
Cytokinins Reduce Viral Replication and Alter Plaque Morphology of Frog Virus 3 In Vitro


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
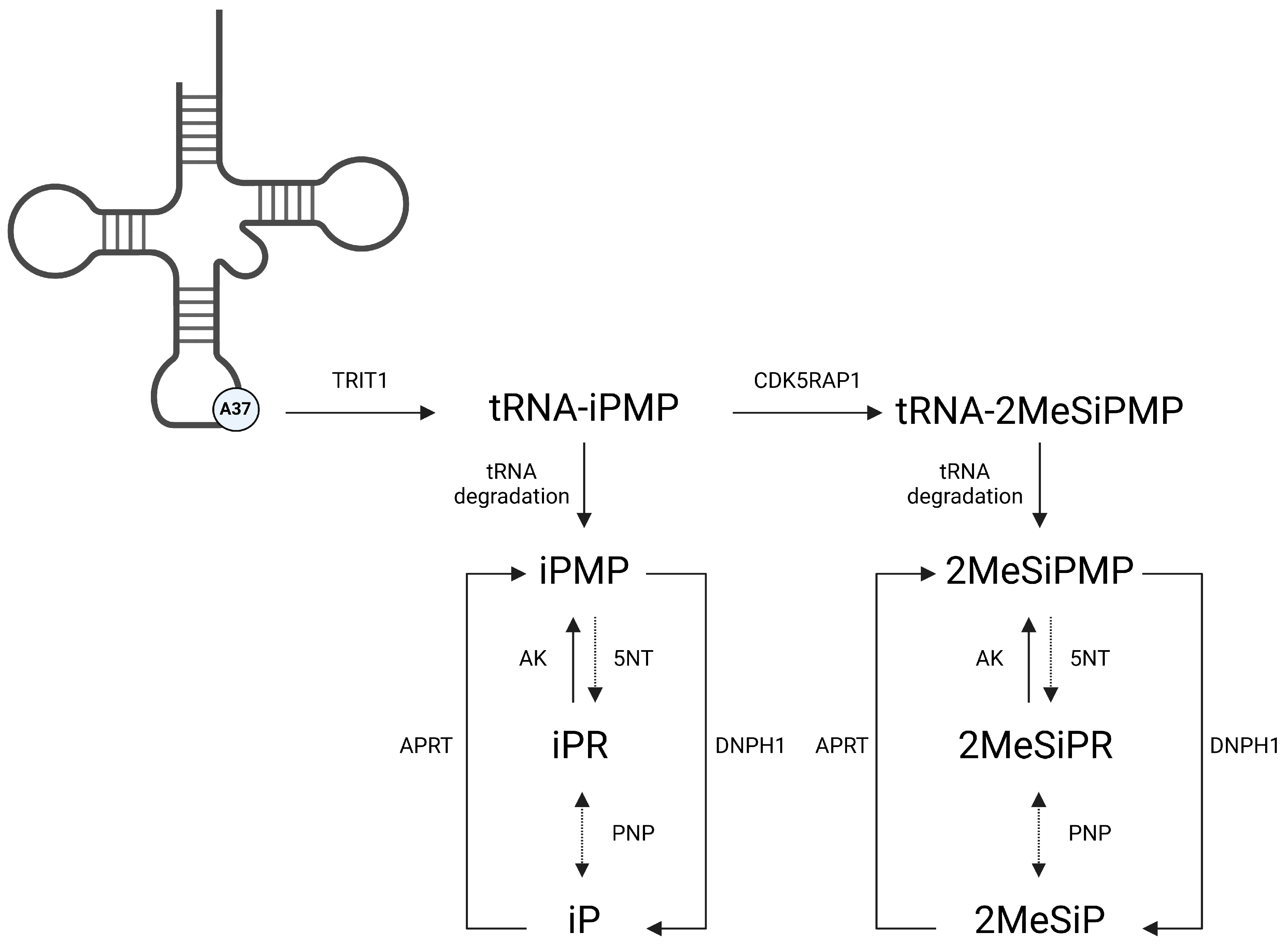
1. Introduction
2. Materials and Methods
2.1. Cytokinin Solution Preparation
2.2. Epithelioma Papulosum Cyprini (EPC) Cells
2.3. FV3 Propagation in EPC Cells
2.4. Plaque Formation and Morphology Assays
2.5. Single-Step Growth Curves
2.6. Cytotoxicity and Proliferation Assays
2.7. Statistical Analysis
3. Results
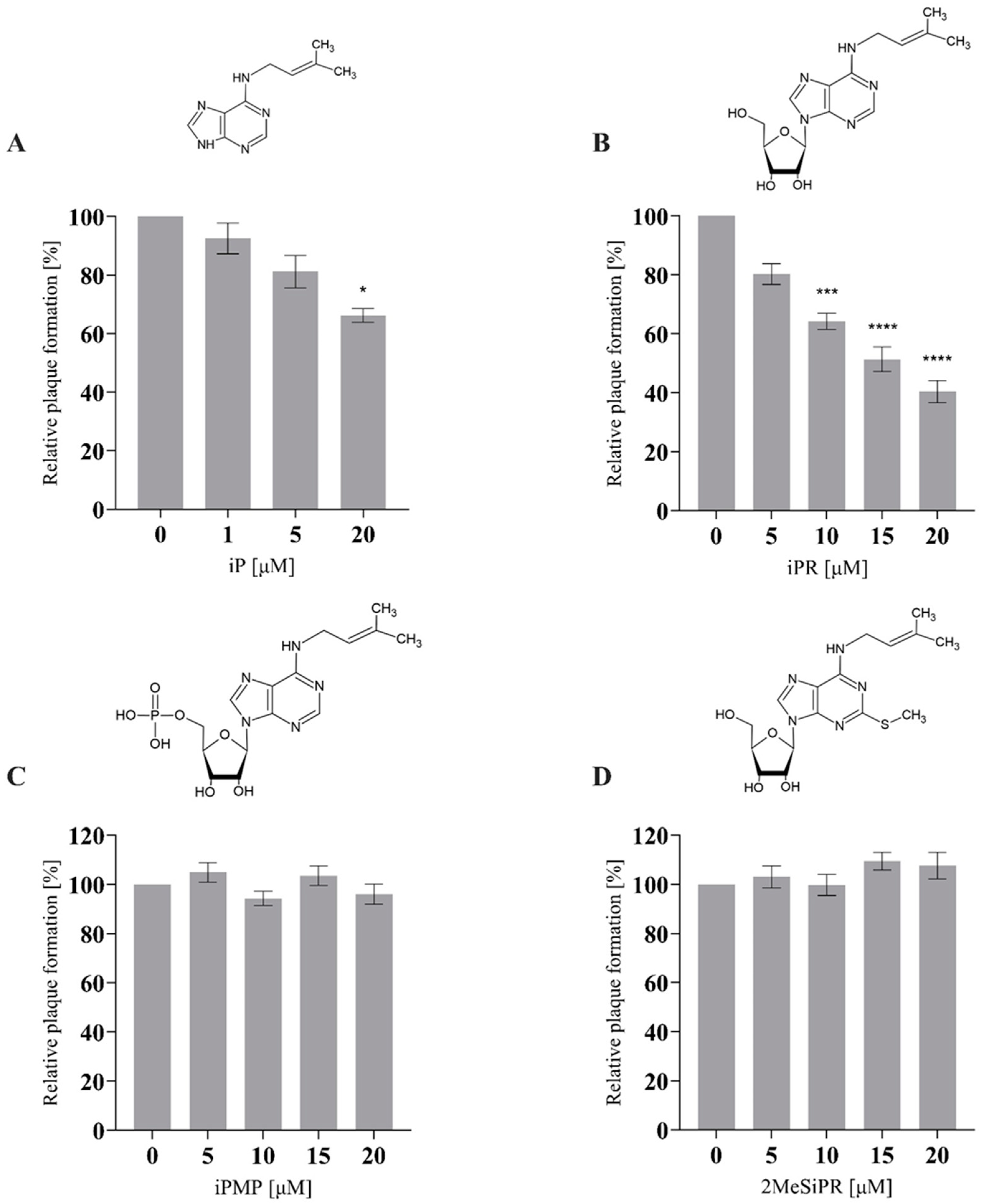
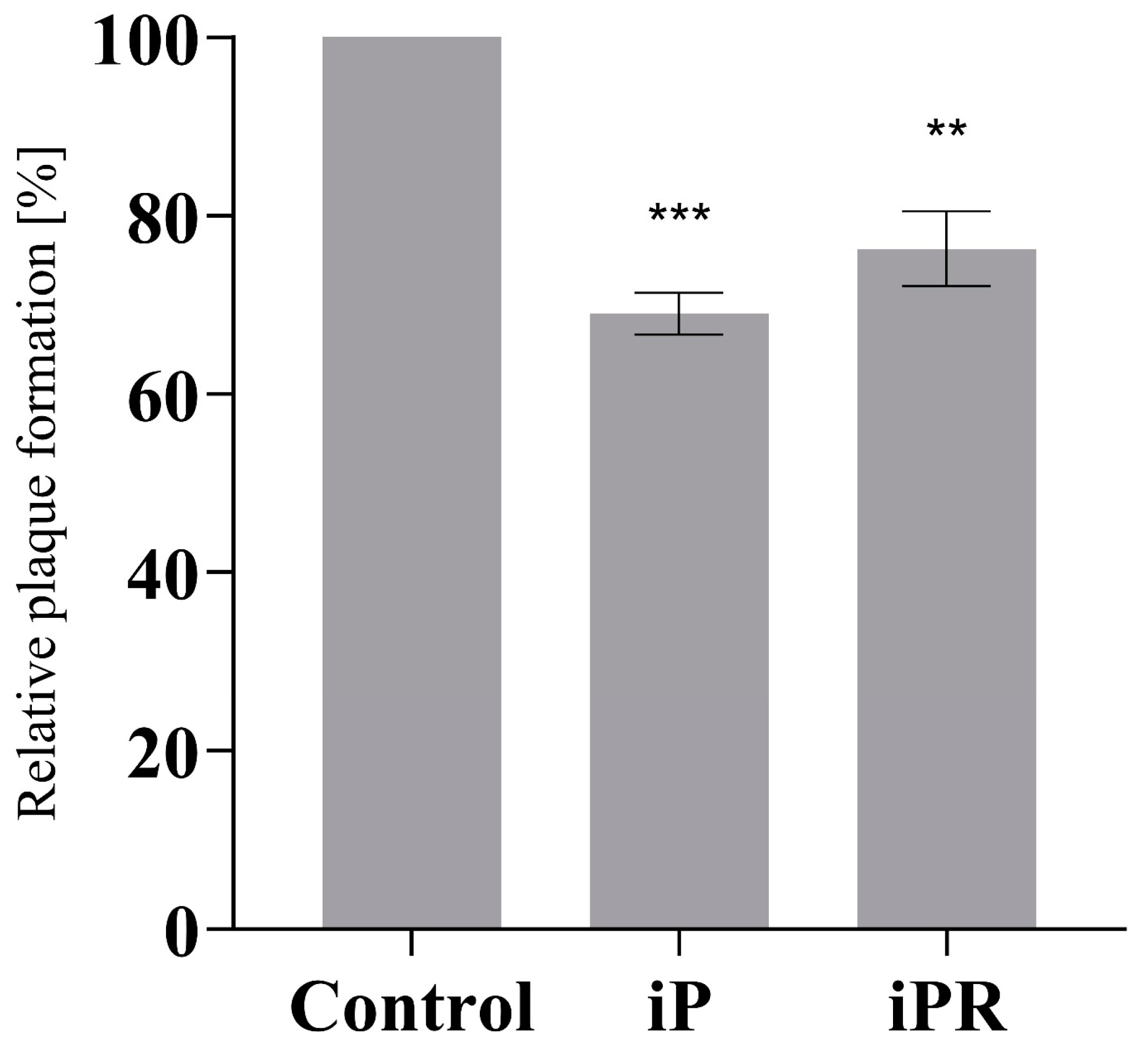
3.1. iP and iPR Reduce Relative Plaque Formation during FV3 Infection
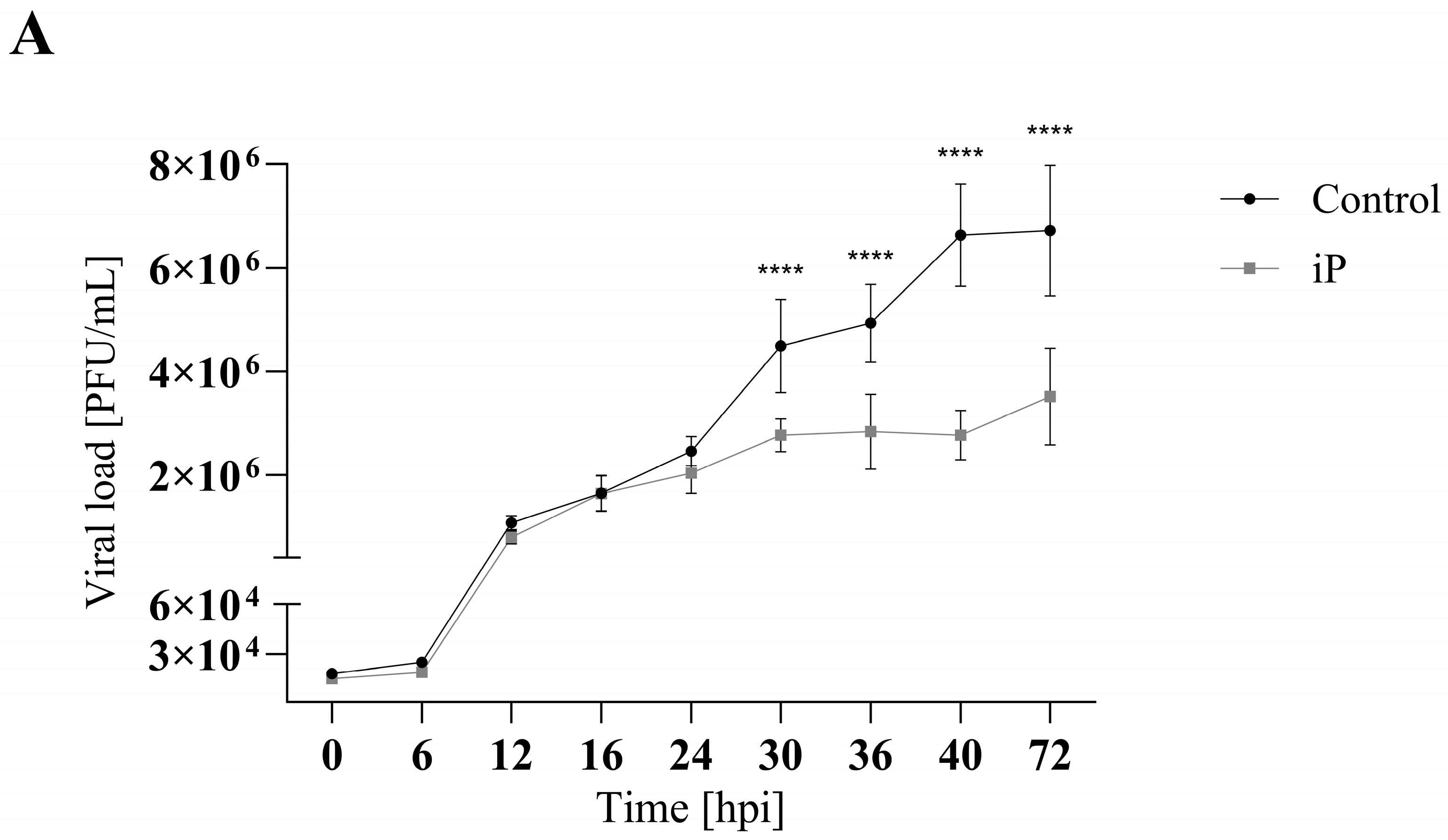
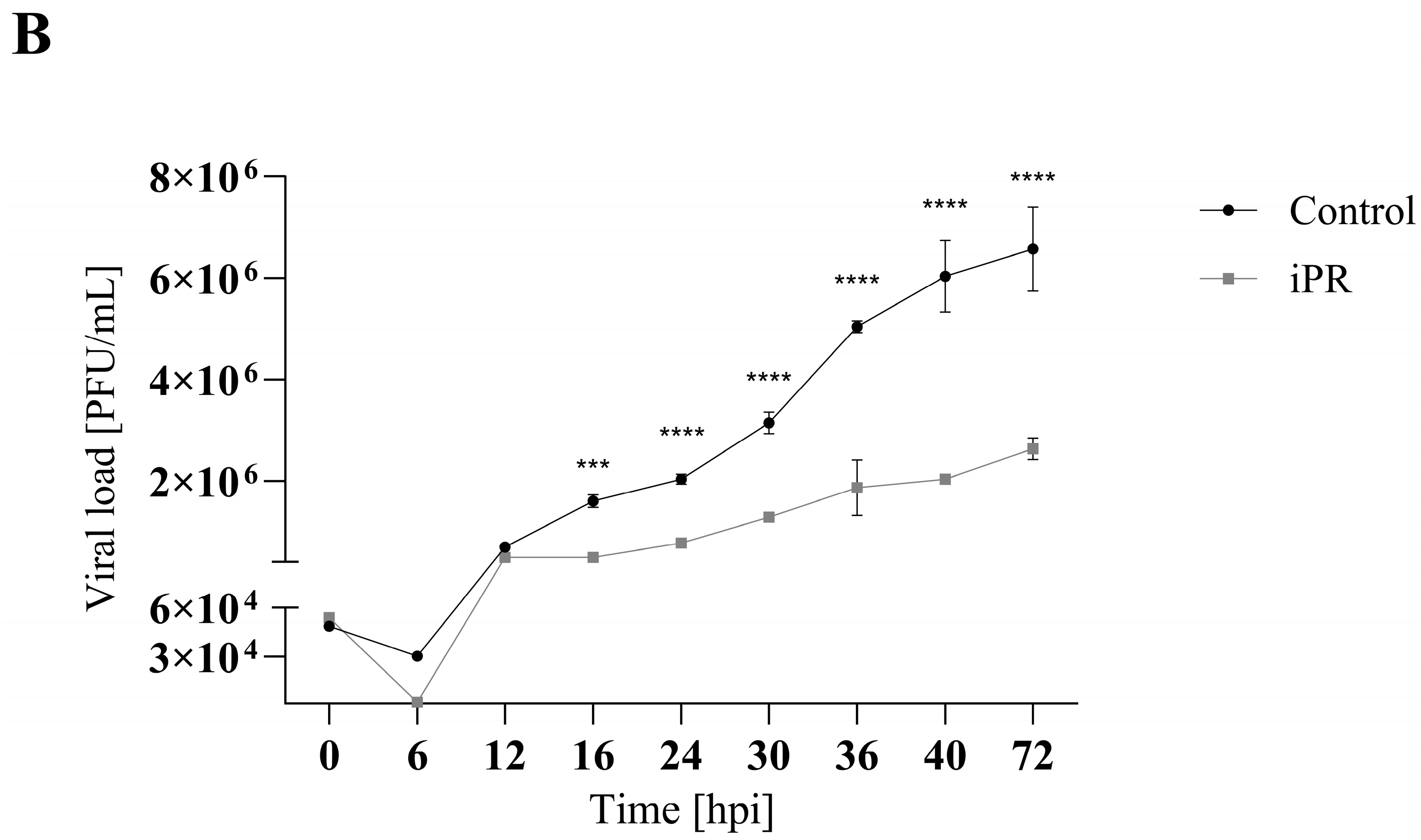
3.2. iP and iPR Begin to Significantly Reduce FV3 Titers after 30- and 16-h Post Infection, Respectively
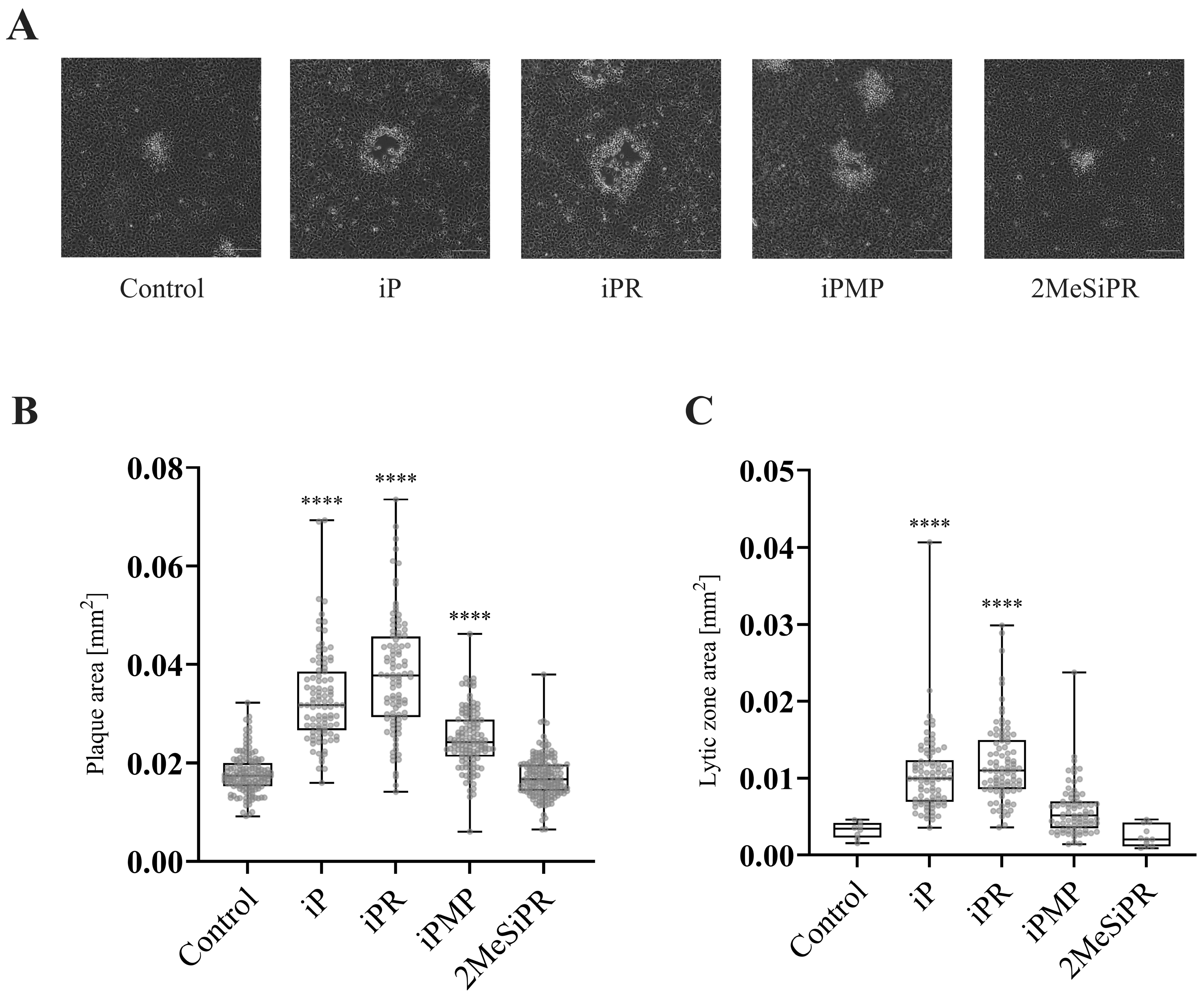
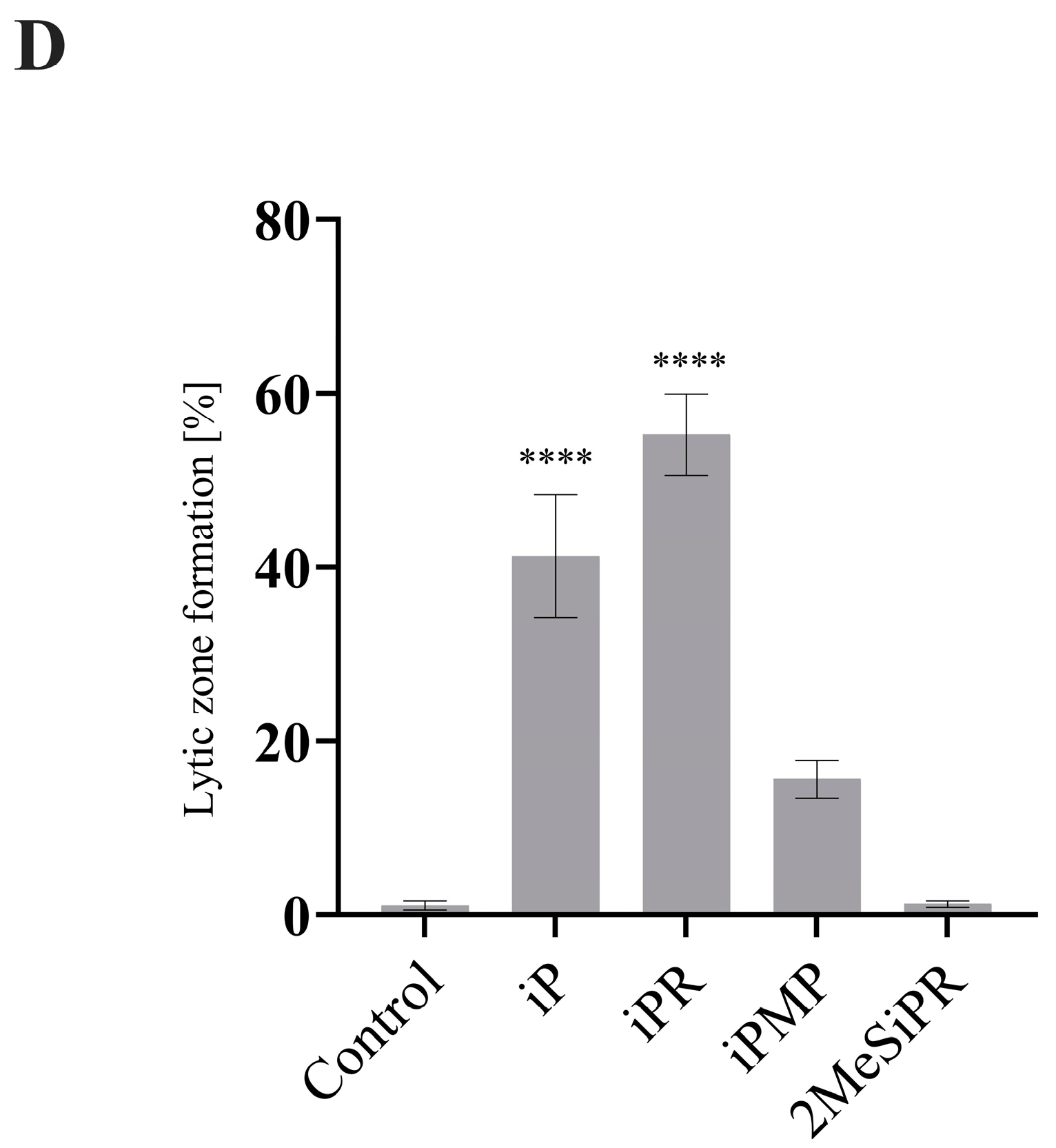
3.3. iP and iPR Alter FV3 Plaque Size and Morphology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spíchal, L. Cytokinins—Recent News and Views of Evolutionally Old Molecules. Funct. Plant Biol. 2012, 39, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Bidon, B.; Kabbara, S.; Courdavault, V.; Glévarec, G.; Oudin, A.; Héricourt, F.; Carpin, S.; Spíchal, L.; Binder, B.M.; Cock, J.M.; et