
Detection of Porcine Circovirus Type 3 in Free-Ranging Wild Boars and Ticks in Jiangsu Province, China


Abstract
1. Introduction
2. Materials and Methods
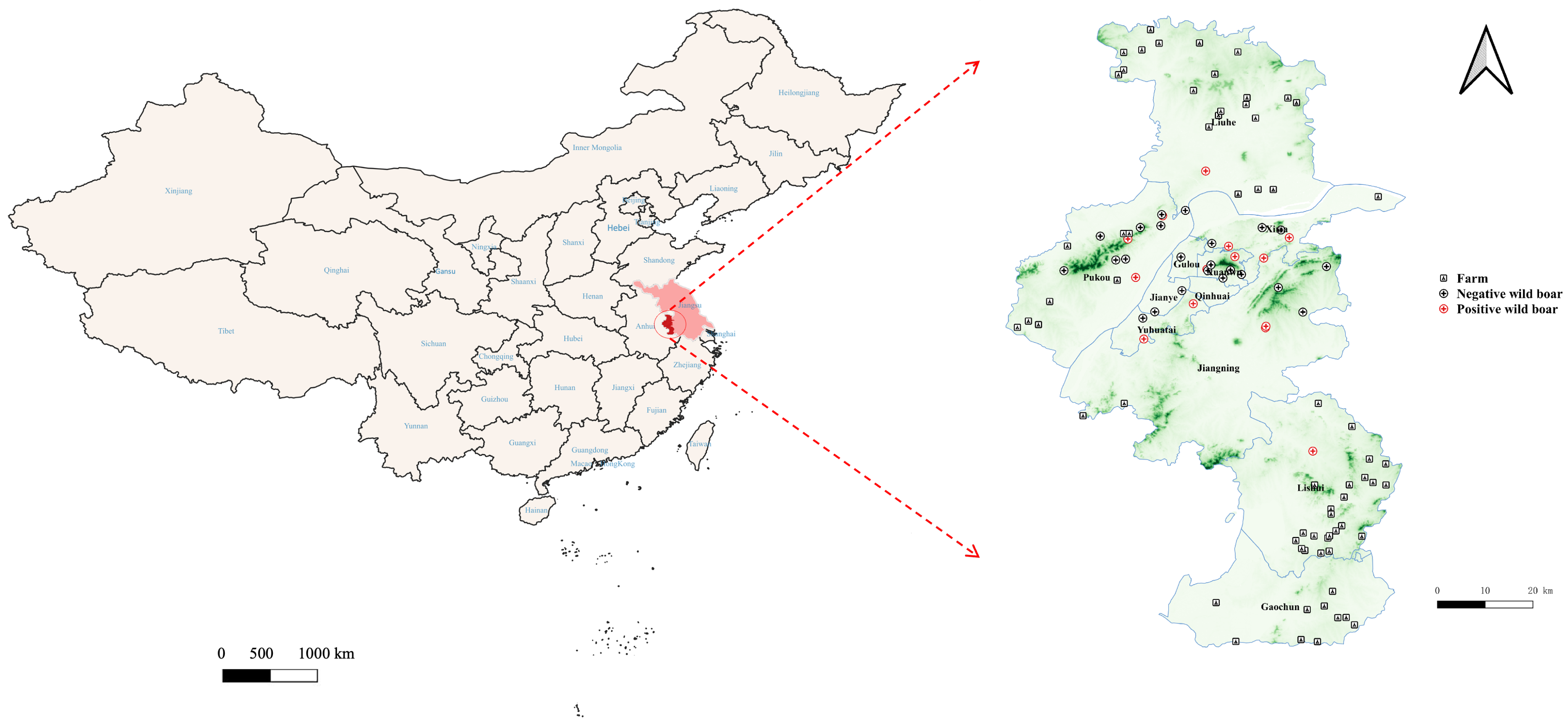
2.1. Sample Collection
2.2. Sample Preparation
2.3. Conventional PCR for PCV3 Detection in Wild Boar Samples
2.4. Detection of PCV3 Antibodies in Wild Boar Sample
2.5. Morphological and Molecular Identification of Ticks
2.6. RT-qPCR for PCV3 Detection in Ticks
2.7. Genome Sequencing and Phylogenetic Analysis
2.8. Statistical Analysis
3. Results
3.1. Conventional PCR Results of PCV3 in Wild Boars
3.2. Antibody Detection Results for PCV3
3.3. Morphological Identification of Ticks and RT-qPCR Results of PCV3 in Ticks
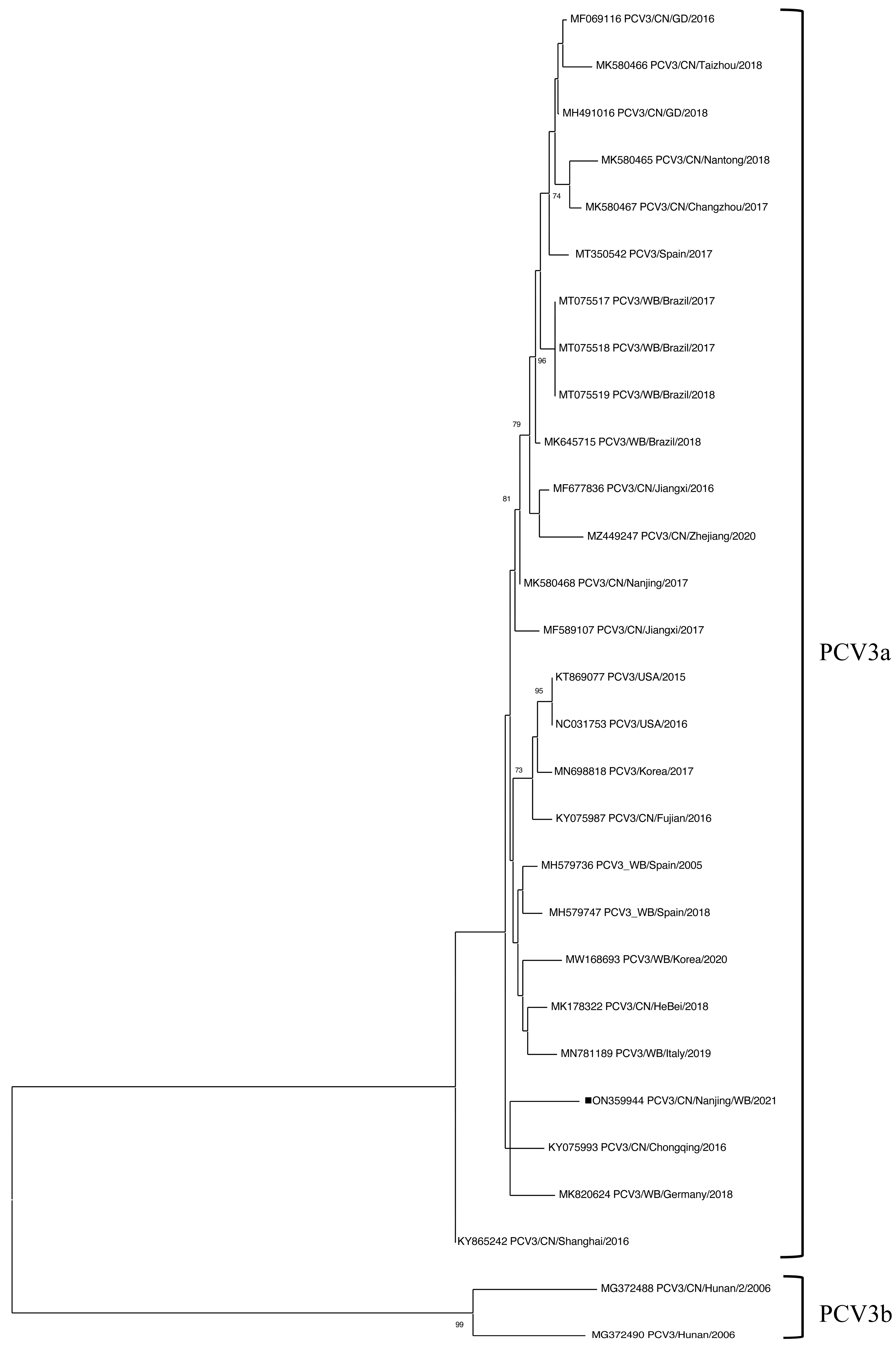
3.4. Sequence Analysis of PCV3 in Wild Boar Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91, e01879-16. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Giannitti, F.; Rossow, S.; Marthaler, D.; Knutson, T.P.; Li, L.; Deng, X.; Resende, T.; Vannucci, F.; Delwart, E. Detection of a novel circovirus PCV3 in pigs with cardiac and multi-systemic inflammation. Virol. J. 2016, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Niu, G.; Liu, X.; Zhang, X.; Zhang, Y.; Ren, L. Recent progress on porcine circovirus type 3. Infect. Genet. Evol. 2019, 73, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, M.; Temeeyasen, G.; Pineyro, P.E. Five years of porcine circovirus 3: What have we learned about the clinical disease, immune pathogenesis, and diagnosis. Virus Res. 2022, 314, 198764. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Tucciarone, C.M.; Drigo, M.; Cecchinato, M.; Martini, M.; Mondin, A.; Menandro, M.L. First report of wild boar susceptibility to Porcine circovirus type 3: High prevalence in the Colli Euganei Regional Park (Italy) in the absence of clinical signs. Transbound. Emerg. Dis. 2018, 65, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Klaumann, F.; Correa-Fiz, F.; Franzo, G.; Sibila, M.; Nunez, J.I.; Segales, J. Current Knowledge on Porcine circovirus 3 (PCV-3): A Novel Virus With a Yet Unknown Impact on the Swine Industry. Front. Vet. Sci. 2018, 5, 315. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Wu, X.; Zhang, L.; Xin, C.; Liu, Y.; Shi, J.; Peng, Z.; Xu, S.; Fu, F.; Yu, J.; et al. The occurrence of porcine circovirus 3 without clinical infection signs in Shandong Province. Transbound. Emerg. Dis. 2017, 64, 1337–1341. [Google Scholar] [CrossRef] [PubMed]
- Bera, B.C.; Choudhary, M.; Anand, T.; Virmani, N.; Sundaram, K.; Choudhary, B.; Tripathi, B.N. Detection and genetic characterization of porcine circovirus 3 (PCV3) in pigs in India. Transbound. Emerg. Dis. 2020, 67, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Sukmak, M.; Thanantong, N.; Poolperm, P.; Boonsoongnern, A.; Ratanavanichrojn, N.; Jirawattanapong, P.; Woonwong, Y.; Soda, N.; Kaminsonsakul, T.; Phuttapatimok, S.; et al. The retrospective identification and molecular epidemiology of porcine circovirus type 3 (PCV3) in swine in Thailand from 2006 to 2017. Transbound. Emerg. Dis. 2019, 66, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Saporiti, V.; Valls, L.; Maldonado, J.; Perez, M.; Correa-Fiz, F.; Segales, J.; Sibila, M. Porcine Circovirus 3 Detection in Aborted Fetuses and Stillborn Piglets from Swine Reproductive Failure Cases. Viruses 2021, 13, 264. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, J.Y.; Jung, J.Y.; Kim, H.Y.; Park, Y.R.; Lee, K.K.; Lyoo, Y.S.; Yeo, S.G.; Park, C.K. Detection and genetic characterization of porcine circovirus 3 from aborted fetuses and pigs with respiratory disease in Korea. J. Vet. Sci. 2018, 19, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Kedkovid, R.; Woonwong, Y.; Arunorat, J.; Sirisereewan, C.; Sangpratum, N.; Lumyai, M.; Kesdangsakonwut, S.; Teankum, K.; Jittimanee, S.; Thanawongnuwech, R. Porcine circovirus type 3 (PCV3) infection in grower pigs from a Thai farm suffering from porcine respiratory disease complex (PRDC). Vet. Microbiol. 2018, 215, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.L.; Zhou, X.; Zhang, H.; Hause, B.M.; Lin, T.; Liu, R.; Chen, Q.L.; Wei, W.K.; Lv, D.H.; Wen, X.H.; et al. Comparative epidemiology of porcine circovirus type 3 in pigs with different clinical presentations. Virol. J. 2017, 14, 222. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Su, M.; Guo, D.; Li, C.; Wei, S.; Feng, L.; Sun, D. Molecular detection and phylogenetic analysis of porcine circovirus type 3 in 21 Provinces of China during 2015–2017. Transbound. Emerg. Dis. 2019, 66, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Yuzhakov, A.G.; Raev, S.A.; Alekseev, K.P.; Grebennikova, T.V.; Verkhovsky, O.A.; Zaberezhny, A.D.; Aliper, T.I. First detection and full genome sequence of porcine circovirus type 3 in Russia. Virus Genes. 2018, 54, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.; Piñeyro, P.; Derscheid, R.; Hause, B.; Byers, E.; Dion, K.; Long, D.; Sievers, C.; Tangen, J.; Williams, T.; et al. PCV3-associated disease in the United States swine herd. Emerg. Microbes Infect. 2019, 8, 684–698. [Google Scholar] [CrossRef] [PubMed]
- Mora-Diaz, J.; Pineyro, P.; Shen, H.; Schwartz, K.; Vannucci, F.; Li, G.; Arruda, B.; Gimenez-Lirola, L. Isolation of PCV3 from Perinatal and Reproductive Cases of PCV3-Associated Disease and In Vivo Characterization of PCV3 Replication in CD/CD Growing Pigs. Viruses 2020, 12, 219. [Google Scholar] [CrossRef] [PubMed]
- Temeeyasen, G.; Lierman, S.; Arruda, B.L.; Main, R.; Vannucci, F.; Gimenez-Lirola, L.G.; Pineyro, P.E. Pathogenicity and immune response against porcine circovirus type 3 infection in caesarean-derived, colostrum-deprived pigs. J. Gen. Virol. 2021, 102. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, D.; Wang, J.; Zhu, S.; She, R.; Ren, X.; Tian, J.; Quan, R.; Hou, L.; Li, Z.; et al. Induction of Porcine Dermatitis and Nephropathy Syndrome in Piglets by Infection with Porcine Circovirus Type 3. J. Virol. 2019, 93, e02045-18. [Google Scholar] [CrossRef] [PubMed]
- Saporiti, V.; Cruz, T.F.; Correa-Fiz, F.; Núñez, J.I.; Sibila, M.; Segalés, J. Similar frequency of Porcine circovirus 3 (PCV-3) detection in serum samples of pigs affected by digestive or respiratory disorders and age-matched clinically healthy pigs. Transbound. Emerg. Dis. 2020, 67, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J.; Lindsay, D.S.; Sriranganathan, N. Wild boars as sources for infectious diseases in livestock and humans. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2697–2707. [Google Scholar] [CrossRef] [PubMed]
