
Plant Lectin, MoMo30, Pressures HIV-1 to Select for Variants with Deleted N-Linked Glycosylation Sites

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. HIV-1 Evolution Experiment with MoMo30 Treatment
2.2. Isolation of Virus from Supernatant
2.3. Total RNA Sequencing
2.4. p24 ELISA
2.5. Single-Round Infection Assay
2.6. Statistical Analysis
3. Results
3.1. RNA Sequencing
3.2. Analysis of Potential N-Linked Glycosylation Site Mutations
3.3. Analysis of gp120 Variable Loop Mutationsa
3.4. Analysis of Mutations with gp41 Functional Domains
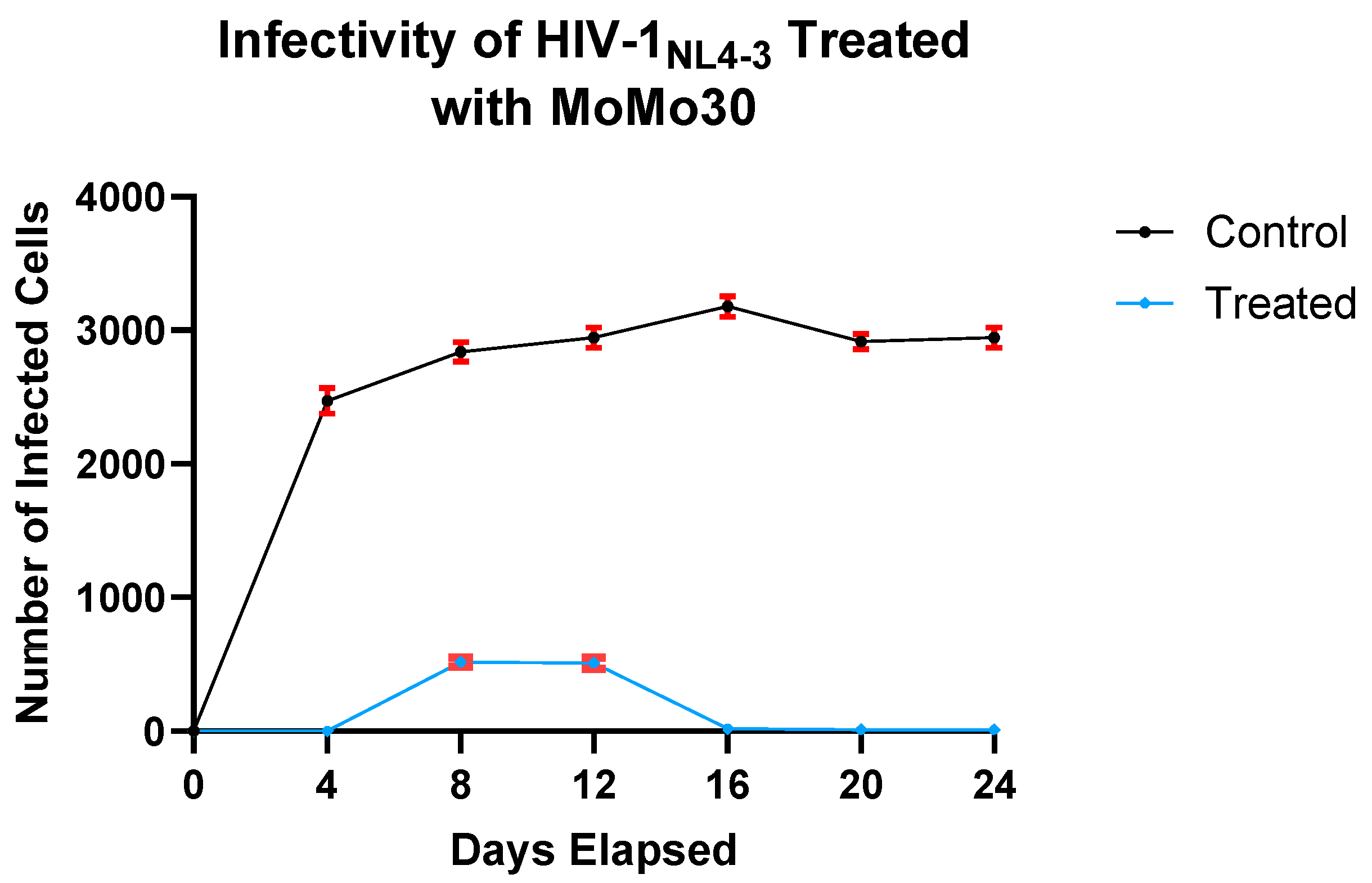
3.5. p24 LEISA: Effects of MoMo30 on HIV Replication over Time
3.6. Single-Round Infection Assay
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swinkels, H.M.; Nguyen, A.D.; Gulick, P.G. HIV and AIDS. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Masenga, S.K.; Mweene, B.C.; Luwaya, E.; Muchaili, L.; Chona, M.; Kirabo, A. HIV–Host Cell Interactions. Cells 2023, 12, 1351. [Google Scholar] [CrossRef] [PubMed]
- Foka, F.E.T.; Mufhandu, H.T. Current ARTs, Virologic Failure, and Implications for AIDS Management: A Systematic Review. Viruses 2023, 15, 1732. [Google Scholar] [CrossRef] [PubMed]
- Sever, B.; Otsuka, M.; Fujita, M.; Ciftci, H. A Review of FDA-Approved Anti-HIV-1 Drugs, Anti-Gag Compounds, and Potential Strategies for HIV-1 Eradication. Int. J. Mol. Sci. 2024, 25, 3659. [Google Scholar] [CrossRef]
- Kim, J.; Vasan, S.; Kim, J.H.; Ake, J.A. Current approaches to HIV vaccine development: A narrative review. J. Int. AIDS Soc. 2021, 24 (Suppl. S7), e25793. [Google Scholar] [CrossRef] [PubMed]
- Govindan, R.; Stephenson, K.E. HIV Vaccine Development at a Crossroads: New B and T Cell Approaches. Vaccines 2024, 12, 1043. [Google Scholar] [CrossRef]
- Chen, B. Molecular Mechanism of HIV-1 Entry. Trends Microbiol. 2019, 27, 878–891. [Google Scholar] [CrossRef]
- Xiao, T.; Cai, Y.; Chen, B. HIV-1 Entry and Membrane Fusion Inhibitors. Viruses 2021, 13, 735. [Google Scholar] [CrossRef]
- Zhao, C.; Li, H.; Swartz, T.H.; Chen, B.K.; Wu, L.; Prasad, V.R. The HIV Env Glycoprotein Conformational States on Cells and Viruses. mBio 2022, 13, e01825-21. [Google Scholar] [CrossRef]
- Cao, L.; Diedrich, J.K.; Kulp, D.W.; Pauthner, M.; He, L.; Park, S.-K.R.; Sok, D.; Su, C.Y.; Delahunty, C.M.; Menis, S.; et al. Global site-specific N-glycosylation analysis of HIV envelope glycoprotein. Nat. Commun. 2017, 8, 14954. [Google Scholar] [CrossRef]
- Lambert, G.S.; Upadhyay, C. HIV-1 Envelope Glycosylation and the Signal Peptide. Vaccines 2021, 9, 176. [Google Scholar] [CrossRef]