al. Cytokinin and Ethylene Cell Signaling Pathways from Prokaryotes to Eukaryotes. Cells 2020, 9, 2526. [Google Scholar] [CrossRef] [PubMed]
- Kabbara, S.; Schmülling, T.; Papon, N. CHASEing Cytokinin Receptors in Plants, Bacteria, Fungi, and Beyond. Trends Plant Sci. 2018, 23, 179–181. [Google Scholar] [CrossRef]
- Mayaka, J.B.; Huang, Q.; Xiao, Y.; Zhong, Q.; Ni, J.; Shen, Y. The Lonely Guy (LOG) Homologue SiRe_0427 from the Thermophilic Archaeon Sulfolobus Islandicus REY15A Is a Phosphoribohydrolase Representing a Novel Group. Appl. Environ. Microbiol. 2019, 85, e01739-19. [Google Scholar] [CrossRef] [PubMed]
- Golovko, A.; Hjälm, G.; Sitbon, F.; Nicander, B. Cloning of a Human TRNA Isopentenyl Transferase. Gene 2000, 258, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Reiter, V.; Matschkal, D.M.S.; Wagner, M.; Globisch, D.; Kneuttinger, A.C.; Müller, M.; Carell, T.; Mu, M.; Carell, T. The CDK5 Repressor CDK5RAP1 Is a Methylthiotransferase Acting on Nuclear and Mitochondrial RNA. Nucleic Acids Res. 2012, 40, 6235–6240. