
Orthohantaviruses: An Overview of the Current Status of Diagnostics and Surveillance

 , , ,
, , ,
Abstract
1. Introduction
2. Orthohantavirus Genome, Replication, and Reassortment
3. Clinical Aspects in Humans and Other Animals
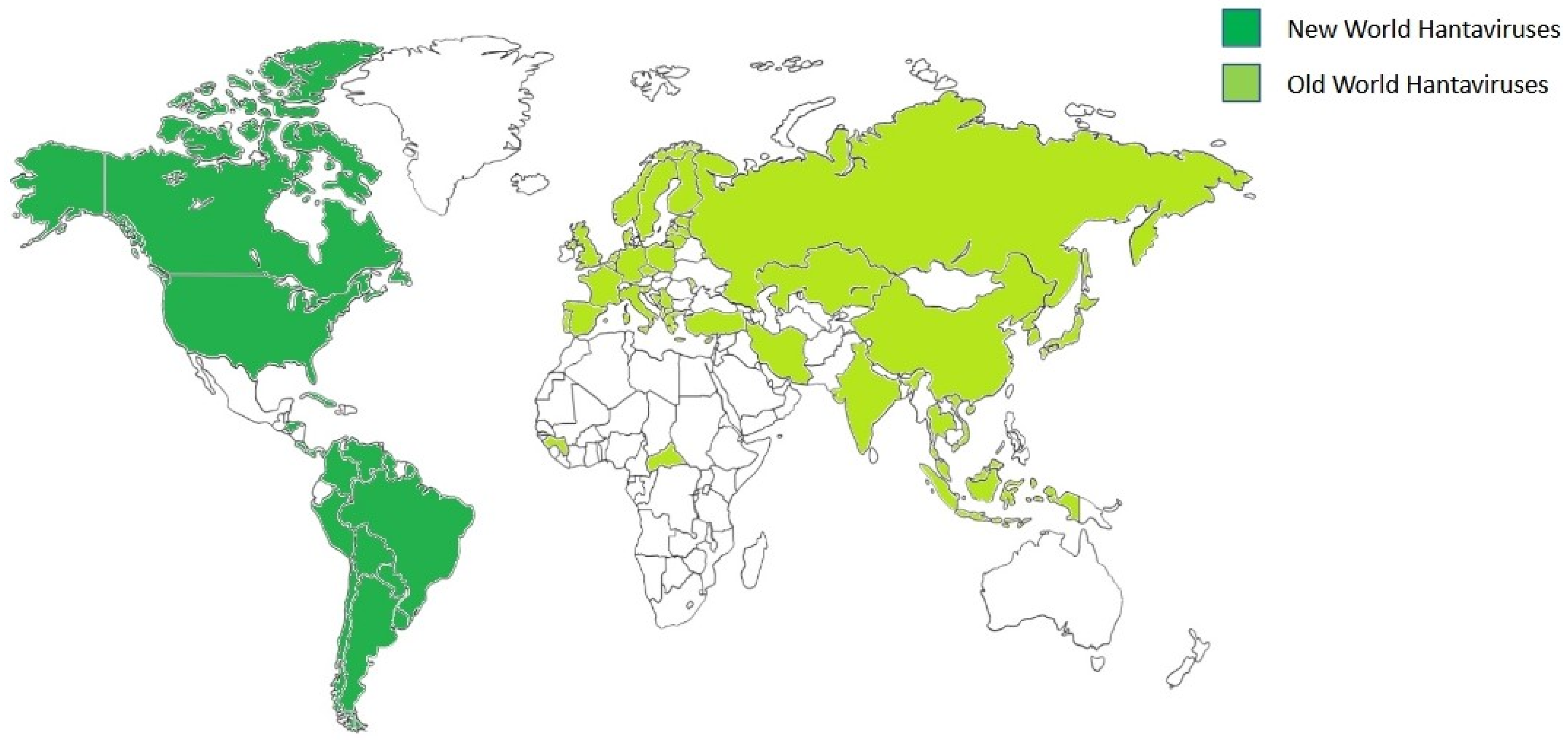
4. Epidemiology
5. Diagnosis
5.1. Serological Diagnosis
5.1.1. ELISA
5.1.2. Immunofluorescence Assay
5.1.3. Immunoblot Assay
5.1.4. Neutralisation Test
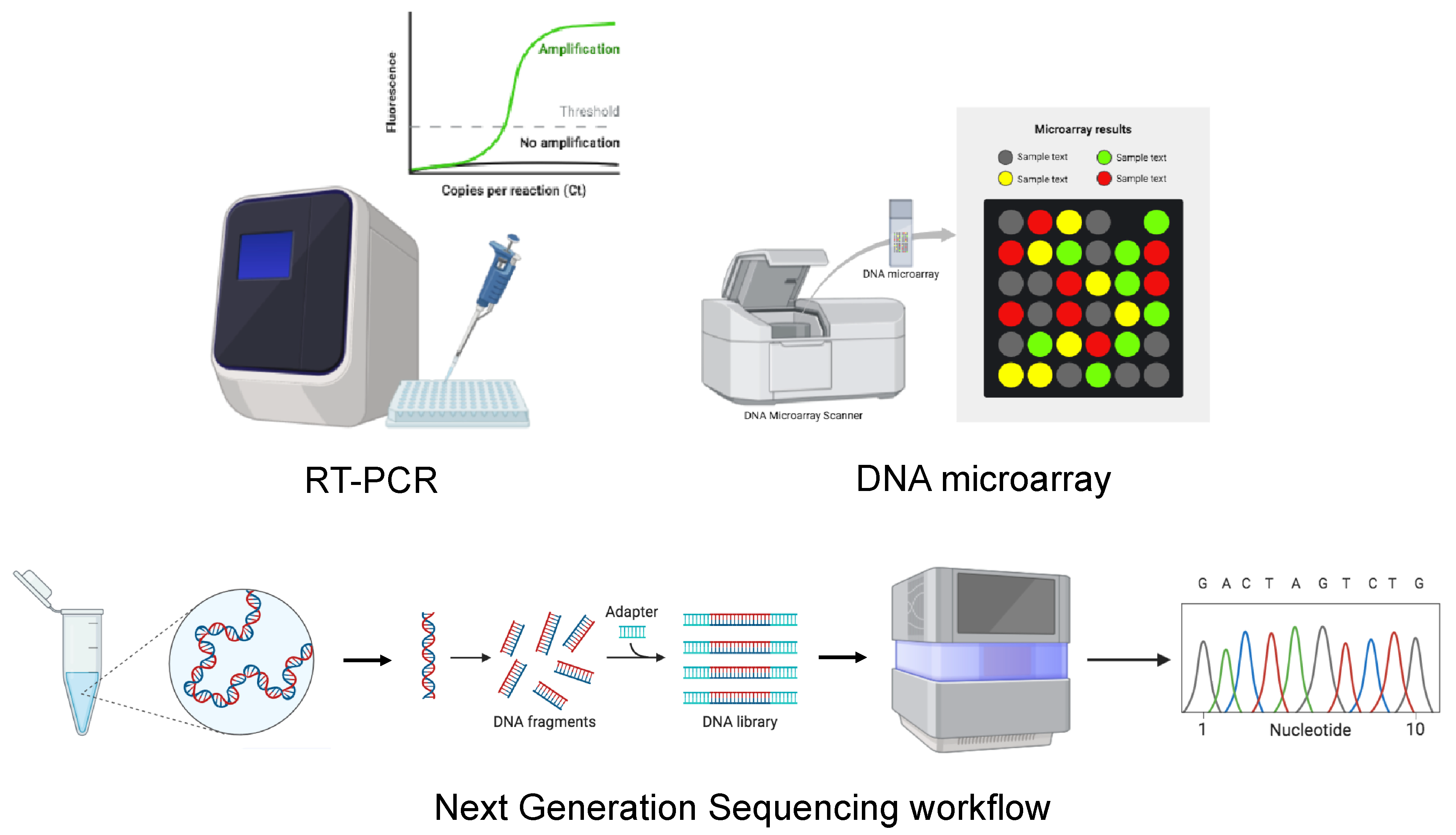
5.2. Molecular Diagnosis and Sequencing
5.2.1. RT-PCR
5.2.2. DNA Microarray
5.2.3. Viral Sequencing
5.3. Virus Isolation in Cell Culture
5.4. Immunohistochemistry
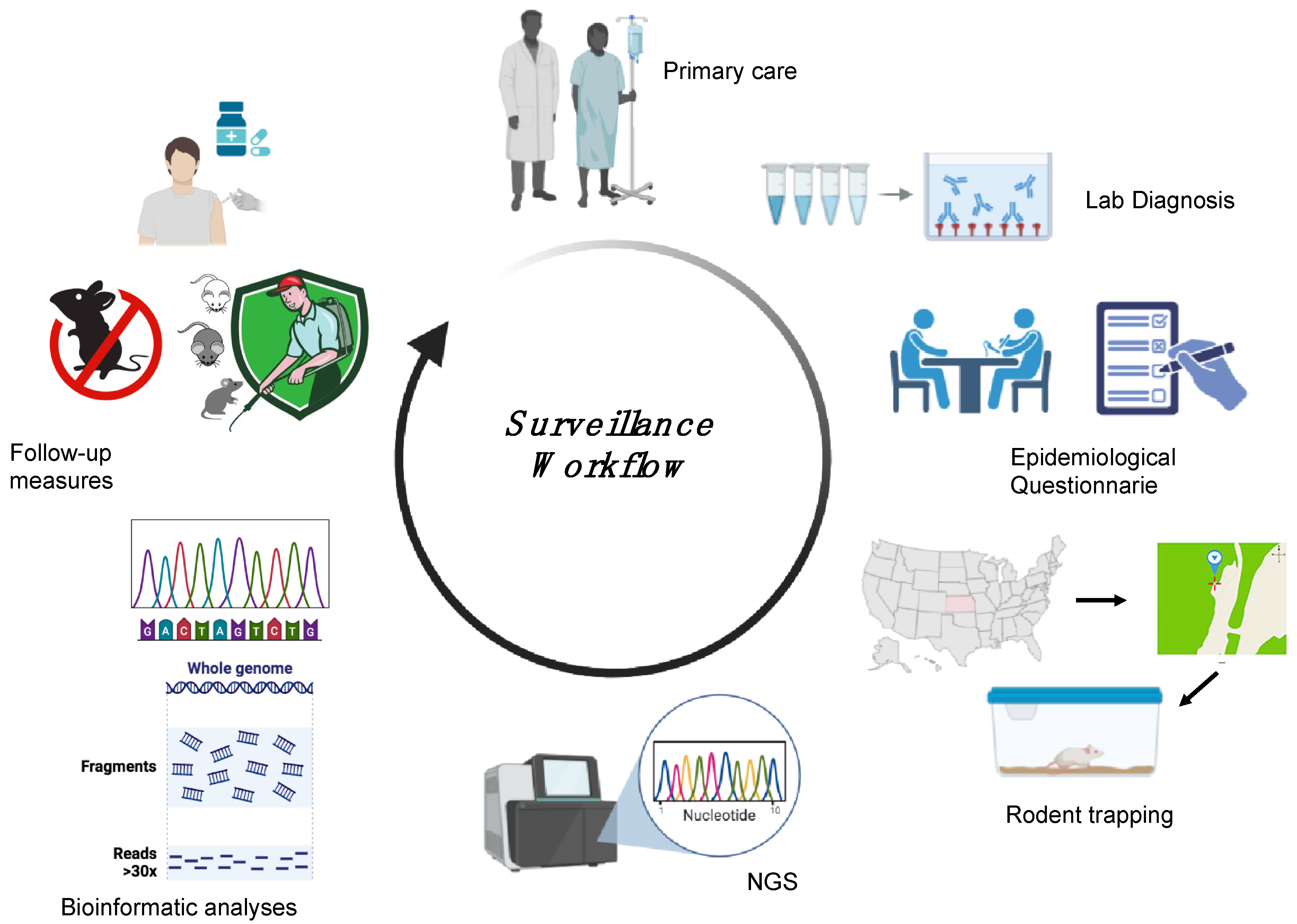
6. HTV Surveillance in Humans and Animals
7. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ANDV | Andes virus |
| ARAV | Araraquara virus |
| BSL-3 | biosafety level 3 |
| CDC | Centers for Disease Control and Prevention |
| CHOV | Choclo virus |
| DOBV | Dobrava-Belgrade Virus |
| EBOV | Ebola Virus |
| ECDC | European Centre for Disease Prevention and Control |
| ELISA | enzyme-linked immunosorbent assay |
| EQA | external quality assessment |
| ERGIC | endoplasmic reticulum-Golgi intermediate compartment |
| FRNT | focus reduction neutralisation test |
| GPC | glycoprotein precursor |
| GWAS | genome wide association study |
| HCPS | hantavirus cardio-pulmonary syndrome |
| HFRS | haemorrhagic fever with renal syndrome |
| HPS | hantavirus pulmonary syndrome |
| HTNV | Hantaan Virus |
| HTVs | hantaviruses |
| IBA | immunoblot assay |
| IFA | immunofluorescence assay |
| IFN | interferon |
| JUQ | Juquitiba virus |
| LANV | Laguna Negra virus |
| LASV | Lassa Virus |
| MNT | microneutralisation test |
| NGS | next-generation sequencing |
| NS | non-structural |
| p.f.u. | plaque-forming units |
| PPNT | pseudoparticle neutralisation test |
| PRNT | plaque reduction neutralisation test |
| PUUV | Puumala Virus |
| RdRp | RNA-dependent RNA polymerase |
| rNp | recombinant nucleocapsid proteins |
| SEOV | Seoul Virus |
| SISPA | single-primer amplification |
| SNV | Sin Nombre virus |
| TESSy | European Surveillance System |
| WHO | World Health Organization |
| ZIKV | Zika Virus |
References
- Goodfellow, S.M.; Nofchissey, R.A.; Ye, C.; Dunnum, J.L.; Cook, J.A.; Bradfute, S.B. Use of a Novel Detection Tool to Survey Orthohantaviruses in Wild-Caught Rodent Populations. Viruses 2022, 14, 682. [Google Scholar] [CrossRef] [PubMed]
- Hussein, I.T.M.; Haseeb, A.; Haque, A.; Mir, M.A. Recent Advances in Hantavirus Molecular Biology and Disease. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 74, pp. 35–75. ISBN 978-0-12-387022-3. [Google Scholar]
- Watson, D.C.; Sargianou, M.; Papa, A.; Chra, P.; Starakis, I.; Panos, G. Epidemiology of Hantavirus Infections in Humans: A Comprehensive, Global Overview. Crit. Rev. Microbiol. 2014, 40, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Noack, D.; Goeijenbier, M.; Reusken, C.B.E.M.; Koopmans, M.P.G.; Rockx, B.H.G. Orthohantavirus Pathogenesis and Cell Tropism. Front. Cell. Infect. Microbiol. 2020, 10, 399. [Google Scholar] [CrossRef] [PubMed]
- Clement, J.; Maes, P.; Van Ranst, M. Hantaviruses in the Old and New World. In Perspectives in Medical Virology; Elsevier: Amsterdam, The Netherlands, 2006; Volume 16, pp. 161–177. ISBN 978-0-444-52074-6. [Google Scholar]
- Chen, R.-X.; Gong, H.-Y.; Wang, X.; Sun, M.-H.; Ji, Y.-F.; Tan, S.-M.; Chen, J.-M.; Shao, J.-W.; Liao, M. Zoonotic Hantaviridae with Global Public Health Significance. Viruses 2023, 15, 1705. [Google Scholar] [CrossRef]
- Tkachenko, E.A.; Morozov, V.G.; Dzagurova, T.K.; Yunicheva, Y.V.; Pilikova, O.M.; Zavora, D.L.; Ishmukhametov, A.A.; Gorodin, V.N.; Bakhtina, V.A.; Zagidullin, I.M.; et al. Etiologic and Clinical Epidemiological Features of Hemorrhagic Fever with Renal Syndrome (HFRS) in the Krasnodar Krai. Epidemiol. Infect. Dis. 2016, 21, 22–30. [Google Scholar] [CrossRef]
