

Expression Profiles of lncRNAs and mRNAs in the Mouse Brain Infected with Pseudorabies Virus: A Bioinformatic Analysis


Abstract
1. Introduction
2. Materials and Methods
2.1. Mice, Cell Lines, and Virus
2.2. Animal Inoculation
2.3. DNA Extraction and Viral Copy Number Determination
2.4. Total RNA Extraction
2.5. Library Construction and Sequencing
2.6. Quality Control
2.7. Read Mapping to the Reference Genome
2.8. Gene Expression Level Calculation
2.9. Differential Expression Analysis
2.10. GO Functional Enrichment and KEGG Pathway Enrichment Analysis
2.11. lncRNA Target Gene Prediction and Functional Analysis
2.12. Protein–Protein Interaction (PPI) Network Construction and Module Analysis
2.13. Quantitative Real-Time PCR (qPCR)
2.14. Statistical Analysis
3. Results
3.1. Validation of PRV Infection in Mouse Brains
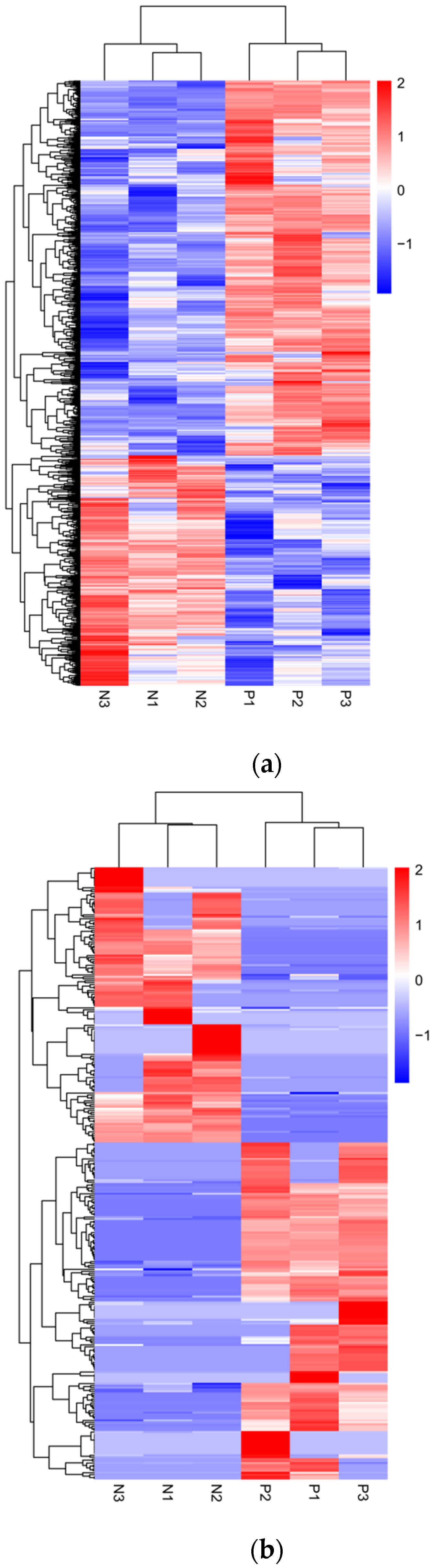
3.2. Analysis of Differential Expression Profiles of lncRNAs and mRNAs
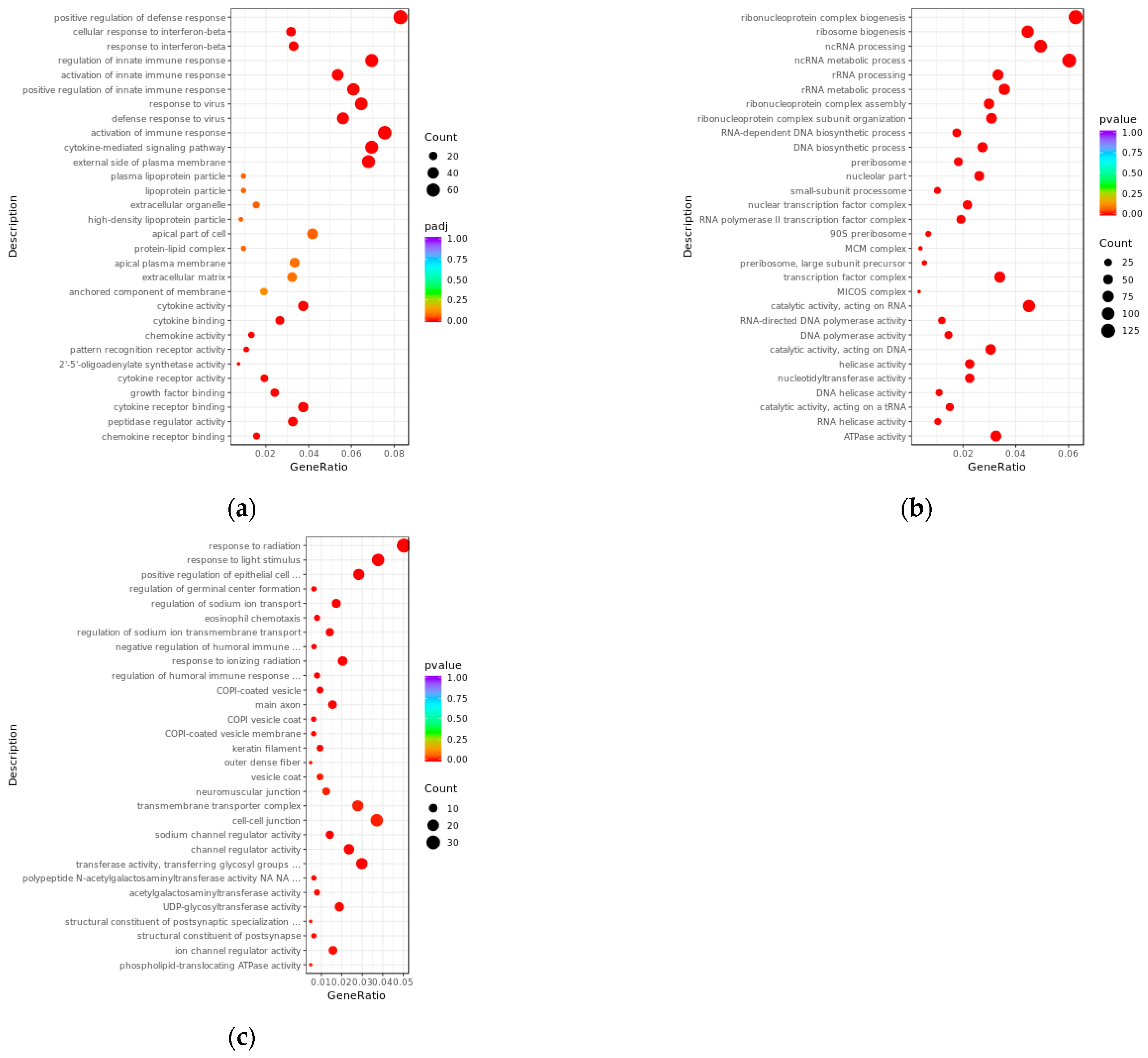
3.3. GO Enrichment Analysis of Differentially Expressed lncRNAs and mRNAs
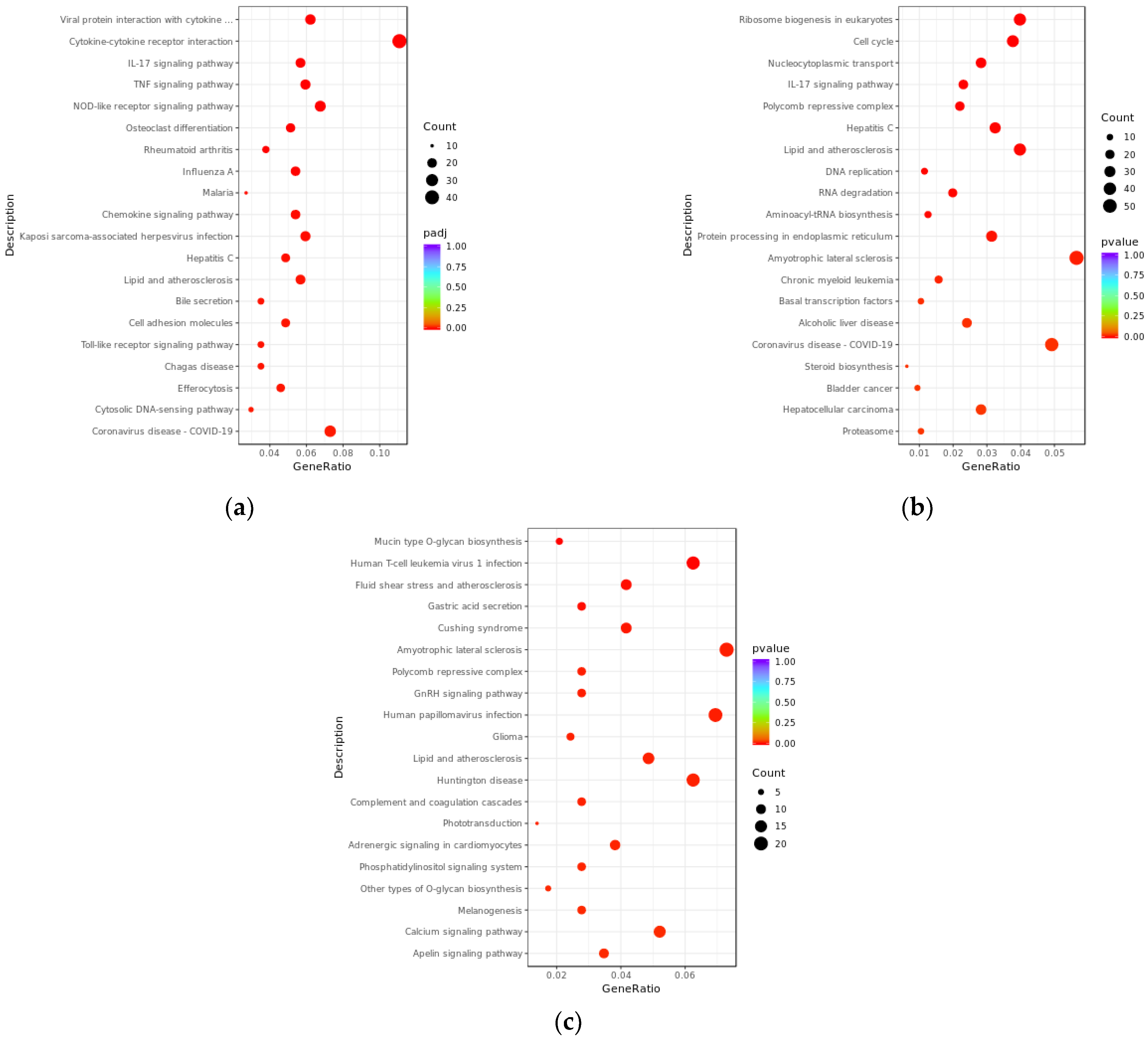
3.4. KEGG Enrichment Analysis of Differentially Expressed lncRNAs and mRNAs
3.5. Construction and Module Analysis of Protein–Protein Interaction (PPI) Network
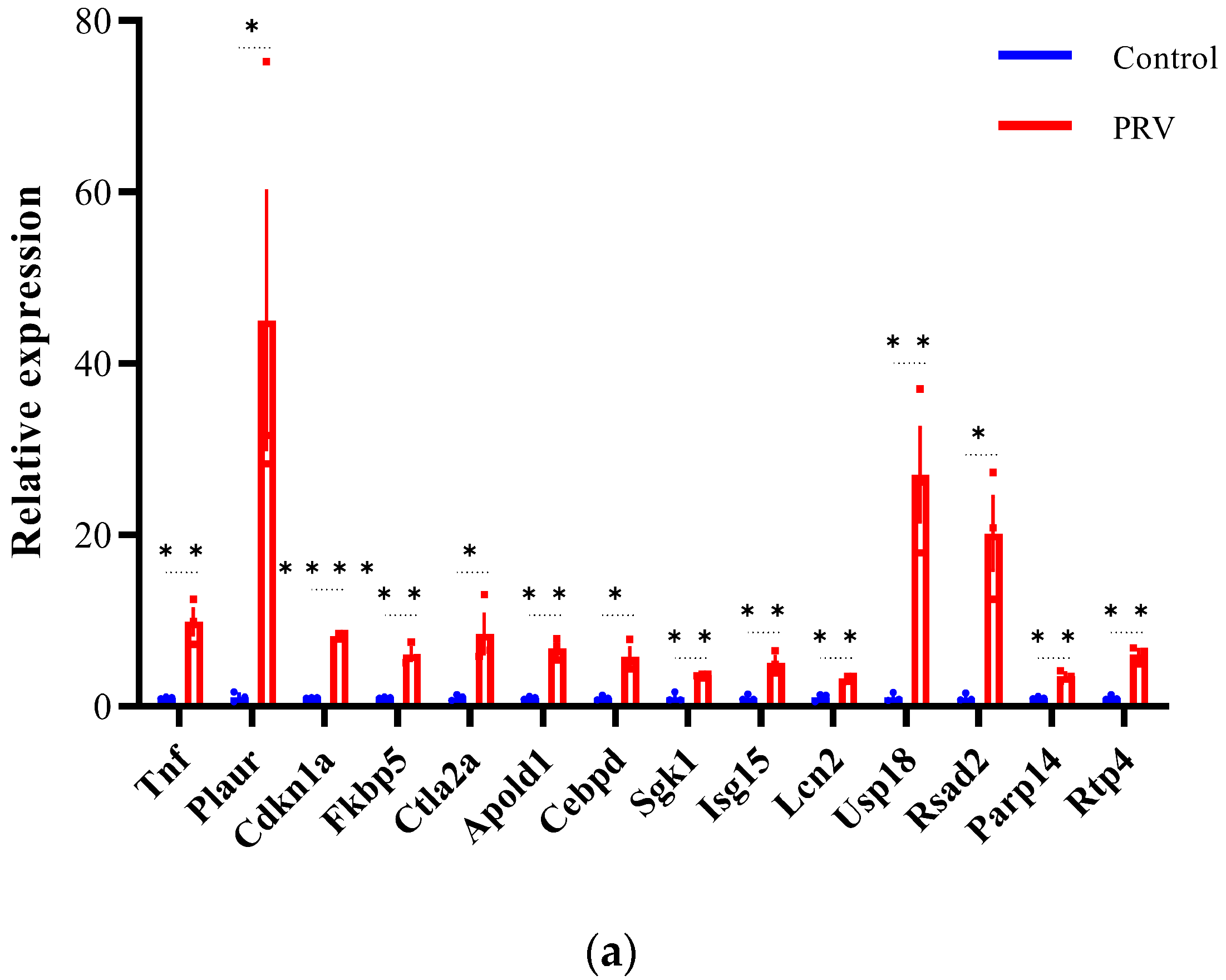
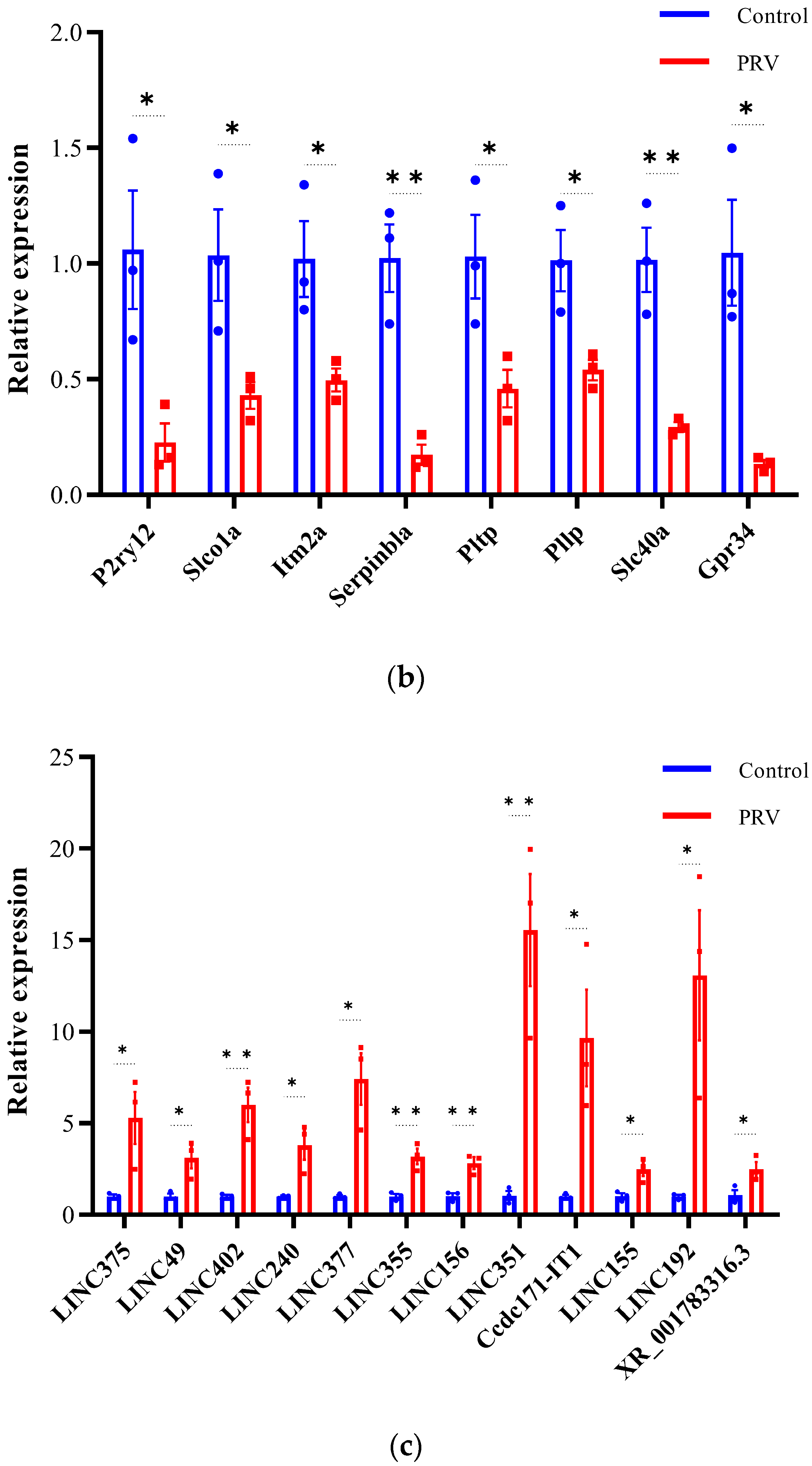
3.6. Validation of Differentially Expressed lncRNAs and mRNAs by qRT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zheng, H.H.; Fu, P.F.; Chen, H.Y.; Wang, Z.Y. Pseudorabies Virus: From Pathogenesis to Prevention Strategies. Viruses 2022, 14, 1638. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, K.; Stoikou, A.; Papadopoulos, D.K.; Tsapouri-Kanoula, E.; Giantsis, I.A.; Papadopoulos, D.; Stamelou, E.; Sofia, M.; Billinis, C.; Karapetsiou, C.; et al. Pseudorabies Virus Prevalence in Lung Samples of Hunted Wild Boars in Northwestern Greece. Pathogens 2024, 13, 929. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, G.; Wan, C.; Li, Y.; Peng, L.; Fang, R.; Peng, Y.; Ye, C. A Comparison of Pseudorabies Virus Latency to Other alpha-Herpesvirinae Subfamily Members. Viruses 2022, 14, 1386. [Google Scholar] [CrossRef]
- Klopfleisch, R.; Klupp, B.G.; Fuchs, W.; Kopp, M.; Teifke, J.P.; Mettenleiter, T.C. Influence of pseudorabies virus proteins on neuroinvasion and neurovirulence in mice. J. Virol. 2006, 80, 5571–5576. [Google Scholar] [CrossRef]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef]
- Brittle, E.E.; Reynolds, A.E.; Enquist, L.W. Two modes of pseudorabies virus neuroinvasion and lethality in mice. J. Virol. 2004, 78, 12951–12963. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A Novel Human Acute Encephalitis Caused by Pseudorabies Virus Variant Strain. Clin. Infect. Dis. 2021, 73, e3690–e3700. [Google Scholar] [CrossRef]