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, L.; Li, C.; Zhou, J.; Li, Z. CDK5RAP1 Deficiency Induces Cell Cycle Arrest and Apoptosis in Human Breast Cancer Cell Line by the ROS/JNK Signaling Pathway. Oncol. Rep. 2015, 33, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.M.; Seegobin, M.; Kisiala, A.; Noble, A.; Brunetti, C.; Emery, R.J.N. Phytohormone Metabolism in Human Cells: Cytokinins Are Taken up and Interconverted in HeLa Cell Culture. FASEB bioAdvances 2019, 1, 320–331. [Google Scholar] [CrossRef]
- Seegobin, M.; Kisiala, A.; Noble, A.; Kaplan, D.; Brunetti, C.; Emery, R.J.N. Canis Familiaris Tissues Are Characterized by Different Profiles of Cytokinins Typical of the TRNA Degradation Pathway. FASEB J. 2018, 32, 6575–6581. [Google Scholar] [CrossRef]
- Aoki, M.M.; Kisiala, A.B.; Rahman, T.; Morrison, E.N.; Emery, R.J.N. Cytokinins Are Pervasive among Common in Vitro Culture Media: An Analysis of Their Forms, Concentrations and Potential Sources. J. Biotechnol. 2021, 334, 43–46. [Google Scholar] [CrossRef]
- Pisanti, S.; Picardi, P.; Ciaglia, E.; Margarucci, L.; Ronca, R.; Giacomini, A.; Malfitano, A.M.; Casapullo, A.; Laezza, C.; Gazzerro, P.; et al. Antiangiogenic Effects of N6-Isopentenyladenosine, an Endogenous Isoprenoid End Product, Mediated by AMPK Activation. FASEB J. 2014, 28, 1132–1144. [Google Scholar] [CrossRef]