- Lee, H.W.; Baek, L.J.; Johnson, K.M. Isolation of Hantaan Virus, the Etiologic Agent of Korean Hemorrhagic Fever, from Wild Urban Rats. J. Infect. Dis. 1982, 146, 638–644. [Google Scholar] [CrossRef]
- Avsic-Zupanc, T.; Xiao, S.; Stojanovic, R.; Gligic, A.; Van Der Groen, G.; Leduc, J.W. Characterization of Dobrava Virus: A Hantavirus from Slovenia, Yugoslavia. J. Med. Virol. 1992, 38, 132–137. [Google Scholar] [CrossRef]
- Brummer-Korvenkontio, M.; Vaheri, A.; Hovi, T.; Von Bonsdorff, C.-H.; Vuorimies, J.; Manni, T.; Penttinen, K.; Oker-Blom, N.; Lahdevirta, J. Nephropathia Epidemica: Detection of Antigen in Bank Voles and Serologic Diagnosis of Human Infection. J. Infect. Dis. 1980, 141, 131–134. [Google Scholar] [CrossRef]
- Chandy, S.; Mathai, D. Globally Emerging Hantaviruses: An Overview. Indian J. Med. Microbiol. 2017, 35, 165–175. [Google Scholar] [CrossRef]
- Klempa, B.; Tkachenko, E.A.; Dzagurova, T.K.; Yunicheva, Y.V.; Morozov, V.G.; Okulova, N.M.; Slyusareva, G.P.; Smirnov, A.; Kruger, D.H. Hemorrhagic Fever with Renal Syndrome Caused by 2 Lineages of Dobrava Hantavirus, Russia1. Emerg. Infect. Dis. 2008, 14, 617–625. [Google Scholar] [CrossRef]
- Kolodziej, M.; Melgies, A.; Joniec-Wiechetek, J.; Michalski, A.; Nowakowska, A.; Pitucha, G.; Niemcewicz, M. First Molecular Characterization of Dobrava-Belgrade Virus Found in Apodemus Flavicollis in Poland. Ann. Agric. Environ. Med. 2018, 25, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [PubMed]
- Madrières, S.; Tatard, C.; Murri, S.; Vulin, J.; Galan, M.; Piry, S.; Pulido, C.; Loiseau, A.; Artige, E.; Benoit, L.; et al. How Bank Vole-PUUV Interactions Influence the Eco-Evolutionary Processes Driving Nephropathia Epidemica Epidemiology—An Experimental and Genomic Approach. Pathogens 2020, 9, 789. [Google Scholar] [CrossRef]
- Cosseddu, G.M.; Sozio, G.; Valleriani, F.; Di Gennaro, A.; Pascucci, I.; Gavaudan, S.; Marianneau, P.; Monaco, F. Serological Survey of Hantavirus and Flavivirus Among Wild Rodents in Central Italy. Vector-Borne Zoonotic Dis. 2017, 17, 777–779. [Google Scholar] [CrossRef]
- Voutilainen, L.; Kallio, E.R.; Niemimaa, J.; Vapalahti, O.; Henttonen, H. Temporal Dynamics of Puumala Hantavirus Infection in Cyclic Populations of Bank Voles. Sci. Rep. 2016, 6, 21323. [Google Scholar] [CrossRef]
- Kabwe, E.; Davidyuk, Y.; Shamsutdinov, A.; Garanina, E.; Martynova, E.; Kitaeva, K.; Malisheni, M.; Isaeva, G.; Savitskaya, T.; Urbanowicz, R.A.; et al. Orthohantaviruses, Emerging Zoonotic Pathogens. Pathogens 2020, 9, 775. [Google Scholar] [CrossRef]
- Vial, P.A.; Ferrés, M.; Vial, C.; Klingström, J.; Ahlm, C.; López, R.; Le Corre, N.; Mertz, G.J. Hantavirus in Humans: A Review of Clinical Aspects and Management. Lancet Infect. Dis. 2023, 23, e371–e382. [Google Scholar] [CrossRef]
- Armien, B.; Pascale, J.M.; Muñoz, C.; Mariñas, J.; Núñez, H.; Herrera, M.; Trujillo, J.; Sánchez, D.; Mendoza, Y.; Hjelle, B.; et al. Hantavirus Fever without Pulmonary Syndrome in Panama. Am. Soc. Trop. Med. Hyg. 2013, 89, 489–494. [Google Scholar] [CrossRef]
- Ferrer, J.F.; Jonsson, C.B.; Esteban, E.; Galligan, D.; Basombrio, M.A.; Peralta-Ramos, M.; Bharadwaj, M.; Torrez-Martinez, N.; Callahan, J.; Segovia, A.; et al. High Prevalence of Hantavirus Infection in Indian Communities of the Paraguayan and Argentinean Gran Chaco. Am. J. Trop. Med. Hyg. 1998, 59, 438–444. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Lima, M. Myths and Facts on Ratadas: Bamboo Blooms, Rainfall Peaks and Rodent Outbreaks in South America. Austral Ecol. 2003, 28, 237–251. [Google Scholar] [CrossRef]
