Nucleoproteins of Negative Strand RNA Viruses; RNA Binding, Oligomerisation and Binding to Polymerase Co-Factor

{kind=link}
{kind=link}
Abstract
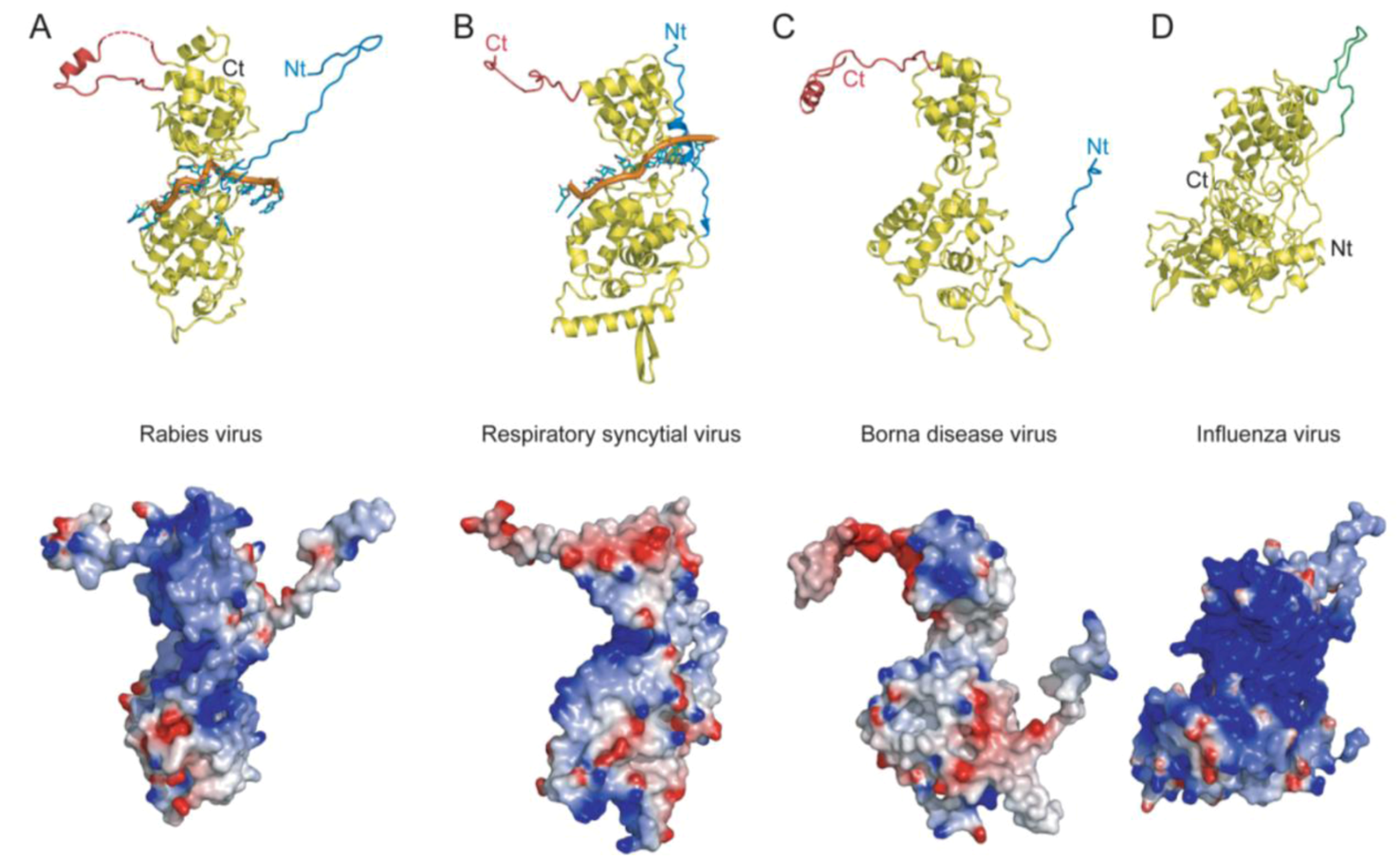
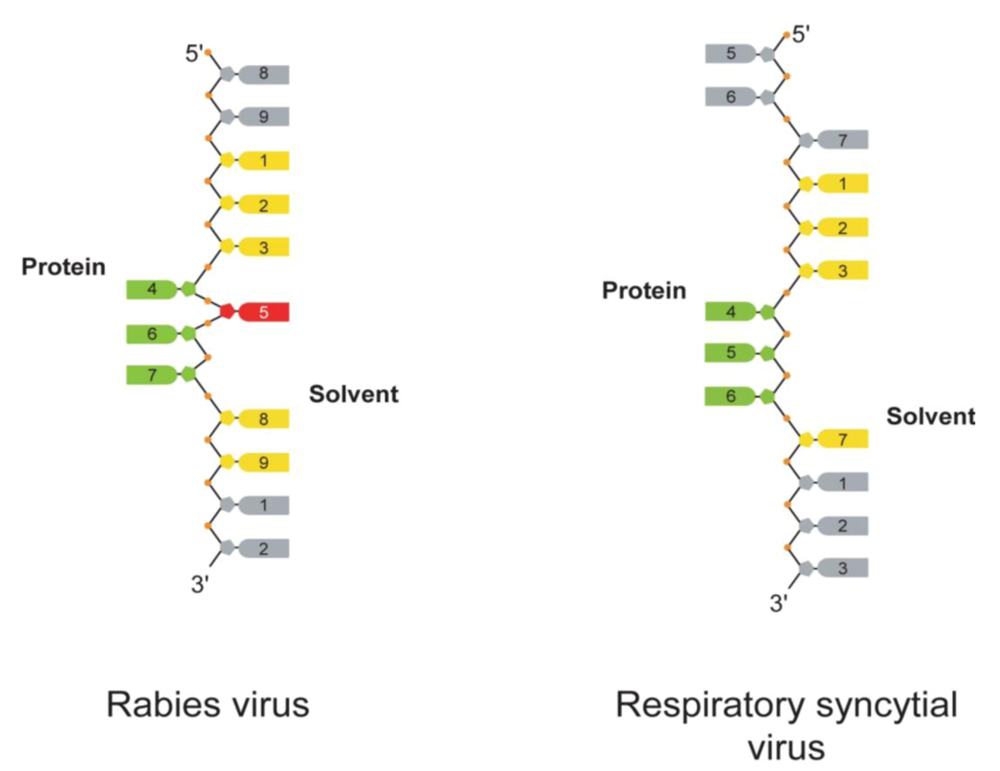
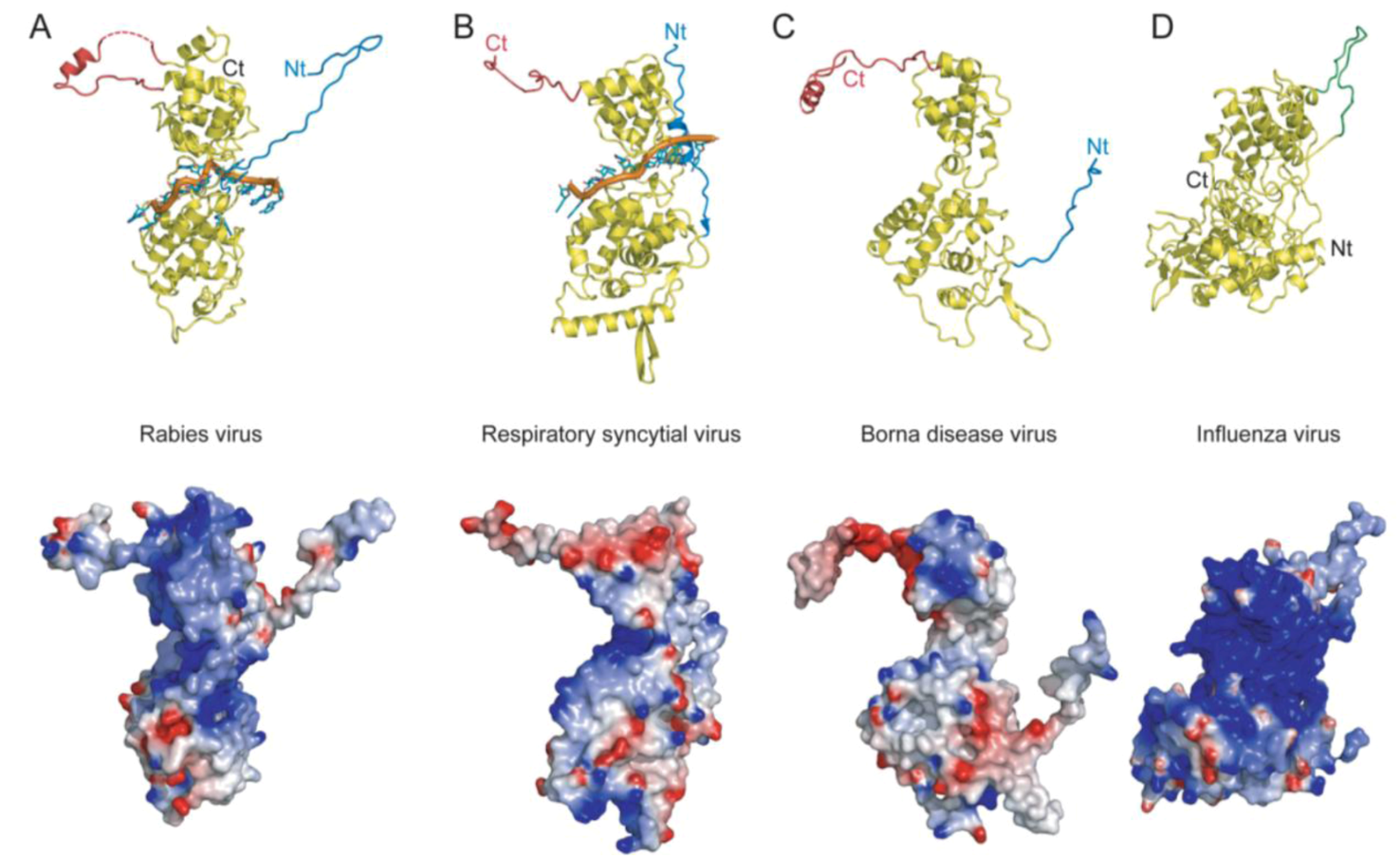
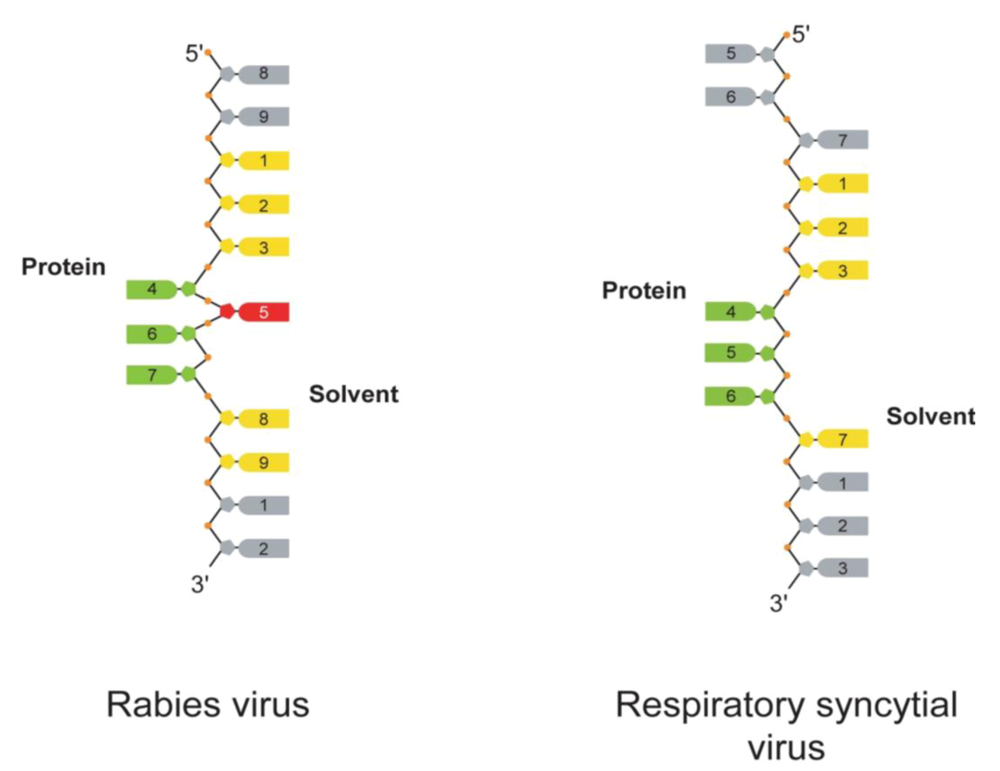
:
Acknowledgments
References
- Tawar, R.G.; Duquerroy, S.; Vonrhein, C.; Varela, P.F.; Damier-Piolle, L.; Castagné, N.; MacLellan, K.; Bedouelle, H.; Bricogne, G.; Bhella, D. Crystal structure of a nucleocapsid-like nucleoprotein-RNA complex of respiratory syncytial virus. Science 2009, 326, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Albertini, A.A.V.; Wernimont, A.K.; Muziol, T.; Ravelli, R.B.G.; Clapier, C.R.; Schoehn, G.; Weissenhorn, W.; Ruigrok, R.W.H. Crystal structure of the rabies virus nucleoprotein-RNA complex. Science 2006, 313, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Green, T.J; Zhang, X.; Wertz, G.W.; Luo, M. Structure of the Vesicular Stomatitis Virus Nucleoprotein-RNA Complex. Science 2006, 313, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, M.G.; Kraus, I.; Dickmanns, A.; Eickmann, M.; Garten, W.; Fickner, R. Crystal structure of the Borna disease virus nucleoprotein. Structure 2003, 11, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Krug, R.M.; Tao, Y.J. The mechanism by which influenza A virus nucleoprotein forms oligomers and binds RNA. Nature 2006, 444, 1078–1082. [Google Scholar] [CrossRef] [PubMed]
- Hock, M.; Kraus, I.; Schoehn, G.; Jamin, M.; Andrei-Selmer, C.; Garten, W.; Weissenhorn, W. RNA induced polymerization of the Borna disease virus nucleoprotein. Virology 2009, 11, 16. [Google Scholar]
- Ng, A.K.; Zhang, H.; Tan, K.; Li, Z.; Liu, J.H.; Chan, P.K.; Li, S.M.; Chan, W.Y.; Au, S.W.; Joachimiak, A.; Walz, T.; Wang, J.H.; Shaw, P.C. Structure of the influenza virus A H5N1 nucleoprotein: implications for RNA binding, oligomerization, and vaccine design. Faseb J. 2008, 22, 3638–3647. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.; Martin-Benito, J.; Zurcher, T.; Valpuesta, J.M.; Carrascosa, J.L.; Ortin, J. Ultrastructural and functional analyses of recombinant influenza virus ribonucleoproteins suggest dimerization of nucleoprotein during virus amplification. J. Virol. 2000, 74, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Longhi, S.; Receveur-Brechot, V.; Karlin, D.; Johansson, K.; Darbon, H.; Bhella, D.; Yeo, R.; Finet, S.; Canard, B. The C-terminal domain of the measles virus