- Karniely, S.; Hamed, F.; Gelman, B.; King, R.; Storm, N.; Eyngor, E.; Even Tov, B. First isolation of foot and mouth disease virus from wild boars in the Middle East. Transbound. Emerg. Dis. 2020, 67, 1725–1729. [Google Scholar] [CrossRef] [PubMed]
- Borba, M.R.; Sanches, E.M.; Correa, A.M.; Spanamberg, A.; de Souza Leal, J.; Soares, M.P.; Guillot, J.; Driemeier, D.; Ferreiro, L. Immunohistochemical and ultra-structural detection of Pneumocystis in wild boars (Sus scrofa) co-infected with porcine circovirus type 2 (PCV2) in Southern Brazil. Med. Mycol. 2011, 49, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, A.; Pellegrini, F.; Camero, M.; Catella, C.; Buonavoglia, D.; Fusco, G.; Martella, V.; Lanave, G. Genetic Diversity of Porcine Circovirus Types 2 and 3 in Wild Boar in Italy. Animals 2022, 12, 953. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, M.G.; Serra, F.; Esposito, C.; D’Alessio, N.; Ferrara, G.; Cioffi, B.; Anzalone, A.; Pagnini, U.; De Carlo, E.; Fusco, G.; et al. Prevalence of Infection with Porcine Circovirus Types 2 and 3 in the Wild Boar Population in the Campania Region (Southern Italy). Animals 2021, 11, 3215. [Google Scholar] [CrossRef] [PubMed]
- Klaumann, F.; Dias-Alves, A.; Cabezon, O.; Mentaberre, G.; Castillo-Contreras, R.; Lopez-Bejar, M.; Casas-Diaz, E.; Sibila, M.; Correa-Fiz, F.; Segales, J. Porcine circovirus 3 is highly prevalent in serum and tissues and may persistently infect wild boar (Sus scrofa scrofa). Transbound. Emerg. Dis. 2019, 66, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Prinz, C.; Stillfried, M.; Neubert, L.K.; Denner, J. Detection of PCV3 in German wild boars. Virol. J. 2019, 16, 25. [Google Scholar] [CrossRef] [PubMed]
- Souza, T.; Gava, D.; Schaefer, R.; Leme, R.A.; Porto, G.D.S.; Alfieri, A.A. Porcine Circovirus 3a Field Strains in Free-Living Wild Boars in Parana State, Brazil. Animals 2021, 11, 1634. [Google Scholar] [CrossRef] [PubMed]