- Martínez, V.P.; Di Paola, N.; Alonso, D.O.; Pérez-Sautu, U.; Bellomo, C.M.; Iglesias, A.A.; Coelho, R.M.; López, B.; Periolo, N.; Larson, P.A.; et al. “Super-Spreaders” and Person-to-Person Transmission of Andes Virus in Argentina. N. Engl. J. Med. 2020, 383, 2230–2241. [Google Scholar] [CrossRef] [PubMed]
- Hardestam, J.; Lundkvist, Å.; Klingström, J. Sensitivity of Andes Hantavirus to Antiviral Effect of Human Saliva. Emerg. Infect. Dis. 2009, 15, 1140–1142. [Google Scholar] [CrossRef] [PubMed]
- Schmaljohn, C.S.; Hasty, S.E.; Harrison, S.A.; Dalrymple, J.M. Characterization of Hantaan Virions, the Prototype Virus of Hemorrhagic Fever with Renal Syndrome. J. Infect. Dis. 1983, 148, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Kukkonen, S.K.J.; Vaheri, A.; Plyusnin, A. L Protein, the RNA-Dependent RNA Polymerase of Hantaviruses. Arch. Virol. 2005, 150, 533–556. [Google Scholar] [CrossRef]
- Muyangwa, M.; Martynova, E.V.; Khaiboullina, S.F.; Morzunov, S.P.; Rizvanov, A.A. Hantaviral Proteins: Structure, Functions, and Role in Hantavirus Infection. Front. Microbiol. 2015, 6, 1326. [Google Scholar] [CrossRef]
- Vera-Otarola, J.; Solis, L.; Soto-Rifo, R.; Ricci, E.P.; Pino, K.; Tischler, N.D.; Ohlmann, T.; Darlix, J.-L.; López-Lastra, M. The Andes Hantavirus NSs Protein Is Expressed from the Viral Small mRNA by a Leaky Scanning Mechanism. J. Virol. 2012, 86, 2176–2187. [Google Scholar] [CrossRef]
- Davidyuk, Y.; Shamsutdinov, A.; Kabwe, E.; Ismagilova, R.; Martynova, E.; Belyaev, A.; Shuralev, E.; Trifonov, V.; Savitskaya, T.; Isaeva, G.; et al. Prevalence of the Puumala Orthohantavirus Strains in the Pre-Kama Area of the Republic of Tatarstan, Russia. Pathogens 2020, 9, 540. [Google Scholar] [CrossRef]
- Khaiboullina, S.; Morzunov, S.; St. Jeor, S. Hantaviruses: Molecular Biology, Evolution and Pathogenesis. Curr. Mol. Med. 2005, 5, 773–790. [Google Scholar] [CrossRef]
- Flick, K.; Hooper, J.W.; Schmaljohn, C.S.; Pettersson, R.F.; Feldmann, H.; Flick, R. Rescue of Hantaan Virus Minigenomes. Virology 2003, 306, 219–224. [Google Scholar] [CrossRef]
- Jääskeläinen, K.M.; Kaukinen, P.; Minskaya, E.S.; Plyusnina, A.; Vapalahti, O.; Elliott, R.M.; Weber, F.; Vaheri, A.; Plyusnin, A. Tula and Puumala Hantavirus NSs ORFs Are Functional and the Products Inhibit Activation of the Interferon-beta Promoter. J. Med. Virol. 2007, 79, 1527–1536. [Google Scholar] [CrossRef]
- Jin, M.; Park, J.; Lee, S.; Park, B.; Shin, J.; Song, K.-J.; Ahn, T.-I.; Hwang, S.-Y.; Ahn, B.-Y.; Ahn, K. Hantaan Virus Enters Cells by Clathrin-Dependent Receptor-Mediated Endocytosis. Virology 2002, 294, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, H.N.; Chung, D.-H.; Plane, S.J.; Sztul, E.; Chu, Y.; Guttieri, M.C.; McDowell, M.; Ali, G.; Jonsson, C.B. Dynein-Dependent Transport of the Hantaan Virus Nucleocapsid Protein to the Endoplasmic Reticulum-Golgi Intermediate Compartment. J. Virol. 2007, 81, 8634–8647. [Google Scholar] [CrossRef] [PubMed]
- Mittler, E.; Dieterle, M.E.; Kleinfelter, L.M.; Slough, M.M.; Chandran, K.; Jangra, R.K. Hantavirus Entry: Perspectives and Recent Advances. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 104, pp. 185–224. ISBN 978-0-12-818394-6. [Google Scholar]
- Mir, M.A.; Panganiban, A.T. The Triplet Repeats of the Sin Nombre Hantavirus 5′ Untranslated Region Are Sufficient in Cis for Nucleocapsid-Mediated Translation Initiation. J. Virol. 2010, 84, 8937–8944. [Google Scholar] [CrossRef]
- Meier, K.; Thorkelsson, S.R.; Quemin, E.R.J.; Rosenthal, M. Correction: Meier et al. Hantavirus Replication Cycle—An Updated Structural Virology Perspective. Viruses 2021, 13, 1561. Viruses 2023, 15, 273. [Google Scholar] [CrossRef]
- Koehler, F.C.; Di Cristanziano, V.; Späth, M.R.; Hoyer-Allo, K.J.R.; Wanken, M.; Müller, R.-U.; Burst, V. The Kidney in Hantavirus Infection—Epidemiology, Virology, Pathophysiology, Clinical Presentation, Diagnosis and Management. Clin. Kidney J. 2022, 15, 1231–1252. [Google Scholar] [CrossRef]