- Rajabi, M.; Signorelli, P.; Gorincioi, E.; Ghidoni, R.; Santaniello, E. Antiproliferative Activity of N6-Isopentenyladenosine on MCF-7 Breast Cancer Cells: Cell Cycle Analysis and DNA-Binding Study. DNA Cell Biol. 2010, 29, 687–691. [Google Scholar] [CrossRef]
- Ishii, Y.; Hori, Y.; Sakai, S.; Honma, Y. Control of Differentiation and Apoptosis of Human Myeloid Leukemia Cells by Cytokinins and Cytokinin Nucleosides, Plant Redifferentiation-Inducing Hormones. Cell Growth Differ. 2002, 13, 19–26. [Google Scholar] [PubMed]
- Cabello, C.M.; III, W.B.B.; Ley, S.; Lamore, S.D.; Azimian, S.; Wondrak, G.T. The Experimental Chemotherapeutic N6-Furfuryladenosine (Kinetin-Riboside) Induces Rapid ATP Depletion, Genotoxic Stress, and CDKN1A (P21) Upregulation in Human Cancer Cell Lines. Biochem. Pharmacol. 2009, 77, 1125–1138. [Google Scholar] [CrossRef]
- Voller, J.; Béres, T.; Zatloukal, M.; Kaminski, P.A.; Niemann, P.; Doležal, K.; Džubák, P.; Hajdúch, M.; Strnad, M. The Natural Cytokinin 2OH3MeOBAR Induces Cell Death by a Mechanism That Is Different from That of the “Classical” Cytokinin Ribosides. Phytochemistry 2017, 136, 156–164. [Google Scholar] [CrossRef]
- Mlejnek, P.; Doležel, P. Apoptosis Induced by N6-Substituted Derivatives of Adenosine Is Related to Intracellular Accumulation of Corresponding Mononucleotides in HL-60 Cells. Toxicol. In Vitro 2005, 19, 985–990. [Google Scholar] [CrossRef]