- Upadhyay, C.; Rao, P.; Behzadi, M.A.; Feyznezhad, R.; Lambert, G.S.; Kumar, R.; Kumar, M.; Yang, W.; Jiang, X.; Luo, C.C.; et al. Signal peptide exchange alters HIV-1 envelope antigenicity and immunogenicity. Front. Immunol. 2024, 15, 1476924. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Berndsen, Z.T.; Hengartner, N.W.; Korber, B.T.; Ward, A.B.; Gnanakaran, S. Quantification of the Resilience and Vulnerability of HIV-1 Native Glycan Shield at Atomistic Detail. iScience 2020, 23, 101836. [Google Scholar] [CrossRef] [PubMed]
- Thavarajah, J.J.; Hønge, B.L.; Wejse, C.M. The Use of Broadly Neutralizing Antibodies (bNAbs) in HIV-1 Treatment and Prevention. Viruses 2024, 16, 911. [Google Scholar] [CrossRef]
- Kong, R.; Xu, K.; Zhou, T.; Acharya, P.; Lemmin, T.; Liu, K.; Ozorowski, G.; Soto, C.; Taft, J.D.; Bailer, R.T.; et al. Fusion peptide of HIV-1 as a site of vulnerability to neutralizing antibody. Science 2016, 352, 828–833. [Google Scholar] [CrossRef]
- Piai, A.; Fu, Q.; Cai, Y.; Ghantous, F.; Xiao, T.; Shaik, M.M.; Peng, H.; Rits-Volloch, S.; Chen, W.; Seaman, M.S.; et al. Structural basis of transmembrane coupling of the HIV-1 envelope glycoprotein. Nat. Commun. 2020, 11, 2317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, Y.; Hu, H.; Zhang, S.; Wang, P.; Chong, H.; He, J.; Wang, X.; He, Y. Structural Insights into the Mechanisms of Action of Short-Peptide HIV-1 Fusion Inhibitors Targeting the Gp41 Pocket. Front. Cell. Infect. Microbiol. 2018, 8, 51. [Google Scholar] [CrossRef]
- Zhu, Y.; Su, S.; Qin, L.; Wang, Q.; Shi, L.; Ma, Z.; Tang, J.; Jiang, S.; Lu, L.; Ye, S.; et al. Rational improvement of gp41-targeting HIV-1 fusion inhibitors: An innovatively designed Ile-Asp-Leu tail with alternative conformations. Sci. Rep. 2016, 6, 31983. [Google Scholar] [CrossRef]
- Barre, A.; Van Damme, E.J.M.; Klonjkowski, B.; Simplicien, M.; Sudor, J.; Benoist, H.; Rougé, P. Legume Lectins with Different Specificities as Potential Glycan Probes for Pathogenic Enveloped Viruses. Cells 2022, 11, 339. [Google Scholar] [CrossRef]
- Cummings, R.D.; Etzler, M.; Hahn, M.G.; Darvill, A.; Godula, K.; Woods, R.J.; Mahal, L.K. Glycan-Recognizing Probes as Tools. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Mohnen, D., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2022; pp. 645–662. [Google Scholar]
- Francesconi, O.; Donnici, L.; Fragai, M.; Pesce, E.; Bombaci, M.; Fasciani, A.; Manganaro, L.; Conti, M.; Grifantini, R.; De Francesco, R.; et al. Synthetic carbohydrate-binding agents neutralize SARS-CoV-2 by inhibiting binding of the spike protein to ACE2. iScience 2022, 25, 104239. [Google Scholar] [CrossRef]
- Nabi-Afjadi, M.; Heydari, M.; Zalpoor, H.; Arman, I.; Sadoughi, A.; Sahami, P.; Aghazadeh, S. Lectins and lectibodies: Potential promising antiviral agents. Cell. Mol. Biol. Lett. 2022, 27, 37. [Google Scholar] [CrossRef]