- Riccò, M.; Ferraro, P.; Peruzzi, S.; Balzarini, F.; Ranzieri, S. Hantaviruses in Agricultural and Forestry Workers: Knowledge, Attitudes and Practices in Italian Physicians. Trop. Med. Infect. Dis. 2021, 6, 169. [Google Scholar] [CrossRef]
- Li, D.; Schmaljohn, A.L.; Anderson, K.; Schmaljohn, C.S. Complete Nucleotide Sequences of the M and S Segments of Two Hantavirus Isolates from California: Evidence for Reassortment in Nature among Viruses Related to Hantavirus Pulmonary Syndrome. Virology 1995, 206, 973–983. [Google Scholar] [CrossRef]
- Razzauti, M.; Plyusnina, A.; Henttonen, H.; Plyusnin, A. Accumulation of Point Mutations and Reassortment of Genomic RNA Segments Are Involved in the Microevolution of Puumala Hantavirus in a Bank Vole (Myodes Glareolus) Population. J. Gen. Virol. 2008, 89, 1649–1660. [Google Scholar] [CrossRef]
- Razzauti, M.; Plyusnina, A.; Sironen, T.; Henttonen, H.; Plyusnin, A. Analysis of Puumala Hantavirus in a Bank Vole Population in Northern Finland: Evidence for Co-Circulation of Two Genetic Lineages and Frequent Reassortment between Strains. J. Gen. Virol. 2009, 90, 1923–1931. [Google Scholar] [CrossRef]
- Klempa, B. Reassortment Events in the Evolution of Hantaviruses. Virus Genes 2018, 54, 638–646. [Google Scholar] [CrossRef]
- Myhrman, G. Nephropathia Epidemica a New Infectious Disease in Northern Scandinavia. J. Intern. Med. 1951, 140, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Smadel, J.E. Epidemic Hemorrhagic Fever. Am. J. Public Health Nations Health 1953, 43, 1327–1330. [Google Scholar] [CrossRef] [PubMed]
- Powell, G.M. Hemorrhagic Fever: A Study of 300 Cases. Medicine 1954, 33, 97–153. [Google Scholar] [CrossRef] [PubMed]
- Lähdevirta, J. Nephropathia Epidemica in Finland. A Clinical Histological and Epidemiological Study. Ann. Clin. Res. 1971, 3, 1–54. [Google Scholar]
- Noh, J.Y.; Cheong, H.J.; Song, J.Y.; Kim, W.J.; Song, K.-J.; Klein, T.A.; Lee, S.H.; Yanagihara, R.; Song, J.-W. Clinical and Molecular Epidemiological Features of Hemorrhagic Fever with Renal Syndrome in Korea over a 10-Year Period. J. Clin. Virol. 2013, 58, 11–17. [Google Scholar] [CrossRef]
- Jiang, H.; Du, H.; Wang, L.M.; Wang, P.Z.; Bai, X.F. Hemorrhagic Fever with Renal Syndrome: Pathogenesis and Clinical Picture. Front. Cell. Infect. Microbiol. 2016, 6, 1. [Google Scholar] [CrossRef]
- Zou, L.-X.; Chen, M.-J.; Sun, L. Haemorrhagic Fever with Renal Syndrome: Literature Review and Distribution Analysis in China. Int. J. Infect. Dis. 2016, 43, 95–100. [Google Scholar] [CrossRef]
- Duchin, J.S.; Koster, F.T.; Peters, C.J.; Simpson, G.L.; Tempest, B.; Zaki, S.R.; Ksiazek, T.G.; Rollin, P.E.; Nichol, S.; Umland, E.T.; et al. Hantavirus Pulmonary Syndrome: A Clinical Description of 17 Patients with a Newly Recognized Disease. N. Engl. J. Med. 1994, 330, 949–955. [Google Scholar] [CrossRef]
- Ketai, L.H.; Williamson, M.R.; Telepak, R.J.; Levy, H.; Koster, F.T.; Nolte, K.B.; Allen, S.E. Hantavirus Pulmonary Syndrome: Radiographic Findings in 16 Patients. Radiology 1994, 191, 665–668. [Google Scholar] [CrossRef]
- Llah, S.T.; Mir, S.; Sharif, S.; Khan, S.; Mir, M.A. Hantavirus Induced Cardiopulmonary Syndrome: A Public Health Concern. J. Med. Virol. 2018, 90, 1003–1009. [Google Scholar] [CrossRef]
- CDC. Outbreak of Acute Illness--Southwestern United States, 1993. MMWR Morb Mortal Wkly Rep. 1993, 42, 421–424. [Google Scholar]