- Ranieri, R.; Ciaglia, E.; Amodio, G.; Picardi, P.; Proto, M.C.; Gazzerro, P.; Laezza, C.; Remondelli, P.; Bifulco, M.; Pisanti, S. N6-Isopentenyladenosine Dual Targeting of AMPK and Rab7 Prenylation Inhibits Melanoma Growth through the Impairment of Autophagic Flux. Cell Death Differ. 2018, 25, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Laezza, C.; Malfitano, A.M.; Di Matola, T.; Ricchi, P.; Bifulco, M. Involvement of Akt/NF-ΚB Pathway in N6-Isopentenyladenosine-Induced Apoptosis in Human Breast Cancer Cells. Mol. Carcinog. 2010, 49, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Qi, Y.; Wei, J.; Lu, L.; Zhao, X.; Zhou, L. N6-Isopentenyladenosine Promoted HeLa Cell Apoptosis through Inhibitions of AKT and Transforming Growth Factor β–Activated Kinase 1 Activation. Tumor Biol. 2017, 39, 1010428317695966. [Google Scholar] [CrossRef]
- Laezza, C.; Caruso, M.G.; Gentile, T.; Notarnicola, M.; Malfitano, A.M.; Di Matola, T.; Messa, C.; Gazzerro, P.; Bifulco, M. N6-Isopentenyladenosine Inhibits Cell Proliferation and Induces Apoptosis in a Human Colon Cancer Cell Line DLD1. Int. J. Cancer 2009, 124, 1322–1329. [Google Scholar] [CrossRef]