- Balzarini, J.; Van Laethem, K.; Hatse, S.; Froeyen, M.; Peumans, W.; Van Damme, E.; Schols, D. Carbohydrate-binding Agents Cause Deletions of Highly Conserved Glycosylation Sites in HIV GP120: A NEW THERAPEUTIC CONCEPT TO HIT THE ACHILLES HEEL OF HIV. J. Biol. Chem. 2005, 280, 41005–41014. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Van Laethem, K.; Hatse, S.; Froeyen, M.; Van Damme, E.; Bolmstedt, A.; Peumans, W.; De Clercq, E.; Schols, D. Marked Depletion of Glycosylation Sites in HIV-1 gp120 under Selection Pressure by the Mannose-Specific Plant Lectins of Hippeastrum Hybrid and Galanthus nivalis. Mol. Pharmacol. 2005, 67, 1556–1565. [Google Scholar] [CrossRef]
- Vanderlinden, E.; Van Winkel, N.; Naesens, L.; Van Damme, E.J.M.; Persoons, L.; Schols, D. In Vitro Characterization of the Carbohydrate-Binding Agents HHA, GNA, and UDA as Inhibitors of Influenza A and B Virus Replication. Antimicrob. Agents Chemother. 2021, 65, e01732-20. [Google Scholar] [CrossRef]
- Khan, M.; Diop, A.; Gbodossou, E.; Xiao, P.; Coleman, M.; De Barros, K.; Duong, H.; Bond, V.C.; Floyd, V.; Kondwani, K.; et al. Anti-human immunodeficiency virus-1 activity of MoMo30 protein isolated from the traditional African medicinal plant Momordica balsamina. Virol. J. 2023, 20, 50. [Google Scholar] [CrossRef]
- Coleman, M.I.; Khan, M.; Gbodossou, E.; Diop, A.; DeBarros, K.; Duong, H.; Bond, V.C.; Floyd, V.; Kondwani, K.; Rice, V.M.; et al. Identification of a Novel Anti-HIV-1 Protein from Momordica balsamina Leaf Extract. Int. J. Environ. Res. Public Health 2022, 19, 15227. [Google Scholar] [CrossRef] [PubMed]
- DeBarros, K.; Khan, M.; Coleman, M.; Bond, V.C.; Floyd, V.; Gbodossou, E.; Diop, A.; Krumpe, L.R.H.; O’keefe, B.R.; Powell, M.D. MoMo30 Binds to SARS-CoV-2 Spike Variants and Blocks Infection by SARS-CoV-2 Pseudovirus. Viruses 2024, 16, 1433. [Google Scholar] [CrossRef]
- Behrens, A.-J.; Harvey, D.J.; Milne, E.; Cupo, A.; Kumar, A.; Zitzmann, N.; Struwe, W.B.; Moore, J.P.; Crispin, M.; Ross, S.R. Molecular Architecture of the Cleavage-Dependent Mannose Patch on a Soluble HIV-1 Envelope Glycoprotein Trimer. J. Virol. 2017, 91, 01894-16. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.K.; Harvey, D.J.; Bonomelli, C.; Crispin, M.; Doores, K.J.; Kirchhoff, F. Cell- and Protein-Directed Glycosylation of Native Cleaved HIV-1 Envelope. J. Virol. 2015, 89, 8932–8944. [Google Scholar] [CrossRef]
- Beretta, M.; Migraine, J.; Moreau, A.; Essat, A.; Goujard, C.; Chaix, M.-L.; Drouin, A.; Bouvin-Pley, M.; Meyer, L.; Barin, F.; et al. Common evolutionary features of the envelope glycoprotein of HIV-1 in patients belonging to a transmission chain. Sci. Rep. 2020, 10, 16744. [Google Scholar] [CrossRef]