- Leopardi, S.; Drzewnioková, P.; Baggieri, M.; Marchi, A.; Bucci, P.; Bregoli, M.; De Benedictis, P.; Gobbo, F.; Bellinati, L.; Citterio, C.; et al. Identification of Dobrava-Belgrade Virus in Apodemus Flavicollis from North-Eastern Italy during Enhanced Mortality. Viruses 2022, 14, 1241. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-K.; No, J.S.; Lee, D.; Jung, J.; Park, H.; Yi, Y.; Kim, J.-A.; Lee, S.-H.; Kim, Y.; Park, S.; et al. Active Targeted Surveillance to Identify Sites of Emergence of Hantavirus. Clin. Infect. Dis. 2020, 70, 464–473. [Google Scholar] [CrossRef]
- Tortosa, F.; Perre, F.; Tognetti, C.; Lossetti, L.; Carrasco, G.; Guaresti, G.; Iglesias, A.; Espasandin, Y.; Izcovich, A. Seroprevalence of Hantavirus Infection in Non-Epidemic Settings over Four Decades: A Systematic Review and Meta-Analysis. BMC Public Health 2024, 24, 2553. [Google Scholar] [CrossRef]
- Acham-Roschitz, B.; Aberle, S.W.; Pirker, N.; Kaulfersch, W.; Boehm, M.; Roedl, S.; Zenz, W.; Ring, E.; Mache, C.J. Nephropathia Epidemica (Puumala Virus Infection) in Austrian Children. Pediatr. Infect. Dis. J. 2010, 29, 874–876. [Google Scholar] [CrossRef]
- Echterdiek, F.; Kitterer, D.; Alscher, M.D.; Schwenger, V.; Ruckenbrod, B.; Bald, M.; Latus, J. Clinical Course of Hantavirus-Induced Nephropathia Epidemica in Children Compared to Adults in Germany—Analysis of 317 Patients. Pediatr. Nephrol. 2019, 34, 1247–1252. [Google Scholar] [CrossRef]
- Gilson, G.J.; Maciulla, J.A.; Nevils, B.G.; Izquierdo, L.E.; Chatterjee, M.S.; Curet, L.B. Hantavirus Pulmonary Syndrome Complicating Pregnancy. Am. J. Obstet. Gynecol. 1994, 171, 550–554. [Google Scholar] [CrossRef]
- Hofmann, J.; Führer, A.; Bolz, M.; Waldschläger-Terpe, J.; Meier, M.; Lüdders, D.; Enders, M.; Oltmann, A.; Meisel, H.; Krüger, D.H. Hantavirus Infections by Puumala or Dobrava-Belgrade Virus in Pregnant Women. J. Clin. Virol. 2012, 55, 266–269. [Google Scholar] [CrossRef]
- Ji, F.; Zhao, W.; Liu, H.; Zheng, H.; Wang, S.; He, C.; Wang, W.; Zhang, R.; Bai, D.; Tian, C.; et al. Hemorrhagic Fever with Renal Syndrome Caused by Hantaan Virus Infection in Four Pregnant Chinese Women. J. Med. Virol. 2017, 89, 1865–1870. [Google Scholar] [CrossRef]
- Bellomo, C.; Alonso, D.; Coelho, R.; Iglesias, A.; Periolo, N.; Martínez, V.P. A Newborn Infected by Andes Virus Suggests Novel Routes of Hantavirus Transmission: A Case Report. Clin. Microbiol. Infect. 2020, 26, 130–131. [Google Scholar] [CrossRef]
- Ferrés, M.; Martínez-Valdebenito, C.; Angulo, J.; Henríquez, C.; Vera-Otárola, J.; Vergara, M.J.; Pérez, J.; Fernández, J.; Sotomayor, V.; Valdés, M.F.; et al. Mother-to-Child Transmission of Andes Virus through Breast Milk, Chile1. Emerg. Infect. Dis. 2020, 26, 1885–1888. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yu, C.; Chen, Y.; Nian, B.; Chai, M.; Maimaiti, D.; Xu, D.; Zang, X. Hemorrhagic Fever with Renal Syndrome Complicated by Acute Pancreatitis, High Intraocular Pressure, and Pulmonary Involvement: A Case Report. Infect. Drug Resist. 2024, 17, 1919–1925. [Google Scholar] [CrossRef] [PubMed]
- Geeraedts, F.; Wevers, M.; Bosma, F.; Boer, M.D.; Brinkman, J.N.; Delsing, C.; GeurtsvanKessel, C.; Rockx, B.; Van Der Zanden, A.; Laverman, G.D. Use of a Diagnostic Puumala Virus Real-Time RT-PCR in an Orthohantavirus Endemic Region in the Netherlands. Microbiol. Spectr. 2024, 12, e03813-23. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.-D.; Sun, J.-J.; Liu, H.-L. Potential Clinical Biomarkers in Monitoring the Severity of Hantaan Virus Infection. Cytokine 2023, 170, 156340. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Annual Epidemiological Report for 2016: Hantavirus Infection; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2018. [Google Scholar]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.-W. Hantaviruses: Rediscovery and New Beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef]
- CDC. Clinician Brief: Hantavirus Pulmonary Syndrome (HPS). Available online: https://www.cdc.gov/hantavirus/hcp/clinical-overview/hps.html (accessed on 23 May 2024).