- Laezza, C.; Notarnicola, M.; Caruso, M.G.; Messa, C.; Macchia, M.; Bertini, S.; Minutolo, F.; Portella, G.; Fiorentino, L.; Stingo, S.; et al. N6-Isopentenyladenosine Arrests Tumor Cell Proliferation by Inhibiting Farnesyl Diphosphate Synthase and Protein Prenylation. FASEB J. 2006, 20, 412–418. [Google Scholar] [CrossRef]
- Ciaglia, E.; Pisanti, S.; Picardi, P.; Laezza, C.; Sosa, S.; Tubaro, A.; Vitale, M.; Gazzerro, P.; Malfitano, A.M.; Bifulco, M. N6-Isopentenyladenosine Affects Cytotoxic Activity and Cytokines Production by IL-2 Activated NK Cells and Exerts Topical Anti-Inflammatory Activity in Mice. Pharmacol. Res. 2014, 89, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ciaglia, E.; Pisanti, S.; Picardi, P.; Laezza, C.; Malfitano, A.; D’Alessandro, A.; Gazzerro, P.; Vitale, M.; Carbone, E.; Bifulco, M. N6-Isopentenyladenosine, an Endogenous Isoprenoid End Product, Directly Affects Cytotoxic and Regulatory Functions of Human NK Cells through FDPS Modulation. J Leukoc Biol 2013, 94, 1207–1219. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Bioassay Record for AID 720532, Source: National Center for Advancing Translational Sciences (NCATS). Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/720532 (accessed on 5 May 2022).