- Zhang, Z.; Anang, S.; Nguyen, H.T.; Fritschi, C.; Smith, A.B.; Sodroski, J.G. Membrane HIV-1 envelope glycoproteins stabilized more strongly in a pretriggered conformation than natural virus Envs. iScience 2024, 27, 110141. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, S.; Wang, K.; Ding, H.; Zhang, Z.; Anang, S.; Nguyen, H.T.; Kappes, J.C.; Sodroski, J.; Mao, Y. The membrane-proximal external region of human immunodeficiency virus (HIV-1) envelope glycoprotein trimers in A18-lipid nanodiscs. Commun. Biol. 2025, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Datta, R.; Varadarajan, R.; Silvestri, G. Probing the Structure of the HIV-1 Envelope Trimer Using Aspartate Scanning Mutagenesis. J. Virol. 2020, 94, e01426-20. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-C.; Tang, M.; Zhang, M.-Y.; Majeed, S.; Montabana, E.; Stanfield, R.L.; Dimitrov, D.S.; Korber, B.; Sodroski, J.; Wilson, I.A.; et al. Structure of a V3-containing HIV-1 gp120 core. Science 2005, 310, 1025–1028. [Google Scholar] [CrossRef]
- Schapiro, H.M.; Khasnis, M.D.; Ahn, K.; Karagiaridi, A.; Hayden, S.; Cilento, M.E.; Root, M.J.; Campbell, E.M. Regulation of epitope exposure in the gp41 membrane-proximal external region through interactions at the apex of HIV-1 Env. PLOS Pathog. 2022, 18, e1010531. [Google Scholar] [CrossRef]
- Sethi, A.; Tian, J.; Derdeyn, C.A.; Korber, B.; Gnanakaran, S.; Shakhnovich, E.I. A mechanistic understanding of allosteric immune escape pathways in the HIV-1 envelope glycoprotein. PLOS Comput. Biol. 2013, 9, e1003046. [Google Scholar] [CrossRef]
- Yuan, T.; Li, J.; Zhang, M.-Y.; Vartanian, J.-P. HIV-1 envelope glycoprotein variable loops are indispensable for envelope structural integrity and virus entry. PLoS ONE 2013, 8, e69789. [Google Scholar] [CrossRef] [PubMed]
- De Luca, A. The impact of resistance on viral fitness and its clinical implications. In Antiretroviral Resistance in Clinical Practice; Geretti, A.M., Ed.; Mediscript: London, UK, 2006. [Google Scholar]
- Schorcht, A.; Cottrell, C.A.; Pugach, P.; Ringe, R.P.; Han, A.X.; Allen, J.D.; Kerkhof, T.L.G.M.v.D.; Seabright, G.E.; Schermer, E.E.; Ketas, T.J.; et al. The Glycan Hole Area of HIV-1 Envelope Trimers Contributes Prominently to the Induction of Autologous Neutralization. J. Virol. 2022, 96, e01552-21. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Zuo, T.; Hora, B.; Cai, F.; Ding, H.; Kappes, J.; Ochsenbauer, C.; Kong, W.; Yu, X.; et al. Accumulated mutations by 6 months of infection collectively render transmitted/founder HIV-1 significantly less fit. J. Infect. 2020, 80, 210–218. [Google Scholar] [CrossRef]