- CDC. Reported Cases of Hantavirus Disease. Available online: https://www.cdc.gov/hantavirus/data-research/cases/index.html (accessed on 26 June 2024).
- European Centre for Disease Prevention and Control. Hantavirus Infection Annual Epidemiological Report for 2020; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2023. [Google Scholar]
- Wang, Y.X.G.; Voutilainen, L.; Aminikhah, M.; Helle, H.; Huitu, O.; Laakkonen, J.; Lindén, A.; Niemimaa, J.; Sane, J.; Sironen, T.; et al. The Impact of Wildlife and Environmental Factors on Hantavirus Infection in the Host and Its Translation into Human Risk. Proc. R. Soc. B 2023, 290, 20222470. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Surveillance and Updates for Hantavirus. Available online: https://www.ecdc.europa.eu/en/hantavirus-infection/surveillance-and-disease-data (accessed on 20 January 2025).
- Mustonen, J.; Henttonen, H.; Vaheri, A. Hantavirus Infections among Military Forces. Mil. Med. 2024, 189, 551–555. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Disease Information about Hantavirus. Available online: https://www.ecdc.europa.eu/en/hantavirus-infection/facts (accessed on 20 January 2025).
- Munir, N.; Jahangeer, M.; Hussain, S.; Mahmood, Z.; Ashiq, M.; Ehsan, F.; Akram, M.; Ali Shah, S.M.; Riaz, M.; Sana, A. Hantavirus Diseases Pathophysiology, Their Diagnostic Strategies and Therapeutic Approaches: A Review. Clin. Exp. Pharmacol. Physiol. 2021, 48, 20–34. [Google Scholar] [CrossRef]
- Escadafal, C.; Avšič-Županc, T.; Vapalahti, O.; Niklasson, B.; Teichmann, A.; Niedrig, M.; Donoso-Mantke, O. Second External Quality Assurance Study for the Serological Diagnosis of Hantaviruses in Europe. PLoS Negl. Trop. Dis. 2012, 6, e1607. [Google Scholar] [CrossRef]
- Lederer, S.; Lattwein, E.; Hanke, M.; Sonnenberg, K.; Stoecker, W.; Lundkvist, Å.; Vaheri, A.; Vapalahti, O.; Chan, P.K.S.; Feldmann, H.; et al. Indirect Immunofluorescence Assay for the Simultaneous Detection of Antibodies against Clinically Important Old and New World Hantaviruses. PLoS Negl. Trop. Dis. 2013, 7, e2157. [Google Scholar] [CrossRef]
- Li, W.; Cao, S.; Zhang, Q.; Li, J.; Zhang, S.; Wu, W.; Qu, J.; Li, C.; Liang, M.; Li, D. Comparison of Serological Assays to Titrate Hantaan and Seoul Hantavirus-Specific Antibodies. Virol. J. 2017, 14, 133. [Google Scholar] [CrossRef] [PubMed]
- Kramski, M.; Meisel, H.; Klempa, B.; Krüger, D.H.; Pauli, G.; Nitsche, A. Detection and Typing of Human Pathogenic Hantaviruses by Real-Time Reverse Transcription-PCR and Pyrosequencing. Clin. Chem. 2007, 53, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Altona Diagnostics. RealStar® Hantavirus-HFRS RT-PCR Kit 1.0 RUO. Available online: https://altona-diagnostics.com/product/realstar-hantavirus-hfrs-rt-pcr-kit-1-0-ruo/ (accessed on 1 February 2025).
- Altona Diagnostics. RealStar® Hantavirus-HPS RT-PCR Kit 1.0 RUO. Available online: https://altona-diagnostics.com/product/realstar-hantavirus-hps-rt-pcr-kit-1-0-ruo/ (accessed on 1 February 2025).
- Nunes, B.T.D.; De Mendonça, M.H.R.; Simith, D.D.B.; Moraes, A.F.; Cardoso, C.C.; Prazeres, I.T.E.; De Aquino, A.A.; Santos, A.D.C.M.; Queiroz, A.L.N.; Rodrigues, D.S.G.; et al. Development of RT-qPCR and Semi-Nested RT-PCR Assays for Molecular Diagnosis of Hantavirus Pulmonary Syndrome. PLoS Negl. Trop. Dis. 2019, 13, e0007884. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, M.; Schmidt, P.; Vackova, M.; Krivanec, K.; Munclinger, P.; Hufert, F.T. Identification of Genetic Evidence for Dobrava Virus Spillover in Rodents by Nested Reverse Transcription (RT)-PCR and TaqMan RT-PCR. J. Clin. Microbiol. 2005, 43, 808–812. [Google Scholar] [CrossRef]
- Niskanen, S.; Jääskeläinen, A.; Vapalahti, O.; Sironen, T. Evaluation of Real-Time RT-PCR for Diagnostic Use in Detection of Puumala Virus. Viruses 2019, 11, 661. [Google Scholar] [CrossRef]
- Aitichou, M.; Saleh, S.S.; McElroy, A.K.; Schmaljohn, C.; Ibrahim, M.S. Identification of Dobrava, Hantaan, Seoul, and Puumala Viruses by One-Step Real-Time RT-PCR. J. Virol. Methods 2005, 124, 21–26. [Google Scholar] [CrossRef]
- Pang, Z.; Li, A.; Li, J.; Qu, J.; He, C.; Zhang, S.; Li, C.; Zhang, Q.; Liang, M.; Li, D. Comprehensive Multiplex One-Step Real-Time TaqMan qRT-PCR Assays for Detection and Quantification of Hemorrhagic Fever Viruses. PLoS ONE 2014, 9, e95635. [Google Scholar] [CrossRef]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Denys, C.; Koivogui, L.; ter Meulen, J.; Krüger, D.H. Hantavirus in African Wood Mouse, Guinea. Emerg. Infect. Dis. 2006, 12, 838–840. [Google Scholar] [CrossRef]
- Erdin, M.; Stanoeva, K.R.; Mögling, R.; Korva, M.; Knap, N.; Resman Rus, K.; Domingo, C.; Reimerink, J.H.; De Vries, A.; Alburkat, H.; et al. External Quality Assessment of Orthohantavirus and Lymphocytic Choriomeningitis Virus Molecular Detection and Serology in Europe, 2021. Eurosurveillance 2023, 28, 2300054. [Google Scholar] [CrossRef]
- Bagamian, K.H.; Towner, J.S.; Kuenzi, A.J.; Douglass, R.J.; Rollin, P.E.; Waller, L.A.; Mills, J.N. Transmission Ecology of Sin Nombre Hantavirus in Naturally Infected North American Deermouse Populations in Outdoor Enclosures. PLoS ONE 2012, 7, e47731. [Google Scholar] [CrossRef]