- National Center for Biotechnology Information. PubChem Bioassay Record for AID 720533, Source: National Center for Advancing Translational Sciences (NCATS). Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/720533 (accessed on 5 May 2022).
- Tararov, V.I.; Tijsma, A.; Kolyachkina, S.V.; Oslovsky, V.E.; Neyts, J.; Drenichev, M.S.; Leyssen, P.; Mikhailov, S.N. Chemical Modification of the Plant Isoprenoid Cytokinin N6-Isopentenyladenosine Yields a Selective Inhibitor of Human Enterovirus 71 Replication. Eur. J. Med. Chem. 2015, 90, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Souza, T.M.L.; Pinho, V.D.; Setim, C.F.; Sacramento, C.Q.; Marcon, R.; Fintelman-Rodrigues, N.; Chaves, O.A.; Heller, M.; Temerozo, J.R.; Ferreira, A.C.; et al. Preclinical Development of Kinetin as a Safe Error-Prone SARS-CoV-2 Antiviral Able to Attenuate Virus-Induced Inflammation. Nat. Commun. 2023, 14, 199. [Google Scholar] [CrossRef]
- Orlov, A.A.; Drenichev, M.S.; Oslovsky, V.E.; Kurochkin, N.N.; Solyev, P.N.; Kozlovskaya, L.I.; Palyulin, V.A.; Karganova, G.G.; Mikhailov, S.N.; Osolodkin, D.I. New Tools in Nucleoside Toolbox of Tick-Borne Encephalitis Virus Reproduction Inhibitors. Bioorganic Med. Chem. Lett. 2017, 27, 1267–1273. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Yu, K.H.; Jancovich, J.K. The Molecular Biology of Frog Virus 3 and Other Iridoviruses Infecting Cold-Blooded Vertebrates. Viruses 2011, 3, 1959–1985. [Google Scholar] [CrossRef]
- Tian, Y.; Khwatenge, C.N.; Li, J.; De Jesus Andino, F.; Robert, J.; Sang, Y.; Cuesta, A.; Cho, K.; Jesus Andino, D.F. Targeted Transcriptomics of Frog Virus 3 in Infected Frog Tissues Reveal Non-Coding Regulatory Elements and MicroRNAs in the Ranaviral Genome and Their Potential Interaction with Host Immune Response. Front. Immunol. 2021, 12, 705253. [Google Scholar] [CrossRef] [PubMed]
- Goorha, R. Frog Virus 3 DNA Replication Occurs in Two Stages. J. Virol. 1982, 43, 519–528. [Google Scholar] [CrossRef]
- Koonin, E.V.; Yutin, N. Evolution of the Large Nucleocytoplasmic DNA Viruses of Eukaryotes and Convergent Origins of Viral Gigantism, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 103, ISBN 9780128177228. [Google Scholar]
- Kaján, G.L.; Doszpoly, A.; Tarján, Z.L.; Vidovszky, M.Z.; Papp, T. Virus–Host Coevolution with a Focus on Animal and Human DNA Viruses. J. Mol. Evol. 2020, 88, 41–56. [Google Scholar] [CrossRef]
- Abràmoff, M.; Magalhães, P.; Ram, S. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 249–258. [Google Scholar]
- Ke, F.; Wang, Z.H.; Ming, C.Y.; Zhang, Q.Y. Ranaviruses Bind Cells from Different Species through Interaction with Heparan Sulfate. Viruses 2019, 11, 593. [Google Scholar] [CrossRef] [PubMed]
- Voller, J.; Béres, T.; Zatloukal, M.; Džubák, P.; Hajdúch, M.; Doležal, K.; Schmülling, T.; Miroslav, S. Anti-Cancer Activities of Cytokinin Ribosides. Phytochem. Rev. 2019, 18, 1101–1113. [Google Scholar] [CrossRef]
- Goh, K.C.M.; Tang, C.K.; Norton, D.C.; Gan, E.S.; Tan, H.C.; Sun, B.; Syenina, A.; Yousuf, A.; Ong, X.M.; Kamaraj, U.S.; et al. Molecular Determinants of Plaque Size as an Indicator of Dengue Virus Attenuation. Sci. Rep. 2016, 6, 26100. [Google Scholar] [CrossRef] [PubMed]
- Voller, J.; Zatloukal, M.; Lenobel, R.; Doleal, K.; Bére, T.; Krytof, V.; Spíchal, L.; Niemann, P.; Dubák, P.; Hajdúch, M.; et al. Anticancer Activity of Natural Cytokinins: A Structure-Activity Relationship Study. Phytochemistry 2010, 71, 1350–1359. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Yu, W. Frog Virus 3-Mediated Translational Shut-off: Frog Virus 3 Messages Are Translationally More Efficient than Host and Heterologous Viral Messages under Conditions of Increased Translational Stress. Virus Res. 1990, 16, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Essbauer, S.; Bremont, M.; Ahne, W. Comparison of the EIF-2α Homologous Proteins of Seven Ranaviruses (Iridoviridae). Virus Genes 2001, 23, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Rothenburg, S.; Chinchar, V.G.; Dever, T.E. Characterization of a Ranavirus Inhibitor of the Antiviral Protein Kinase PKR. BMC Microbiol. 2011, 11, 56. [Google Scholar] [CrossRef]
