme A-M, Müller V. Enhanced Heterosexual Transmission Hypothesis for the Origin of Pandemic HIV-1. Viruses. 2012; 4(10):1950-1983. https://doi.org/10.3390/v4101950
Chicago/Turabian StyleSousa, João Dinis de, Carolina Alvarez, Anne-Mieke Vandamme, and Viktor Müller. 2012. "Enhanced Heterosexual Transmission Hypothesis for the Origin of Pandemic HIV-1" Viruses 4, no. 10: 1950-1983. https://doi.org/10.3390/v4101950
APA StyleSousa, J. D. d., Alvarez, C., Vandamme, A.-M., & Müller, V. (2012). Enhanced Heterosexual Transmission Hypothesis for the Origin of Pandemic HIV-1. Viruses, 4(10), 1950-1983. https://doi.org/10.3390/v4101950