- Smith, C.S.; Underwood, D.J.; Gordon, A.; Pyne, M.J.; Smyth, A.; Genge, B.; Driver, L.; Mayer, D.G.; Oakey, J. Identification and Epidemiological Analysis of a Putative Novel Hantavirus in Australian Flying Foxes. Virus Genes 2025, 61, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Z.; Zhang, F.-X.; Gao, N.; Wang, J.-B.; Zhao, Z.-W.; Li, M.-H.; Chen, H.-X.; Zou, Y.; Plyusnin, A. Hantaviruses in Rodents and Humans, Inner Mongolia Autonomous Region, China. Emerg. Infect. Dis. 2009, 15, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Plyusnin, A.; Hörling, J.; Kanerva, M.; Mustonen, J.; Cheng, Y.; Partanen, J.; Vapalahti, O.; Kukkonen, S.K.; Niemimaa, J.; Henttonen, H.; et al. Puumala Hantavirus Genome in Patients with Nephropathia Epidemica: Correlation of PCR Positivity with HLA Haplotype and Link to Viral Sequences in Local Rodents. J. Clin. Microbiol. 1997, 35, 1090–1096. [Google Scholar] [CrossRef]
- Puthavathana, P.; Ho, W.L.; Yong Kang, C. Typing of Hantaviruses from Five Continents by Polymerase Chain Reaction. Virus Res. 1992, 26, 1–14. [Google Scholar] [CrossRef]
- Nordström, H.; Johansson, P.; Li, Q.; Lundkvist, Å.; Nilsson, P.; Elgh, F. Microarray Technology for Identification and Distinction of Hantaviruses. J. Med. Virol. 2004, 72, 646–655. [Google Scholar] [CrossRef]
- Filippone, C.; Castel, G.; Murri, S.; Ermonval, M.; Korva, M.; Avšič-Županc, T.; Sironen, T.; Vapalahati, O.; McElhinney, L.M.; Ulrich, R.G.; et al. Revisiting the Genetic Diversity of Emerging Hantaviruses Circulating in Europe Using a Pan-Viral Resequencing Microarray. Sci. Rep. 2019, 9, 12404. [Google Scholar] [CrossRef]
- Oude Munnink, B.B.; Münger, E.; Nieuwenhuijse, D.F.; Kohl, R.; Van Der Linden, A.; Schapendonk, C.M.E.; Van Der Jeugd, H.; Kik, M.; Rijks, J.M.; Reusken, C.B.E.M.; et al. Genomic Monitoring to Understand the Emergence and Spread of Usutu Virus in the Netherlands, 2016–2018. Sci. Rep. 2020, 10, 2798. [Google Scholar] [CrossRef]
- Gire, S.K.; Goba, A.; Andersen, K.G.; Sealfon, R.S.G.; Park, D.J.; Kanneh, L.; Jalloh, S.; Momoh, M.; Fullah, M.; Dudas, G.; et al. Genomic Surveillance Elucidates Ebola Virus Origin and Transmission during the 2014 Outbreak. Science 2014, 345, 1369–1372. [Google Scholar] [CrossRef]
- Kim, W.-K.; Cho, S.; Lee, S.-H.; No, J.S.; Lee, G.-Y.; Park, K.; Lee, D.; Jeong, S.T.; Song, J.-W. Genomic Epidemiology and Active Surveillance to Investigate Outbreaks of Hantaviruses. Front. Cell. Infect. Microbiol. 2021, 10, 532388. [Google Scholar] [CrossRef]
- Klein, T.A.; Kim, H.-C.; Chong, S.-T.; Kim, J.-A.; Lee, S.-Y.; Kim, W.-K.; Nunn, P.V.; Song, J.-W. Hantaan Virus Surveillance Targeting Small Mammals at Nightmare Range, a High Elevation Military Training Area, Gyeonggi Province, Republic of Korea. PLoS ONE 2015, 10, e0118483. [Google Scholar] [CrossRef]
- Hjelle, B.; Torrez-Martinez, N.; Koster, F.T.; Jay, M.; Ascher, M.S.; Brown, T.; Reynolds, P.; Ettestad, P.; Voorhees, R.E.; Sarisky, J.; et al. Epidemiologic Linkage of Rodent and Human Hantavirus Genomic Sequences in Case Investigations of Hantavirus Pulmonary Syndrome. J. Infect. Dis. 1996, 173, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Schilling, S.; Emmerich, P.; Klempa, B.; Auste, B.; Schnaith, E.; Schmitz, H.; Krüger, D.H.; Günther, S.; Meisel, H. Hantavirus Disease Outbreak in Germany: Limitations of Routine Serological Diagnostics and Clustering of Virus Sequences of Human and Rodent Origin. J. Clin. Microbiol. 2007, 45, 3008–3014. [Google Scholar] [CrossRef] [PubMed]
- Quer, J.; Colomer-Castell, S.; Campos, C.; Andrés, C.; Piñana, M.; Cortese, M.F.; González-Sánchez, A.; Garcia-Cehic, D.; Ibáñez, M.; Pumarola, T.; et al. Next-Generation Sequencing for Confronting Virus Pandemics. Viruses 2022, 14, 600. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.W.; Matthews, D.A.; Hiscox, J.A.; Elmore, M.J.; Pollakis, G.; Rambaut, A.; Hewson, R.; García-Dorival, I.; Bore, J.A.; Koundouno, R.; et al. Temporal and Spatial Analysis of the 2014–2015 Ebola Virus Outbreak in West Africa. Nature 2015, 524, 97–101. [Google Scholar] [CrossRef]
- Siddle, K.J.; Eromon, P.; Barnes, K.G.; Mehta, S.; Oguzie, J.U.; Odia, I.; Schaffner, S.F.; Winnicki, S.M.; Shah, R.R.; Qu, J.; et al. Genomic Analysis of Lassa Virus during an Increase in Cases in Nigeria in 2018. N. Engl. J. Med. 2018, 379, 1745–1753. [Google Scholar] [CrossRef]
- Grubaugh, N.D.; Ladner, J.T.; Kraemer, M.U.G.; Dudas, G.; Tan, A.L.; Gangavarapu, K.; Wiley, M.R.; White, S.; Thézé, J.; Magnani, D.M.; et al. Genomic Epidemiology Reveals Multiple Introductions of Zika Virus into the United States. Nature 2017, 546, 401–405. [Google Scholar] [CrossRef]
- No, J.S.; Kim, W.-K.; Cho, S.; Lee, S.-H.; Kim, J.-A.; Lee, D.; Song, D.H.; Gu, S.H.; Jeong, S.T.; Wiley, M.R.; et al. Comparison of Targeted Next-Generation Sequencing for Whole-Genome Sequencing of Hantaan Orthohantavirus in Apodemus Agrarius Lung Tissues. Sci. Rep. 2019, 9, 16631. [Google Scholar] [CrossRef]
- McMullan, L.K.; Albariño, C.G.; Ksiazek, T.G.; Nichol, S.T.; Spiropoulou, C.F. Complete Genome Sequences of a Hantavirus Isolate from New York. Genome Announc. 2018, 6, e00188-18. [Google Scholar] [CrossRef]
- Cho, S.; Kim, W.-K.; No, J.S.; Lee, S.-H.; Jung, J.; Yi, Y.; Park, H.C.; Lee, G.-Y.; Park, K.; Kim, J.-A.; et al. Urinary Genome Detection and Tracking of Hantaan Virus from Hemorrhagic Fever with Renal Syndrome Patients Using Multiplex PCR-Based next-Generation Sequencing. PLoS Negl. Trop. Dis. 2021, 15, e0009707. [Google Scholar] [CrossRef]
- Pérez-Umphrey, A.A.; Settlecowski, A.E.; Elbers, J.P.; Williams, S.T.; Jonsson, C.B.; Bonisoli-Alquati, A.; Snider, A.M.; Taylor, S.S. Genetic Variants Associated with Hantavirus Infection in a Reservoir Host Are Related to Regulation of Inflammation and Immune Surveillance. Infect. Genet. Evol. 2023, 116, 105525. [Google Scholar] [CrossRef]