- Yakita, M.; Chujo, T.; Wei, F.Y.; Hirayama, M.; Kato, K.; Takahashi, N.; Naganuma, K.; Nagata, M.; Kawahara, K.; Nakayama, H.; et al. Extracellular N6-Isopentenyladenosine (I6A) Addition Induces Cotranscriptional I6A Incorporation into Ribosomal RNAs. RNA 2022, 28, 1013–1027. [Google Scholar] [CrossRef]
- Liaqat, A.; Stiller, C.; Michel, M.; Sednev, M.V.; Höbartner, C. N6-Isopentenyladenosine in RNA Determines the Cleavage Site of Endonuclease Deoxyribozymes. Angew. Chem. Int. Ed. 2020, 59, 18627–18631. [Google Scholar] [CrossRef]
- Holopainen, R.; Tapiovaara, H.; Honkanen, J. Expression Analysis of Immune Response Genes in Fish Epithelial Cells Following Ranavirus Infection. Fish Shellfish Immunol. 2012, 32, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Escalon, B.L.; Robert, J.; Chinchar, V.G. Differential Transcription of Fathead Minnow Immune-Related Genes Following Infection with Frog Virus 3, an Emerging Pathogen of Ectothermic Vertebrates. Virology 2014, 456–457, 77–86. [Google Scholar] [CrossRef]
- Chen, G.; Robert, J. Antiviral Immunity in Amphibians. Viruses 2011, 3, 2065–2086. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.J.; Li, W.P.; Mao, X.T.; Huang, T.; Wang, H.L.; Li, Y.N.; Liu, B.Q.; Zhong, J.Y.; Renjie, C.; Jin, J.; et al. Single-Nucleotide Methylation Specifically Represses Type I Interferon in Antiviral Innate Immunity. J. Exp. Med. 2021, 218, e20201798. [Google Scholar] [CrossRef] [PubMed]
- Dassano, A.; Mancuso, M.; Giardullo, P.; De Cecco, L.; Ciuffreda, P.; Santaniello, E.; Saran, A.; Dragani, T.A.; Colombo, F. N6-Isopentenyladenosine and Analogs Activate the NRF2-Mediated Antioxidant Response. Redox Biol. 2014, 2, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Blad, C.C.; von Drabbe Künzel, J.K.F.; de Vries, H.; Mulder-Krieger, T.; Bar-Yehuda, S.; Fishman, P.; IJzerman, A.P. Putative Role of the Adenosine A3 Receptor in the Antiproliferative Action of N6-(2-Isopentenyl)Adenosine. Purinergic Signal. 2011, 7, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Ciaglia, E.; Nicolin, V.; Pescatore, A.; Prota, L.; Capunzo, M.; Ursini, M.V.; Nori, S.L.; Bifulco, M. The Isoprenoid End Product N6-Isopentenyladenosine Reduces Inflammatory Response through the Inhibition of the NFκB and STAT3 Pathways in Cystic Fibrosis Cells. Inflamm. Res. 2018, 67, 315–326. [Google Scholar] [CrossRef]
- Bhutta, M.S.; Gallo, E.S.; Borenstein, R. Multifaceted Role of AMPK in Viral Infections. Cells 2021, 10, 1118. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Yang, Y.C.; Huang, C.L.; Kuo, T.Y.; Lin, J.H.; Yang, D.M.; Huang, N.K. When Cytokinin, a Plant Hormone, Meets the Adenosine A2A Receptor: A Novel Neuroprotectant and Lead for Treating Neurodegenerative Disorders? PLoS ONE 2012, 7, e38865. [Google Scholar] [CrossRef]
- Pastor-Anglada, M.; Pérez-Torras, S. Who Is Who in Adenosine Transport. Front. Pharmacol. 2018, 9, 627. [Google Scholar] [CrossRef]
- Scaletti, E.; Huschmann, F.U.; Mueller, U.; Weiss, M.S.; Sträter, N. Substrate Binding Modes of Purine and Pyrimidine Nucleotides to Human Ecto-5′-Nucleotidase (CD73) and Inhibition by Their Bisphosphonic Acid Derivatives. Purinergic Signal. 2021, 17, 693–704. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seegobin, M.; Logan, S.R.; Emery, R.J.N.; Brunetti, C.R. Cytokinins Reduce Viral Replication and Alter Plaque Morphology of Frog Virus 3 In Vitro. Viruses 2024, 16, 826. https://doi.org/10.3390/v16060826
Seegobin M, Logan SR, Emery RJN, Brunetti CR. Cytokinins Reduce Viral Replication and Alter Plaque Morphology of Frog Virus 3 In Vitro. Viruses. 2024; 16(6):826. https://doi.org/10.3390/v16060826
Chicago/Turabian StyleSeegobin, Mark, Samantha R. Logan, R. J. Neil Emery, and Craig R. Brunetti. 2024. "Cytokinins Reduce Viral Replication and Alter Plaque Morphology of Frog Virus 3 In Vitro" Viruses 16, no. 6: 826. https://doi.org/10.3390/v16060826
APA StyleSeegobin, M., Logan, S. R., Emery, R. J. N., & Brunetti, C. R. (2024). Cytokinins Reduce Viral Replication and Alter Plaque Morphology of Frog Virus 3 In Vitro. Viruses, 16(6), 826. https://doi.org/10.3390/v16060826