- Balzarini, J. Targeting the glycans of glycoproteins: A novel paradigm for antiviral therapy. Nat. Rev. Microbiol. 2007, 5, 583–597. [Google Scholar] [CrossRef]
- Liu, Y.-M.; Mahmud, S.A.; Chen, X.; Chen, T.-H.; Liao, K.-S.; Lo, J.M.; Wu, Y.-M.; Ho, M.-C.; Wu, C.-Y.; Wong, C.-H.; et al. A Carbohydrate-Binding Protein from the Edible Lablab Beans Effectively Blocks the Infections of Influenza Viruses and SARS-CoV-2. Cell Rep. 2020, 32, 108016. [Google Scholar] [CrossRef]
- Santisteban Celis, I.C.; Matoba, N. Lectibodies as antivirals. Antivir. Res. 2024, 227, 105901. [Google Scholar] [CrossRef] [PubMed]
- Tampere, M.; Pettke, A.; Salata, C.; Wallner, O.; Koolmeister, T.; Cazares-Körner, A.; Visnes, T.; Hesselman, M.C.; Kunold, E.; Wiita, E.; et al. Novel Broad-Spectrum Antiviral Inhibitors Targeting Host Factors Essential for Replication of Pathogenic RNA Viruses. Viruses 2020, 12, 1423. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Gutierrez, S.; Buckley, J.; Battaglia, G. The Role of Host Cell Glycans on Virus Infectivity: The SARS-CoV-2 Case. Adv. Sci. 2022, 10, e2201853. [Google Scholar] [CrossRef]
- Dugan, A.E.; Peiffer, A.L.; Kiessling, L.L. Advances in glycoscience to understand viral infection and colonization. Nat. Methods 2022, 19, 384–387. [Google Scholar] [CrossRef]
- Li, Y.; Liu, D.; Wang, Y.; Su, W.; Liu, G.; Dong, W. The Importance of Glycans of Viral and Host Proteins in Enveloped Virus Infection. Front. Immunol. 2021, 12, 638573. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.N.; Jahan, R.; Nissapatorn, V.; Wilairatana, P.; Rahmatullah, M. Plant lectins as prospective antiviral biomolecules in the search for COVID-19 eradication strategies. Biomed. Pharmacother. 2022, 146, 112507. [Google Scholar] [CrossRef]
- Swanson, M.D.; Boudreaux, D.M.; Salmon, L.; Chugh, J.; Winter, H.C.; Meagher, J.L.; André, S.; Murphy, P.V.; Oscarson, S.; Roy, R.; et al. Engineering a therapeutic lectin by uncoupling mitogenicity from antiviral activity. Cell 2015, 163, 746–758. [Google Scholar] [CrossRef]
- Burton, D.R.; Hangartner, L. Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design. Annu. Rev. Immunol. 2016, 34, 635–659. [Google Scholar] [CrossRef]
- Moore, P.L.; Williamson, C.; Morris, L. Virological features associated with the development of broadly neutralizing antibodies to HIV-1. Trends Microbiol. 2015, 23, 204–211. [Google Scholar] [CrossRef]
- Timofeeva, A.; Sedykh, S.; Nevinsky, G. Post-Immune Antibodies in HIV-1 Infection in the Context of Vaccine Development: A Variety of Biological Functions and Catalytic Activities. Vaccines 2022, 10, 384. [Google Scholar] [CrossRef]