- Yang, H.; Han, H.; Wang, H.; Cui, Y.; Liu, H.; Ding, S. A Case of Human Viral Encephalitis Caused by Pseudorabies Virus Infection in China. Front. Neurol. 2019, 10, 534. [Google Scholar] [CrossRef]
- Wang, Y.; Nian, H.; Li, Z.; Wang, W.; Wang, X.; Cui, Y. Human encephalitis complicated with bilateral acute retinal necrosis associated with pseudorabies virus infection: A case report. Int. J. Infect. Dis. 2019, 89, 51–54. [Google Scholar] [CrossRef]
- Elrashedy, A.; Mousa, W.; Nayel, M.; Salama, A.; Zaghawa, A.; Elsify, A.; Hasan, M.E. Advances in bioinformatics and multi-omics integration: Transforming viral infectious disease research in veterinary medicine. Virol. J. 2025, 22, 22. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Agliano, F.; Rathinam, V.A.; Medvedev, A.E.; Vanaja, S.K.; Vella, A.T. Long Noncoding RNAs in Host-Pathogen Interactions. Trends Immunol. 2019, 40, 492–510. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Gao, Y.; Liu, X.; Bai, J.; Jiang, P.; Wang, X. Long non-coding RNA LNC_000641 regulates pseudorabies virus replication. Vet. Res. 2021, 52, 52. [Google Scholar] [CrossRef] [PubMed]
- Van Solingen, C.; Cyr, Y.; Scacalossi, K.R.; de Vries, M.; Barrett, T.J.; de Jong, A.; Gourvest, M.; Zhang, T.; Peled, D.; Kher, R.; et al. Long noncoding RNA CHROMR regulates antiviral immunity in humans. Proc. Natl. Acad. Sci. USA 2022, 119, e2210321119. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, J.; Hu, Y.; Chai, Y.; Song, J.; Duan, J.; Zhang, S.; Xu, X. Virus-induced lncRNA-BTX allows viral replication by regulating intracellular translocation of DHX9 and ILF3 to induce innate escape. Cell Rep. 2023, 42, 113262. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, W.; Huang, L.; Xiao, J.; Song, X.; Li, F.; Ma, Y.; Wang, X.; Jin, F.; Liu, P.; et al. A novel lncRNA linc-AhRA negatively regulates innate antiviral response in murine microglia upon neurotropic herpesvirus infection. Theranostics 2021, 11, 9623–9651. [Google Scholar] [CrossRef]
- Sui, B.; Chen, D.; Liu, W.; Wu, Q.; Tian, B.; Li, Y.; Hou, J.; Liu, S.; Xie, J.; Jiang, H.; et al. A novel antiviral lncRNA, EDAL, shields a T309 O-GlcNAcylation site to promote EZH2 lysosomal degradation. Genome Biol. 2020, 21, 228. [Google Scholar] [CrossRef]
- Basavappa, M.G.; Ferretti, M.; Dittmar, M.; Stoute, J.; Sullivan, M.C.; Whig, K.; Shen, H.; Liu, K.F.; Schultz, D.C.; Beiting, D.P.; et al. The lncRNA ALPHA specifically targets chikungunya virus to control infection. Mol. Cell 2022, 82, 3729–3744.e3710. [Google Scholar] [CrossRef]