nucleoprotein is intrinsically disordered and folds upon binding to the C-terminal moiety of the phosphoprotein. J. Biol. Chem. 2003, 278, 18638–18648. [Google Scholar] [CrossRef] [PubMed]
- Ringkjøbing Jensen, M.; Houben, K.; Lescop, E.; Blanchard, L.; Ruigrok, R.W.H; Blackledge, M. Quantitative Conformational Analysis of the Partially Folded Molecular Recognition Element of Sendai Virus Nucleoprotein from NMR Residual Dipolar Couplings. J.A.C.S. 2008, 130, 8055–8061. [Google Scholar] [CrossRef]
- Kingston, R.L.; Hamel, D.J.; Gay, L.S.; Dahlquist, F.W.; Matthews, B.W. Structural basis for the attachment of a paramyxoviral polymerase to its template. Proc. Natl. Acad. Sci. USA 2004, 101, 8301–8306. [Google Scholar] [CrossRef]
- Houben, K.; Marion, D.; Tarbouriech, N.; Ruigrok, R.W.H.; Blanchard, L. Interaction of the C-terminal domains of Sendai virus N and P proteins: Comparison of polymerase-nucleocapsid interaction within the paramyxovirus family. J. Virol. 2007, 81, 6807–6816. [Google Scholar] [CrossRef] [PubMed]
- Mavrakis, M. Structure and function of the C-terminal domain of the polymerase cofactor of Rabies virus. J. Mol. Biol. 2004, 343, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.A.; Favier, A.; Gerard, F.C.A.; Leyrat, C.; Brutscher, B.; Blondel, D.; Ruigrok, R.W.H.; Blackledge, M.; Jamin, M. Solution structure of the C-terminal N-RNA binding domain of the Vesicular Stomatitis Virus phosphoprotein. J. Mol. Biol. 2008, 382, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Green, T.J.; Luo, M. Structure of the vesicular stomatitis virus nucleocapsid in complex with the nucleocapsid-binding domain of the small polymerase cofactor, P. Proc. Natl. Acad. Sci. USA, 2009, 106, 11721–11726. [Google Scholar] [CrossRef]
- Ribeiro, E.A.; Leyrat, C.; Gérard, F.C.A.; Albertini, A.A.V.; Falk, C.; Ruigrok, R.W.H.; Jamin, M. Binding of rabies virus polymerase cofactor to recombinant circular nucleoprotein-RNA complexes. J. Mol. Biol. 2009, 394, 558–575. [Google Scholar] [CrossRef] [PubMed]
- Calain, P.; Roux, L. The rule of six, a basic feature for efficient replication of Sendai virus defective interfering RNA. J. Virol. 1993, 67, 4822–4830. [Google Scholar] [PubMed]
- Kolakofsky, D.; Pelet, T.; Garcin, D.; Hausmann, S.; Curran, J.; Roux, L. Paramyxovirus RNA synthesis and the requirement for hexamer genome length: the rule of six revisited. J. Virol. 1998, 72, 891–899. [Google Scholar] [PubMed]