- Sato, J.P.H.; Gava, D.; Schaefer, R.; Cantao, M.E.; Ciacci-Zanella, J.R.; de Barcellos, D. Genetic characterization of porcine circovirus type 2 in captive wild boars in southern Brazil. Trop. Anim. Health Prod. 2017, 49, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, F.; Kato, M.; Ito, Y. Porcine circoviruses in wild boars in Nagano Prefecture, Japan. J. Vet. Med. Sci. 2023, 85, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Auer, A.; Schweitzer, L.; Kübber-Heiss, A.; Posautz, A.; Dimmel, K.; Seitz, K.; Beiglböck, C.; Riedel, C.; Rümenapf, T. Porcine Circoviruses and Herpesviruses Are Prevalent in an Austrian Game Population. Pathogens 2022, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Dhandapani, G.; Yoon, S.W.; Noh, J.Y.; Jang, S.S.; Han, S.H.; Jeong, D.G.; Kim, H.K. Detection of Porcine circovirus 3 from captured wild boars in Korea. Vet. Med. Sci. 2021, 7, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Du, H.; Wang, T.; Sun, H.; Peng, P.; Qin, S.; Geng, H.; Zeng, Z.; Liang, W.; Ling, H.; et al. Epizootiological surveillance of porcine circoviruses in free-ranging wild boars in China. Virol. Sin. 2023, 38, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Bowman, A.S.; Nuttall, P.A. Ticks: Biology, Disease and Control; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Franzo, G.; Grassi, L.; Tucciarone, C.M.; Drigo, M.; Martini, M.; Pasotto, D.; Mondin, A.; Menandro, M.L. A wild circulation: High presence of Porcine circovirus 3 in different mammalian wild hosts and ticks. Transbound. Emerg. Dis. 2019, 66, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Nanjing Municipal People’s Government. Nanjing City Profile. Available online: https://www.nanjing.gov.cn/zjnj/csgk/201910/t20191016_1678402.html (accessed on 3 December 2023).
- Nanjing Municipal Bureau of Culture and Tourism. Guide to Handling Matters. Available online: https://wlj.nanjing.gov.cn/zwfw/bszlxz/202201/t20220104_3251346.html (accessed on 3 December 2023).
- Jiang, H. Behind the wild boar in Nanjing: Monitoring, rescue, scientific prevention and control. Available online: http://news.jstv.com/a/20211203/1638516180570.shtml?jsbcApp=1 (accessed on 3 December 2023).