- Nie, Z.; Zhu, S.; Wu, L.; Sun, R.; Shu, J.; He, Y.; Feng, H. Progress on innate immune evasion and live attenuated vaccine of pseudorabies virus. Front. Microbiol. 2023, 14, 1138016. [Google Scholar] [CrossRef]
- Bo, Z.; Li, X. A Review of Pseudorabies Virus Variants: Genomics, Vaccination, Transmission, and Zoonotic Potential. Viruses 2022, 14, 1003. [Google Scholar] [CrossRef]
- Sehl, J.; Holper, J.E.; Klupp, B.G.; Baumbach, C.; Teifke, J.P.; Mettenleiter, T.C. An improved animal model for herpesvirus encephalitis in humans. PLoS Pathog. 2020, 16, e1008445. [Google Scholar] [CrossRef] [PubMed]
- Ryu, V.; Gumerova, A.A.; Witztum, R.; Korkmaz, F.; Cullen, L.; Kannangara, H.; Moldavski, O.; Barak, O.; Lizneva, D.; Goosens, K.A.; et al. An atlas of brain-bone sympathetic neural circuits in mice. eLife 2024, 13, e95727. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Diamond, M.S. Innate immune interactions within the central nervous system modulate pathogenesis of viral infections. Curr. Opin. Immunol. 2015, 36, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Lampron, A.; Elali, A.; Rivest, S. Innate immunity in the CNS: Redefining the relationship between the CNS and Its environment. Neuron 2013, 78, 214–232. [Google Scholar] [CrossRef]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef]
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- Forrester, J.V.; McMenamin, P.G.; Dando, S.J. CNS infection and immune privilege. Nat. Rev. Neurosci. 2018, 19, 655–671. [Google Scholar] [CrossRef]
- Dhondt, K.P.; Mathieu, C.; Chalons, M.; Reynaud, J.M.; Vallve, A.; Raoul, H.; Horvat, B. Type I interferon signaling protects mice from lethal henipavirus infection. J. Infect. Dis. 2013, 207, 142–151. [Google Scholar] [CrossRef]
- Kochs, G.; Bauer, S.; Vogt, C.; Frenz, T.; Tschopp, J.; Kalinke, U.; Waibler, Z. Thogoto virus infection induces sustained type I interferon responses that depend on RIG-I-like helicase signaling of conventional dendritic cells. J. Virol. 2010, 84, 12344–12350. [Google Scholar] [CrossRef]
- Sancho-Shimizu, V.; Perez de Diego, R.; Lorenzo, L.; Halwani, R.; Alangari, A.; Israelsson, E.; Fabrega, S.; Cardon, A.; Maluenda, J.; Tatematsu, M.; et al. Herpes simplex encephalitis in children with autosomal recessive and dominant TRIF deficiency. J. Clin. Investig. 2011, 121, 4889–4902. [Google Scholar] [CrossRef]
- Perez de Diego, R.; Sancho-Shimizu, V.; Lorenzo, L.; Puel, A.; Plancoulaine, S.; Picard, C.; Herman, M.; Cardon, A.; Durandy, A.; Bustamante, J.; et al. Human TRAF3 adaptor molecule deficiency leads to impaired Toll-like receptor 3 response and susceptibility to herpes simplex encephalitis. Immunity 2010, 33, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Herman, M.; Ciancanelli, M.; Ou, Y.H.; Lorenzo, L.; Klaudel-Dreszler, M.; Pauwels, E.; Sancho-Shimizu, V.; Perez de Diego, R.; Abhyankar, A.; Israelsson, E.; et al. Heterozygous TBK1 mutations impair TLR3 immunity and underlie herpes simplex encephalitis of childhood. J. Exp. Med. 2012, 209, 1567–1582. [Google Scholar] [CrossRef] [PubMed]
