

Ad5-nCoV Vaccination Could Induce HLA-E Restricted CD8+ T Cell Responses Specific for Epitopes on Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Cohort
2.2. Sample Collection
2.3. Epitopes Prediction
2.4. Peptides Synthesis
2.5. K562 Cell Lines Transfected with HLA-E Molecule
2.6. K562/HLA-E Cell Binding Assay
2.7. Isolation of CD8+ T Cells with Magnetic Bead Kits
2.8. K562/HLA-E Cells Pre-Incubation with Peptides
2.9. Ex Vivo IFN-γ Enzyme-Linked Immunospot Assay (ELISpot)
2.10. Peptide/HLA-E*0103 Tetramer Staining
2.11. Statistical Analysis
3. Results
3.1. Thirty-Six HLA-E-Restricted Peptides on SARS-CoV-2 Were Predicted and Synthesized
3.2. Four Nonapeptides of SARS-CoV-2 Exhibited High Binding Affinity to Both HLA-E*0103 and HLA-E*0101 Molecules
3.3. HLA-E Restricted Epitopes of SARS-CoV-2 Were Identified by Inducing Specific IFN-γ-Producing CD8+ T Cell Responses
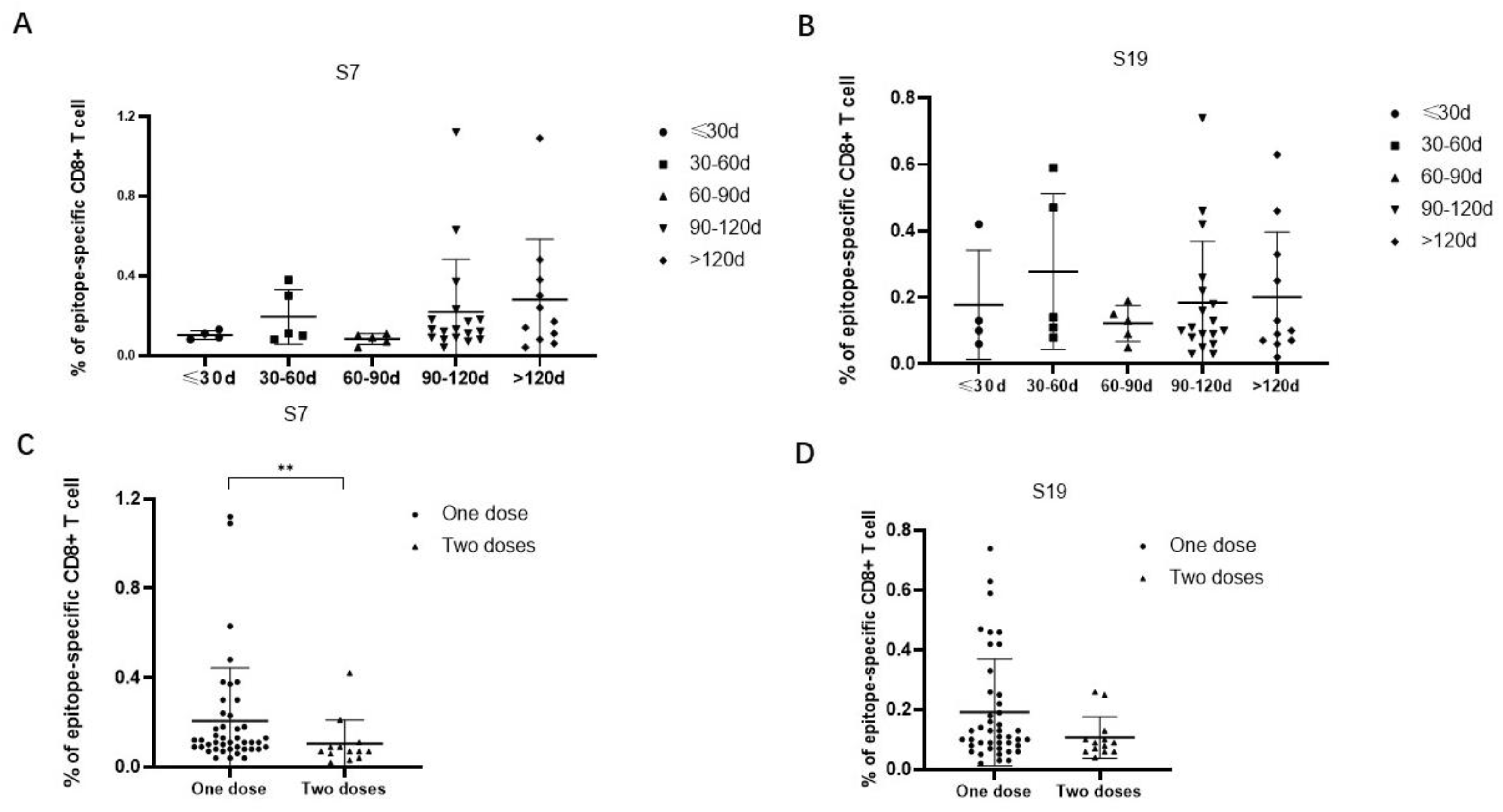
3.4. The Frequencies of HLA-E-Restricted SARS-CoV-2 Epitope-Specific CD8+ T Cells Could Be Detected in the Peripheral Blood of Vaccinated Individuals
3.5. The Frequencies of Epitope-Specific CD8+ T Cells Were Decreased in Individuals Receiving Two Versus One Dose of SARS-CoV-2 Vaccine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, J.; Lai, S.; Gao, G.F.; Shi, W. The emergence, genomic diversity and global spread of SARS-CoV-2. Nature 2021, 600, 408–418. [Google Scholar] [CrossRef]