- Franzo, G.; Legnardi, M.; Centelleghe, C.; Tucciarone, C.M.; Cecchinato, M.; Cortey, M.; Segalés, J.; Drigo, M. Development and validation of direct PCR and quantitative PCR assays for the rapid, sensitive, and economical detection of porcine circovirus 3. J. Vet. Diagn. Investig. 2018, 30, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Fux, R.; Söckler, C.; Link, E.K.; Renken, C.; Krejci, R.; Sutter, G.; Ritzmann, M.; Eddicks, M. Full genome characterization of porcine circovirus type 3 isolates reveals the existence of two distinct groups of virus strains. Virol. J. 2018, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, H.H.; El-Molla, A.; Salib, F.A.; Allam, N.A.; Ghazy, A.A.; Abdel-Shafy, S. Morphological and molecular identification of the brown dog tick Rhipicephalus sanguineus and the camel tick Hyalomma dromedarii (Acari: Ixodidae) vectors of Rickettsioses in Egypt. Vet. World 2016, 9, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wu, S.; Zhang, Y.; Zhang, T.; Feng, C.; Jia, G.; Lin, X. Development of a DNA barcoding system for the Ixodida (Acari: Ixodida). Mitochondrial DNA 2014, 25, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Delwart, E.; Fux, R.; Hause, B.; Su, S.; Zhou, J.; Segales, J. Genotyping Porcine Circovirus 3 (PCV-3) Nowadays: Does It Make Sense? Viruses 2020, 12, 265. [Google Scholar] [CrossRef] [PubMed]
- Varela, A.P.M.; Loiko, M.R.; Andrade, J.d.S.; Tochetto, C.; Cibulski, S.P.; Lima, D.A.; Weber, M.N.; Roehe, P.M.; Mayer, F.Q. Complete genome characterization of porcine circovirus 3 recovered from wild boars in Southern Brazil. Transbound. Emerg. Dis. 2020, 68, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chen, Z.; Li, Y.; Ding, Z.; Zeng, Q.; Wan, T.; Wu, H. Detection of Porcine Circovirus 1/2/3 and Genetic Analysis of Porcine Circovirus 2 in Wild Boar from Jiangxi Province of China. Animals 2022, 12, 2021. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Contreras, R.; Carvalho, J.; Serrano, E.; Mentaberre, G.; Fernández-Aguilar, X.; Colom, A.; González-Crespo, C.; Lavín, S.; López-Olvera, J.R. Urban wild boars prefer fragmented areas with food resources near natural corridors. Sci. Total Environ. 2018, 615, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Fons, F. A Review of the Current Status of Relevant Zoonotic Pathogens in Wild Swine (Sus scrofa) Populations: Changes Modulating the Risk of Transmission to Humans. Transbound. Emerg. Dis. 2017, 64, 68–88. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Cao, G.; Quan, R.-C. Population dynamics and space use of wild boar in a tropical forest, Southwest China. Glob. Ecol. Conserv. 2017, 11, 115–124. [Google Scholar] [CrossRef]
- Shi, R.; Hou, L.; Liu, J. Host immune response to infection with porcine circoviruses. Anim. Dis. 2021, 1, 23. [Google Scholar] [CrossRef]