- Casrouge, A.; Zhang, S.Y.; Eidenschenk, C.; Jouanguy, E.; Puel, A.; Yang, K.; Alcais, A.; Picard, C.; Mahfoufi, N.; Nicolas, N.; et al. Herpes simplex virus encephalitis in human UNC-93B deficiency. Science 2006, 314, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.; Bergmann, C.C. Alpha/Beta Interferon (IFN-alpha/beta) Signaling in Astrocytes Mediates Protection against Viral Encephalomyelitis and Regulates IFN-gamma-Dependent Responses. J. Virol. 2018, 92, e01901-17. [Google Scholar] [CrossRef]
- Pfefferkorn, C.; Kallfass, C.; Lienenklaus, S.; Spanier, J.; Kalinke, U.; Rieder, M.; Conzelmann, K.K.; Michiels, T.; Staeheli, P. Abortively Infected Astrocytes Appear To Represent the Main Source of Interferon Beta in the Virus-Infected Brain. J. Virol. 2016, 90, 2031–2038. [Google Scholar] [CrossRef]
- Kallfass, C.; Ackerman, A.; Lienenklaus, S.; Weiss, S.; Heimrich, B.; Staeheli, P. Visualizing production of beta interferon by astrocytes and microglia in brain of La Crosse virus-infected mice. J. Virol. 2012, 86, 11223–11230. [Google Scholar] [CrossRef]
- Telikani, Z.; Monson, E.A.; Hofer, M.J.; Helbig, K.J. Antiviral response within different cell types of the CNS. Front. Immunol. 2022, 13, 1044721. [Google Scholar] [CrossRef]
- Perng, Y.C.; Lenschow, D.J. ISG15 in antiviral immunity and beyond. Nat. Rev. Microbiol. 2018, 16, 423–439. [Google Scholar] [CrossRef]
- Freitas, B.T.; Scholte, F.E.M.; Bergeron, E.; Pegan, S.D. How ISG15 combats viral infection. Virus Res. 2020, 286, 198036. [Google Scholar] [CrossRef]
- Liu, H.; Li, C.; He, W.; Chen, J.; Yang, G.; Chen, L.; Chang, H. Free ISG15 inhibits Pseudorabies virus infection by positively regulating type I IFN signaling. PLoS Pathog. 2022, 18, e1010921. [Google Scholar] [CrossRef]
- Das Sarma, J.; Burrows, A.; Rayman, P.; Hwang, M.H.; Kundu, S.; Sharma, N.; Bergmann, C.; Sen, G.C. Ifit2 deficiency restricts microglial activation and leukocyte migration following murine coronavirus (m-CoV) CNS infection. PLoS Pathog. 2020, 16, e1009034. [Google Scholar] [CrossRef] [PubMed]