- Tolbert, W.D.; Sherburn, R.T.; Van, V.; Pazgier, M. Structural Basis for Epitopes in the gp120 Cluster A Region that Invokes Potent Effector Cell Activity. Viruses 2019, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Voss, J.E.; Andrabi, R.; McCoy, L.E.; de Val, N.; Fuller, R.P.; Messmer, T.; Su, C.-Y.; Sok, D.; Khan, S.N.; Garces, F.; et al. Elicitation of Neutralizing Antibodies Targeting the V2 Apex of the HIV Envelope Trimer in a Wild-Type Animal Model. Cell Rep. 2017, 21, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Wibmer, C.K.; Moore, P.L.; Morris, L. HIV broadly neutralizing antibody targets. Curr. Opin. HIV AIDS 2015, 10, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Walker, S.; Wise, M.C.; Chokkalingam, N.; Purwar, M.; Moore, A.; Tello-Ruiz, E.; Wu, Y.; Majumdar, S.; Konrath, K.M.; et al. Induction of tier-2 neutralizing antibodies in mice with a DNA-encoded HIV envelope native like trimer. Nat. Commun. 2022, 13, 695. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequencing Statistics per Sample | ||||
|---|---|---|---|---|
| Sample | Read Count | Average Read Length | Depth of Coverage (x) | Average Quality per Read |
| Untreated HIV-1NL4-3 Days 0–12 | 1,798,483 | 150 | ~29,411 | ≥39 |
| Untreated HIV-1NL4-3 Days 13–24 | 4,693,741 | 150 | ~76,736 | ≥39 |
| MoMo30 Treated HIV-1NL4-3 Days 0–12 | 2,461,268 | 150 | ~40,247 | ≥39 |
| MoMo30 Treated HIV-1NL4-3 Days 13–24 | 3,692,770 | 150 | ~60,391 | ≥39 |
| Changes in Potential N-Linked Glycosylation Sites (PNGSs) in HIV-1NL4-3 Envelope Gene | |||
|---|---|---|---|
| Condition | # of PNGSs Days 0–12 | # of PNGSs Days 13–24 | Change from Reference |
| Untreated HIV-1NL4-3 | 31 | 31 | No change |
| MoMo30 Treated HIV-1NL4-3 | 31 | 21 | Decrease of 10 PNGSs during Days 13–24 |
| Amino Acid Changes in gp120 Variable Loops After MoMo30 Treatment | ||
|---|---|---|
| Variable Loop | Amino Acid Changes After MoMo30 Treatment | Potential Impact |
| V1 | No changes observed | — |
| V2 | - Arg263→Pro - Asn265→Tyr/Asp/Ile - Asn267→Leu - Thr268→Pro | - Loss of positive charge and flexibility (Arg→Pro) - Asn265 mutations may disrupt glycosylation site - Asn267→Leu removes potential glycan and adds hydrophobicity - Proline at 268 may introduce kink in backbone |
| V3 | - Gln275→Val/Gly - Arg276→Phe/Leu - Gly277→Leu/Ser - Pro278→Ser | - Shift from polar to hydrophobic (Gln→Val) - Loss of positive charge and introduction of bulky hydrophobic residues (Arg276) - Possible structural rigidity increase (Gly→Leu/Ser) - Pro→Ser substitution may reduce loop constraint |
| V4 | - Gln293_Ala294 ins12 - Ala294→Asp - His295→Leu | - Insertion likely alters local structure/loop length - Ala→Asp introduces negative charge - His→Leu shifts from polar to hydrophobic, may affect receptor interactions |