- Fafi-Kremer, S.; Brengel-Pesce, K.; Barguès, G.; Bourgeat, M.-J.; Genoulaz, O.; Seigneurin, J.-M.; Morand, P. Assessment of automated DNA extraction coupled with real-time PCR for measuring Epstein–Barr virus load in whole blood, peripheral mononuclear cells and plasma. J. Clin. Virol. 2004, 30, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, G.; Duan, W.T.; Sun, M.X.; Wang, M.H.; Wang, S.H.; Cai, X.H.; Tu, Y.B. Self-assembly into virus-like particles of the recombinant capsid protein of porcine circovirus type 3 and its application on antibodies detection. AMB Express 2020, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, W.M.A.; Moustafa, M.A.M.; Thu, M.J.; Kakisaka, K.; Chatanga, E.; Ogata, S.; Hayashi, N.; Taya, Y.; Ohari, Y.; Naguib, D.; et al. Comparative mitogenomics elucidates the population genetic structure of Amblyomma testudinarium in Japan and a closely related Amblyomma species in Myanmar. Evol. Appl. 2022, 15, 1062–1078. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.B.; Kang, J.G.; Kim, H.C.; Chong, S.T.; Lee, I.Y.; Shin, N.S.; Chae, J.S. Identification of Tick Species Collected from Wild Boars and Habitats of Wild Boars and Domestic Pigs in the Republic of Korea. Korean J. Parasitol. 2017, 55, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Nakao, R.; Shinjo, K.; Sakiyama, T.; Ogata, S.; Kusakisako, K.; Kinoshita, G.; Naguib, D.; Chatanga, E.; Mohamed, W.M.A.; Moustafa, M.A.M.; et al. Amblyomma testudinarium infestation on a brown bear (Ursus arctos yesoensis) captured in Hokkaido, a northern island of Japan. Parasitol. Int. 2021, 80, 102209. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Beppu, T.; Inokuma, H.; Okuda, M.; Onishi, T. Ixodid tick species recovered from domestic dogs in Japan. Med. Vet. Entomol. 2003, 17, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.H.; Kim, H.C.; Yun, S.M.; Lim, J.W.; Kim, J.H.; Chong, S.T.; Kim, D.H.; Kim, H.T.; Kim, H.; Klein, T.A.; et al. Detection of SFTS Virus in Ixodes nipponensis and Amblyomma testudinarium (Ixodida: Ixodidae) Collected From Reptiles in the Republic of Korea. J. Med. Entomol. 2016, 53, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Sumrandee, C.; Hirunkanokpun, S.; Doornbos, K.; Kitthawee, S.; Baimai, V.; Grubhoffer, L.; Trinachartvanit, W.; Ahantarig, A. Molecular detection of Rickettsia species in Amblyomma ticks collected from snakes in Thailand. Ticks Tick Borne Dis. 2014, 5, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Ling, F.; Chai, C.; Lu, Y.; Yu, X.; Lin, J.; Sun, J.; Chang, Y.; Ye, X.; Gu, S.; et al. Prevalence of Borrelia burgdorferi sensu lato in ticks from eastern China. Am. J. Trop. Med. Hyg. 2015, 92, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | PCR Product | Assay | Reference |
|---|---|---|---|---|
| PCV3-74F | CACCGTGTGAGTGGATATAC | 1072 bp | Conventional PCR | [40] |
| PCV3-1144R | CACCCCAACGCAATAATTGTA | |||
| PCV3-1137F | TTGGGGTGGGGGTATTTATT | 425 bp | ||
| PCV3-1561R | ACACAGCCGTTACTTCAC | |||
| PCV3-1427F | AGTGCTCCCCATTGAACG | 1007 bp | ||
| PCV3-433R | CGACCAAATCCGGGTAAGC | |||
| PCV3-qF | TGACGGAGACGTCGGGAAAT | 113 bp | RT-qPCR | [39] |
| PCV3-qR | CGGTTTACCCAACCCCATCA | |||
| PCV3-probe | FAM-GGGCGGGGTTTGCGTGATTT-BHQ1 |
| Wild Boar ID | PCV3 PCR Results | PCV3 Antibody Results | |||||