- Carty, M.; Reinert, L.; Paludan, S.R.; Bowie, A.G. Innate antiviral signalling in the central nervous system. Trends Immunol. 2014, 35, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Harschnitz, O.; Studer, L.; Casanova, J.L. Neuron-intrinsic immunity to viruses in mice and humans. Curr. Opin. Immunol. 2021, 72, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Audry, M.; Ciancanelli, M.; Alsina, L.; Azevedo, J.; Herman, M.; Anguiano, E.; Sancho-Shimizu, V.; Lorenzo, L.; Pauwels, E.; et al. Herpes simplex virus encephalitis in a patient with complete TLR3 deficiency: TLR3 is otherwise redundant in protective immunity. J. Exp. Med. 2011, 208, 2083–2098. [Google Scholar] [CrossRef]
- Berankova, M.; Holoubek, J.; Honig, V.; Matusova, Z.; Palus, M.; Salat, J.; Krayem, I.; Vojtiskova, J.; Svoboda, P.; Pranclova, V.; et al. Genotype-driven sensitivity of mice to tick-borne encephalitis virus correlates with differential host responses in peripheral macrophages and brain. J. Neuroinflammation 2025, 22, 22. [Google Scholar] [CrossRef]
- Lim, S.M.; van den Ham, H.J.; Oduber, M.; Martina, E.; Zaaraoui-Boutahar, F.; Roose, J.M.; van Ijcken, W.F.J.; Osterhaus, A.; Andeweg, A.C.; Koraka, P.; et al. Transcriptomic Analyses Reveal Differential Gene Expression of Immune and Cell Death Pathways in the Brains of Mice Infected with West Nile Virus and Chikungunya Virus. Front. Microbiol. 2017, 8, 1556. [Google Scholar] [CrossRef]
- Clarke, P.; Leser, J.S.; Bowen, R.A.; Tyler, K.L. Virus-induced transcriptional changes in the brain include the differential expression of genes associated with interferon, apoptosis, interleukin 17 receptor A, and glutamate signaling as well as flavivirus-specific upregulation of tRNA synthetases. mBio 2014, 5, e00902–e00914. [Google Scholar] [CrossRef]
- Pasieka, T.J.; Cilloniz, C.; Carter, V.S.; Rosato, P.; Katze, M.G.; Leib, D.A. Functional genomics reveals an essential and specific role for Stat1 in protection of the central nervous system following herpes simplex virus corneal infection. J. Virol. 2011, 85, 12972–12981. [Google Scholar] [CrossRef]
- Appolinario, C.M.; Daly, J.M.; Emes, R.D.; Marchi, F.A.; Ribeiro, B.L.D.; Megid, J. Gene Expression Profile Induced by Two Different Variants of Street Rabies Virus in Mice. Viruses 2022, 14, 692. [Google Scholar] [CrossRef]
- Van Loo, G.; Bertrand, M.J.M. Death by TNF: A road to inflammation. Nat. Rev. Immunol. 2023, 23, 289–303. [Google Scholar] [CrossRef]
- Yeh, C.J.; Lin, P.Y.; Liao, M.H.; Liu, H.J.; Lee, J.W.; Chiu, S.J.; Hsu, H.Y.; Shih, W.L. TNF-alpha mediates pseudorabies virus-induced apoptosis via the activation of p38 MAPK and JNK/SAPK signaling. Virology 2008, 381, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Rybak-Wolf, A.; Wyler, E.; Pentimalli, T.M.; Legnini, I.; Oliveras Martinez, A.; Glazar, P.; Loewa, A.; Kim, S.J.; Kaufer, B.B.; Woehler, A.; et al. Modelling viral encephalitis caused by herpes simplex virus 1 infection in cerebral organoids. Nat. Microbiol. 2023, 8, 1252–1266. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, Y.; Tao, Q.; Xu, T.; Lee, F.-q.; Deng, L.-s.; Jian, Z.; Zhao, J.; Yang, Y.; Lai, S.; et al. Transcriptome and metabolome analysis reveals PRV XJ delgE/gI/TK protects intracranially infected mice from death by regulating the inflammation. Front. Microbiol. 2024, 15, 1374646. [Google Scholar] [CrossRef]