| V5 | - Arg280→Stop codon - Asp439→Glu | - Premature stop likely truncates gp120, disrupting structure - Asp→Glu is conservative but may alter local charge density |
| Key Amino Acid Changes in gp41 Functional Domains After MoMo30 Treatment | ||
|---|---|---|
| Primary Domain | Amino Acid Changes After MoMo30 Treatment | Potential Impact |
| N-TERMINAL HEPTAD REPEAT (HR1) | - Arg28→Ser - Ala32→Cys/Ser - Gln33→Leu | - May disrupt coiled-coil heptad repeat; may affect bundle formation - Introduction of cysteine may lead to aberrant disulfide bonding - Hydrophobic substitution may alter helical interface |
| HOMOTRIMER INTERFACE (HR1 LOOP) | - Ala78→Ser - Val79→Glu - Trp94→Cys | - Polar substitution may affect interface packing - Negative charge could disrupt hydrophobic core - Loss of bulky aromatic residue; may impair trimer stability |
| C-TERMINAL HEPTAD REPEAT (HR2) | - Leu112→Stop - Ser115→Val - Glu119→Stop | - Premature truncation; likely disrupts HR2 function - Hydrophobic substitution may affect helical stability - Early stop codon likely abolishes six-helix bundle formation |
| MEMBRANE PROXIMAL EXTERNAL REGION (MPER) | - Glu121→Cys - Trp137→Ser - Trp144→Leu | - Cysteine introduction may affect folding or disulfide linkage - Loss of aromatic residue; may reduce membrane interaction - Hydrophobic substitution; could impair MPER-mediated fusion |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coleman, M.I.; Khan, M.B.; Gbodossou, E.; Diop, A.; DeBarros, K.; Bond, V.C.; Floyd, V.; Kondwani, K.; Rice, V.M.; Powell, M.D. Plant Lectin, MoMo30, Pressures HIV-1 to Select for Variants with Deleted N-Linked Glycosylation Sites. Viruses 2025, 17, 910. https://doi.org/10.3390/v17070910
Coleman MI, Khan MB, Gbodossou E, Diop A, DeBarros K, Bond VC, Floyd V, Kondwani K, Rice VM, Powell MD. Plant Lectin, MoMo30, Pressures HIV-1 to Select for Variants with Deleted N-Linked Glycosylation Sites. Viruses. 2025; 17(7):910. https://doi.org/10.3390/v17070910
Chicago/Turabian StyleColeman, Morgan I., Mahfuz B. Khan, Erick Gbodossou, Amad Diop, Kenya DeBarros, Vincent C. Bond, Virginia Floyd, Kofi Kondwani, Valerie Montgomery Rice, and Michael D. Powell. 2025. "Plant Lectin, MoMo30, Pressures HIV-1 to Select for Variants with Deleted N-Linked Glycosylation Sites" Viruses 17, no. 7: 910. https://doi.org/10.3390/v17070910
APA StyleColeman, M. I., Khan, M. B., Gbodossou, E., Diop, A., DeBarros, K., Bond, V. C., Floyd, V., Kondwani, K., Rice, V. M., & Powell, M. D. (2025). Plant Lectin, MoMo30, Pressures HIV-1 to Select for Variants with Deleted N-Linked Glycosylation Sites. Viruses, 17(7), 910. https://doi.org/10.3390/v17070910