- Egelman, E.H.; Wu, S.S. The Sendai virus nucleocapsid exists in at least four different helical states. J. Virol. 1989, 63, 2233–2243. [Google Scholar] [PubMed]
- Iseni, F.; Baudin, F.; Garcin, D.; Marq, J.-B.; Ruigrok, R.W.H.; Kolakofsky, D. Chemical modification of nucleotide bases and mRNA editing depend on hexamer or nucleoprotein phase in Sendai virus nucleocapsids. RNA 2002, 8, 1056–1067. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Ruigrok, R.W.H.; Crépin, T. Nucleoproteins of Negative Strand RNA Viruses; RNA Binding, Oligomerisation and Binding to Polymerase Co-Factor. Viruses 2010, 2, 27-32. https://doi.org/10.3390/v2010027
Ruigrok RWH, Crépin T. Nucleoproteins of Negative Strand RNA Viruses; RNA Binding, Oligomerisation and Binding to Polymerase Co-Factor. Viruses. 2010; 2(1):27-32. https://doi.org/10.3390/v2010027
Chicago/Turabian StyleRuigrok, Rob W. H., and Thibaut Crépin. 2010. "Nucleoproteins of Negative Strand RNA Viruses; RNA Binding, Oligomerisation and Binding to Polymerase Co-Factor" Viruses 2, no. 1: 27-32. https://doi.org/10.3390/v2010027
APA StyleRuigrok, R. W. H., & Crépin, T. (2010). Nucleoproteins of Negative Strand RNA Viruses; RNA Binding, Oligomerisation and Binding to Polymerase Co-Factor. Viruses, 2(1), 27-32. https://doi.org/10.3390/v2010027