- Ekstrand, M.I.; Enquist, L.W.; Pomeranz, L.E. The alpha-herpesviruses: Molecular pathfinders in nervous system circuits. Trends Mol. Med. 2008, 14, 134–140. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Hussen, B.M.; Jamal, H.H.; Taheri, M.; Sharifi, G. The Emerging Role of Non-Coding RNAs in the Regulation of Virus Replication and Resultant Cellular Pathologies. Int. J. Mol. Sci. 2022, 23, 815. [Google Scholar] [CrossRef]
- Zhang, Q.; Martin-Caraballo, M.; Hsia, S.V. Modulation of Voltage-Gated Sodium Channel Activity in Human Dorsal Root Ganglion Neurons by Herpesvirus Quiescent Infection. J. Virol. 2020, 94, e01823-19. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Chan, M.T.V.; Wu, W.K.K. Bioinformatic analyses suggest augmented interleukin-17 signaling as the mechanism of COVID-19-associated herpes zoster. Environ. Sci. Pollut. Res. Int. 2021, 28, 65769–65775. [Google Scholar] [CrossRef]
- Sun, X.; Jin, X.; Lin, Z.; Liu, X.; Yang, J.; Li, L.; Feng, H.; Zhang, W.; Gu, C.; Hu, X.; et al. Nucleotide-binding oligomerization domain 1 (NOD1) regulates microglial activation in pseudorabies virus infection. Vet. Res. 2024, 55, 161. [Google Scholar] [CrossRef]
- Sun, Q.; Hao, Q.; Prasanth, K.V. Nuclear Long Noncoding RNAs: Key Regulators of Gene Expression. Trends Genet. 2018, 34, 142–157. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, K.; Huang, W.; Tang, W.; Li, H.; Dong, W.; Gu, J.; Zhou, J. Identification of functional lncRNAs in pseudorabies virus type II infected cells. Vet. Microbiol. 2020, 242, 108564. [Google Scholar] [CrossRef]
- Tayel, S.I.; El-Masry, E.A.; Abdelaal, G.A.; Shehab-Eldeen, S.; Essa, A.; Muharram, N.M. Interplay of LncRNAs NEAT1 and TUG1 in Incidence of Cytokine Storm in Appraisal of COVID-19 Infection. Int. J. Biol. Sci. 2022, 18, 4901–4913. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Han, P.; Ye, W.; Chen, H.; Zheng, X.; Cheng, L.; Zhang, L.; Yu, L.; Wu, X.; Xu, Z.; et al. The Long Noncoding RNA NEAT1 Exerts Antihantaviral Effects by Acting as Positive Feedback for RIG-I Signaling. J. Virol. 2017, 91, e02250-16. [Google Scholar] [CrossRef] [PubMed]