- Taylor, M.K.; Williams, E.P.; Wongsurawat, T.; Jenjaroenpun, P.; Nookaew, I.; Jonsson, C.B. Amplicon-Based, Next-Generation Sequencing Approaches to Characterize Single Nucleotide Polymorphisms of Orthohantavirus Species. Front. Cell. Infect. Microbiol. 2020, 10, 565591. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Lee, S.-H.; Kim, J.; Lee, J.; Lee, G.-Y.; Cho, S.; Lee, S.H.; Park, K.; No, J.S.; Budhathoki, S.; et al. Multiplex PCR-Based Nanopore Sequencing and Epidemiological Surveillance of Hantaan Orthohantavirus in Apodemus Agrarius, Republic of Korea. Viruses 2021, 13, 847. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, D.; Slenczka, W.; Alter, P.; Boehm, S.; Feldmann, H. Hemorrhagic Fever with Renal Syndrome: Diagnostic Problems with a Known Disease. J. Clin. Microbiol. 2001, 39, 3414–3416. [Google Scholar] [CrossRef] [PubMed]
- Galeno, H.; Mora, J.; Villagra, E.; Fernandez, J.; Hernandez, J.; Mertz, G.J.; Ramirez, E. First Human Isolate of Hantavirus (Andes Virus) in the Americas. Emerg. Infect. Dis. 2002, 8, 657–661. [Google Scholar] [CrossRef]
- Kruger, D.H.; Figueiredo, L.T.M.; Song, J.-W.; Klempa, B. Hantaviruses—Globally Emerging Pathogens. J. Clin. Virol. 2015, 64, 128–136. [Google Scholar] [CrossRef]
- Hantavirus Outbreak Toolbox. Available online: https://www.who.int/emergencies/outbreak-toolkit/disease-outbreak-toolboxes/hantavirus-outbreak-toolbox (accessed on 1 October 2024).
- PAHO Strengthens Capacities for Hantavirus and Arenavirus Surveillance in the Americas. Available online: https://www.paho.org/en/news/13-3-2024-paho-strengthens-capacities-hantavirus-and-arenavirus-surveillance-americas#:~:text=This%20international%20collaboration%20will%20not,fight%20against%20emerging%20and%20re%2D (accessed on 13 March 2024).
- Knust, B.; Rollin, P.E. Twenty-Year Summary of Surveillance for Human Hantavirus Infections, United States. Emerg. Infect. Dis. 2013, 19, 1934–1937. [Google Scholar] [CrossRef]
- Hantaviruses (Infection with). Available online: https://www.woah.org/app/uploads/2022/02/hantaviruses-infection-with.pdf (accessed on 1 February 2022).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Specificity | Sensitivity | Sample Size | Reference Assay | Biological Matrix | Target Gene | Virus(es) Detected | Test Type | TEST |
|---|---|---|---|---|---|---|---|---|---|
| [91] | NA | NA | 3 | Rapid peroxidase enzyme-linked immunosorbent assay (PAGEIA) | Blood samples | Small (S) and medium (M) segments | SNV | Homemade one-step RT-PCR nested method | SNV-MH strains 1, 2, and 3 RT-PCR |
| [91] | NA | 95–103% | 3 | Rapid peroxidase enzyme-linked immunosorbent assay (PAGEIA) | Blood samples | Small (S) segment | SNV | qRT–PCR with fluorescent signal | SNV Quantitative Real time RT-PCR |
| [92] | No cross-reactivity with Hendra virus genotype 1, Hendra virus genotype 2, ABLV, Kunjin virus, Murray Valley encephalitis virus, Ross River virus, or Japanese encephalitis virus | NA | NA | NA | Brain tissue | Small (S) segment | ROBV | qRT–PCR with fluorescent signal | Quantitative Real time RT-PCR |
| [92] | NA | NA | NA | NA | Brain tissue | Small (S) segment | ROBV | Homemade one-step RT-PCR nested method | nested RT-PCR |
| [81] | 10 copies | NA | NA | NA | DOBV, PUUV, TULV and HTNV, SEOV, and ANDV and SNV | Homemade five-different-qRT-PCR method | 5 real-time RT-PCR assays | ||
| [93] | NA | NA | NA | NA | Lung tissues | Small (S) segment | HNTV and SEOV | Homemade one-step RT-PCR nested method | nested RT-PCR |
| [94] | NA | NA | 20 | Immunoblotting | Peripheral blood mononuclear cells and lung tissues | Entire S segment and partial M segment | PUUV | Homemade one-step RT-PCR nested method followed by Sanger sequencing | nested RT-PCR |
| [86] | DOBV, SAAV, HTNV, TULV, and TOPV | 3.3 copies per reaction | 288 | Immunofluorescence assay | Serum samples | Small (S) segment | PUUV | Homemade real-time RT-PCR method modified from Plyusnin A et al.’s one-step RT-PCR method | real-time RT-PCR and nested RT-PCR |
| [91] | NA | NA | 7 | Rapid peroxidase enzyme-linked immunosorbent assay (PAGEIA) | Blood samples from deer mice | Partial S segment and partial M segment | SNV | Homemade one-step RT-PCR nested method and homemade real-time RT-PCR method | real-time RT-PCR and nested RT-PCR |
| [12] | NA | NA | 612 | NA | Blood samples | Highly conserved L segment | Members of the genus Orthohantavirus | Homemade nested RT-PCR assay | nested RT-PCR |
| [95] | Partial S segment | Members of the genus Orthohantavirus | Five different RT-PCRs analysed via restriction endonuclease digestion | RT-PCR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romeo, M.A.; Tofani, S.; Lapa, D.; Mija, C.; Maggi, F.; Scicluna, M.T.; Nardini, R. Orthohantaviruses: An Overview of the Current Status of Diagnostics and Surveillance. Viruses 2025, 17, 622. https://doi.org/10.3390/v17050622
Romeo MA, Tofani S, Lapa D, Mija C, Maggi F, Scicluna MT, Nardini R. Orthohantaviruses: An Overview of the Current Status of Diagnostics and Surveillance. Viruses. 2025; 17(5):622. https://doi.org/10.3390/v17050622
Chicago/Turabian StyleRomeo, Maria Anele, Silvia Tofani, Daniele Lapa, Cosmina Mija, Fabrizio Maggi, Maria Teresa Scicluna, and Roberto Nardini. 2025. "Orthohantaviruses: An Overview of the Current Status of Diagnostics and Surveillance" Viruses 17, no. 5: 622. https://doi.org/10.3390/v17050622
APA StyleRomeo, M. A., Tofani, S., Lapa, D., Mija, C., Maggi, F., Scicluna, M. T., & Nardini, R. (2025). Orthohantaviruses: An Overview of the Current Status of Diagnostics and Surveillance. Viruses, 17(5), 622. https://doi.org/10.3390/v17050622