|---|---|---|---|---|---|---|---|
| Blood | Serum | Feces | Tissues | Oral Fluid | Ticks | ||
| 1 | NC * | NC | − | NC | − | NC | NC |
| 2 | + ** | − | + | NC | NC | NC | + |
| 3 | − | − | − | NC | NC | NC | + |
| 4 | + | − | − | NC | NC | NC | − |
| 5 | − | − | − | NC | NC | 0/5 | + |
| 6 | + | − | NC | − | NC | 0/11 | − |
| 7 | + | − | NC | − | NC | 0/3 | − |
| 8 | + | + | NC | + | NC | 2/2 | − |
| 9 | + | − | − | NC | NC | 3/18 | + |
| 10 | + | − | NC | NC | NC | NC | + |
| 11 | NC | NC | + | NC | NC | NC | − |
| 12 | + | − | − | NC | NC | NC | + |
| 13 | − | − | NC | − | NC | NC | + |
| 14 | − | − | NC | − | NC | NC | + |
| 15 | + | − | NC | NC | NC | NC | + |
| 16 | − | − | NC | NC | NC | NC | + |
| 17 | − | − | NC | NC | NC | NC | − |
| 18 | − | − | NC | − | NC | NC | + |
| 19 | + | − | NC | NC | NC | 0/4 | + |
| 20 | + | − | NC | − | NC | NC | − |
| 21 | − | + | NC | − | NC | NC | + |
| 22 | + | − | NC | NC | NC | 0/7 | − |
| 23 | − | − | NC | NC | NC | NC | + |
| 24 | − | − | NC | NC | NC | NC | + |
| 25 | − | − | NC | − | NC | NC | − |
| 26 | − | − | NC | NC | NC | NC | − |
| 27 | − | − | NC | NC | NC | NC | − |
| 28 | − | − | NC | NC | NC | NC | + |
| 29 | − | − | NC | NC | NC | NC | − |
| 30 | − | − | NC | NC | NC | NC | − |
| 31 | − | − | NC | NC | NC | NC | − |
| 32 | − | − | NC | − | NC | NC | − |
| 33 | − | − | NC | − | NC | NC | − |
| 34 | − | − | NC | − | NC | 0/6 | + |
| 35 | − | − | NC | NC | NC | NC | − |
| 36 | − | − | NC | NC | NC | NC | − |
| 37 | − | − | NC | NC | NC | NC | − |
| 38 | − | − | NC | NC | NC | NC | − |
| 39 | − | − | NC | NC | NC | NC | − |
| 40 | − | − | NC | NC | NC | NC | − |
| 41 | − | − | NC | + | NC | NC | − |
| 42 | − | − | NC | NC | NC | NC | − |
| 43 | − | − | NC | NC | NC | NC | − |
| 44 | − | − | NC | NC | NC | NC | − |
| 45 | − | − | NC | NC | NC | 1/11 | + |
| 46 | NC | NC | NC | − | NC | NC | NC |
| 47 | NC | NC | NC | + | NC | 0/16 | NC |
| Total | 12/43 | 2/43 | 2/8 | 3/15 | 0/1 | 6/83 | 18/43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, F.; Li, M.; Wang, Y.; Cheng, W.; Li, M.; Deng, C.; Wang, X.; Yang, Z. Detection of Porcine Circovirus Type 3 in Free-Ranging Wild Boars and Ticks in Jiangsu Province, China. Viruses 2025, 17, 1049. https://doi.org/10.3390/v17081049
Sun F, Li M, Wang Y, Cheng W, Li M, Deng C, Wang X, Yang Z. Detection of Porcine Circovirus Type 3 in Free-Ranging Wild Boars and Ticks in Jiangsu Province, China. Viruses. 2025; 17(8):1049. https://doi.org/10.3390/v17081049
Chicago/Turabian StyleSun, Fanqi, Meng Li, Yi Wang, Wangkun Cheng, Meirong Li, Changlin Deng, Xianwei Wang, and Zhen Yang. 2025. "Detection of Porcine Circovirus Type 3 in Free-Ranging Wild Boars and Ticks in Jiangsu Province, China" Viruses 17, no. 8: 1049. https://doi.org/10.3390/v17081049
APA StyleSun, F., Li, M., Wang, Y., Cheng, W., Li, M., Deng, C., Wang, X., & Yang, Z. (2025). Detection of Porcine Circovirus Type 3 in Free-Ranging Wild Boars and Ticks in Jiangsu Province, China. Viruses, 17(8), 1049. https://doi.org/10.3390/v17081049