- Morchikh, M.; Cribier, A.; Raffel, R.; Amraoui, S.; Cau, J.; Severac, D.; Dubois, E.; Schwartz, O.; Bennasser, Y.; Benkirane, M. HEXIM1 and NEAT1 Long Non-coding RNA Form a Multi-subunit Complex that Regulates DNA-Mediated Innate Immune Response. Mol. Cell 2017, 67, 387–399.e5. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fan, P.; Zhao, Y.; Zhang, S.; Lu, J.; Xie, W.; Jiang, Y.; Lei, F.; Xu, N.; Zhang, Y. NEAT1 modulates herpes simplex virus-1 replication by regulating viral gene transcription. Cell. Mol. Life Sci. 2017, 74, 1117–1131. [Google Scholar] [CrossRef]
- Chen, S.; Huang, X.; Xie, Q.; Liu, Q.; Zhu, H. The Role of Long Noncoding RNA BST2-2 in the Innate Immune Response to Viral Infection. J. Virol. 2022, 96, e0020722. [Google Scholar] [CrossRef]
- Tripathi, S.; Sengar, S.; Shree, B.; Mohapatra, S.; Basu, A.; Sharma, V. An RBM10 and NF-κB interacting host lncRNA promotes JEV replication and neuronal cell death. J. Virol. 2023, 97, e0118323. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′-3′) |
| LINC375-F | 5′ CTGCCCTAATGTTATCCC 3′ |
| LINC375-R | 5′ GCCATCTTACAGCCACC 3′ |
| Msl2-IT2-F | 5′ AGTGACTGCCATCTTCT 3′ |
| Msl2-IT2-R | 5′ CACAACCCTGTAACCTA 3′ |
| LINC49-F | 5′ GTTCTGTCGCTTGGTAAT 3′ |
| LINC49-R | 5′ CTGGAGTTGGAGTGGG 3′ |
| Mest-OT9-F | 5′ ATGAATCTACCTCGTGC 3′ |
| Mest-OT9-R | 5′ CTCTTCTCCTAACTCCCT 3′ |
| Msl2-IT1-F | 5′ AGTGACTGCCATCTTCT 3′ |
| Msl2-IT1-R | 5′ CACAACCCTGTAACCTA 3′ |
| LINC402-F | 5′ GAACCCTCCCAACTAAA 3′ |
| LINC402-R | 5′ TCTGCCAAAGGAATCTA 3′ |
| LINC240-F | 5′ CCTCACCCTAAAGTTCG 3′ |
| LINC240-R | 5′ CTGGGCAGGGAAAGTA 3′ |
| LINC377-F | 5′ CTGCCCTAATGTTATCCC 3′ |
| LINC377-R | 5′ GCCATCTTACAGCCACC 3′ |
| LINC355-F | 5′ TGGTTCTGTCGCTTGG 3′ |
| LINC355-R | 5′ CTGGAGTTGGAGTGGG 3′ |
| LINC156-F | 5′ GCCTCCTGGAACAACC 3′ |
| LINC156-R | 5′ AACTGGGCAGGGAAAG 3′ |
| NR_166531.1-F | 5′ CCCAAACCAAATCCAC 3′ |
| NR_166531.1-R | 5′ ACAATCACCTGGCACAT 3′ |
| LINC351-F | 5′ TTCCCAGAACACTCCTAA 3′ |
| LINC351-R | 5′ AGTCTCGACCGTCAGC 3′ |
| LINC155-F | 5′ GCCTCCTGGAACAACC 3′ |
| LINC155-R | 5′ AACTGGGCAGGGAAAG 3′ |
| XR_004939660.1-F | 5′ GTTACGGAAAGCATCTC 3′ |
| XR_004939660.1-R | 5′ CAGAGCAAGTGGGTCA 3′ |
| XR_387385.5-F | 5′ ATCCGCCTTCTTGTGC 3′ |
| XR_387385.5-R | 5′ TGTTTGATGGTGGTGTCG 3′ |
| Ccdc171-IT1-F | 5′ GAACCCTCCCAACTAAA 3′ |
| Ccdc171-IT1-R | 5′ TCTGCCAAAGGAATCTA 3′ |
| XR_387382.1-F | 5′ ATCCGCCTTCTTGTGC 3′ |
| XR_387382.1-R | 5′ TGTTTGATGGTGGTGTCG 3′ |
| XR_381404.4-F | 5′ GGATGTGATGTGGAGGTT 3′ |
| XR_381404.4-R | 5′ GCATTCAGTGGCTTCTATT 3′ |
| LINC192-F | 5′ CCTCACCCTAAAGTTCAA 3′ |
| LINC192-R | 5′ AAGGAGTACCTGGTTCAT 3′ |
| XR_001783316.3-F | 5′ ATCTGTATCAGTTAGGGTTC 3′ |
| XR_001783316.3-R | 5′ TTGCCAGCACATCTTT 3′ |
| Plaur-F | 5′ CGGGAATGGCAAGATGA 3′ |
| Plaur-R | 5′ TCTGGTCCAAAGAGGTGC 3′ |
| Cdkn1a-F | 5′ TTCCGCACAGGAGCAAA 3′ |
| Cdkn1a-R | 5′ AAGTCAAAGTTCCACCGTTCT 3′ |
| Fkbp5-F | 5′ ATGAGGGCACCAGTAACAA 3′ |
| Fkbp5-R | 5′ CCAAGGCTAAAGGCAAAT 3′ |
| Ctla2a-F | 5′ TGAGCAGGGCAAGACC 3′ |
| Ctla2a-R | 5′ TCAGGCAAATCAGGAG 3′ |
| Apold1-F | 5′ GTGCGGAGGGTGCAGGAGAT 3′ |
| Apold1-R | 5′ AGCCGAAGAAGACGATGAAGTAGA 3′ |
| Cebpd-F | 5′ CCTGCCATGTACGACGACGAG 3′ |
| Cebpd-R | 5′ GCCGCTTTGTGGTTGCTGTT 3′ |
| Sgk1-F | 5′ GGGCACATCGTCCTCACT 3′ |
| Sgk1-R | 5′ CGGTCATACGGCTGCTTA 3′ |
| Isg15-F | 5′ CGATTTCCTGGTGTCCGTGA 3′ |
| Isg15-R | 5′ CTCGCTGCAGTTCTGTACCA 3′ |
| Usp18-F | 5′ CGTGCCGTTGTTTGTC 3′ |
| Usp18-R | 5′ GGCTTTGCGTCCTTATC 3′ |
| Rsad2-F | 5′ CCCTCTGTGAGCATAGTGA 3′ |
| Rsad2-R | 5′ GCCACCTTGTAATCCCT 3′ |
| Parp14-F | 5′ AGATTGGTCGTCAGTTCG 3′ |
| Parp14-R | 5′ GCTATGTCCCTCTGTAGGT 3′ |
| Rtp4-F | 5′ CCCCGATGACTTCAGTAC 3′ |
| Rtp4-R | 5′ CCTGAGCAGAGGTCCAAC 3′ |
| Tnf-qF | 5′ CTCATTCCTGCTTGTGGC 3′ |
| Tnf-qR | 5′ CACTTGGTGGTTTGCTACG 3′ |
| P2ry12-qF | 5′ GCTTGGCAATGAGGAT 3′ |
| P2ry12-qR | 5′ GGTAGCGGTCAATGGTT 3′ |
| Slco1a4-qF | 5′ GGTTGCCTGCTGCTCT 3′ |
| Slco1a4-qR | 5′ TTCCGTTCTCCATCATTCT 3′ |
| Itm2a-qF | 5′ GCCCAAGAGCACCATT 3′ |
| Itm2a-qR | 5′ TTCCCTTCTCAAAGTCGT 3′ |
| Serpinb1a-qF | 5′ ATACCCTCAACTCTAACCTG 3′ |
| Serpinb1a-qR | 5′ AATGTAGCAATGCCTCC 3′ |
| Pltp-qF | 5′ TCTGCCCTGTGCTCTACC 3′ |
| Pltp-qR | 5′ GGCTCCAGTTGTCCTCCTT 3′ |
| Pllp-qF | 5′ CGAAAGTGAGCACGAGGAC 3′ |
| Pllp-qR | 5′ CACCAGCCAGAGGAAGACA 3′ |
| Slc40a1-qF | 5′ TTGGCTTGCTGGTATTG 3′ |
| Slc40a1-qR | 5′ AACAGATGTATTCGGTTGAT 3′ |
| Gpr34-qF | 5′ CGTGGGACTGGTTGGA 3′ |
| Gpr34-qR | 5′ GGGAGGCAGAAGATGA 3′ |
| Lcn2-F | 5′ GCCCAGGACTCAACTCA 3′ |
| Lcn2-R | 5′ GCCCACAACGTACCACC 3′ |
| Actb-F | 5′ AATCGTGCGTGACATCAA 3′ |
| Actb-R | 5′ AGAAGGAAGGCTGGAAAA 3′ |
| PRV-EP0-F | 5′ CGGGCGAAGACAAACAAAGG 3′ |
| PRV-EP0-R | 5′ GGGCGGTAGAAGCCAAAGATC 3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Tu, T.; Luo, Y.; Yao, X.; Yang, Z.; Wang, Y. Expression Profiles of lncRNAs and mRNAs in the Mouse Brain Infected with Pseudorabies Virus: A Bioinformatic Analysis. Viruses 2025, 17, 580. https://doi.org/10.3390/v17040580
Li Y, Tu T, Luo Y, Yao X, Yang Z, Wang Y. Expression Profiles of lncRNAs and mRNAs in the Mouse Brain Infected with Pseudorabies Virus: A Bioinformatic Analysis. Viruses. 2025; 17(4):580. https://doi.org/10.3390/v17040580
Chicago/Turabian StyleLi, Yanwei, Teng Tu, Yan Luo, Xueping Yao, Zexiao Yang, and Yin Wang. 2025. "Expression Profiles of lncRNAs and mRNAs in the Mouse Brain Infected with Pseudorabies Virus: A Bioinformatic Analysis" Viruses 17, no. 4: 580. https://doi.org/10.3390/v17040580
APA StyleLi, Y., Tu, T., Luo, Y., Yao, X., Yang, Z., & Wang, Y. (2025). Expression Profiles of lncRNAs and mRNAs in the Mouse Brain Infected with Pseudorabies Virus: A Bioinformatic Analysis. Viruses, 17(4), 580. https://doi.org/10.3390/v17040580