- Wu, S.; Zhong, G.; Zhang, J.; Shuai, L.; Zhang, Z.; Wen, Z.; Wang, B.; Zhao, Z.; Song, X.; Chen, Y.; et al. A single dose of an adenovirus-vectored vaccine provides protection against SARS-CoV-2 challenge. Nat. Commun. 2020, 11, 4081. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.-C.; Li, Y.-H.; Guan, X.-H.; Hou, L.-H.; Wang, W.-J.; Li, J.-X.; Wu, S.-P.; Wang, B.-S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Guo, Y.; Zhang, G.; Sun, H.; Zhang, J.; Li, M.; Chen, Z.; Han, J.; Zhang, Y.; Zhang, X.; et al. Broadly neutralizing antibodies against Omicron-included SARS-CoV-2 variants induced by vaccination. Signal Transduct. Target. Ther. 2022, 7, 139. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.-C.; Guan, X.-H.; Li, Y.-H.; Huang, J.-Y.; Jiang, T.; Hou, L.-H.; Li, J.-X.; Yang, B.-F.; Wang, L.; Wang, W.-J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, L.; Guo, X.; Jin, P.; Wu, S.; Zhu, J.; Pan, H.; Wang, X.; Song, Z.; Wan, J.; et al. Heterologous AD5-nCOV plus CoronaVac versus homologous CoronaVac vaccination: A randomized phase 4 trial. Nat. Med. 2022, 28, 401–409. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Zhang, Z.; Wu, J.; Zhang, J.; Hu, H.; Zhu, T.; Zhang, J.; Luo, L.; Fan, P.; et al. Safety, tolerability, and immunogenicity of an aerosolised adenovirus type-5 vector-based COVID-19 vaccine (Ad5-nCoV) in adults: Preliminary report of an open-label and randomised phase 1 clinical trial. Lancet Infect. Dis. 2021, 21, 1654–1664. [Google Scholar] [CrossRef]
- Terreri, S.; Mortari, E.P.; Vinci, M.R.; Russo, C.; Alteri, C.; Albano, C.; Colavita, F.; Gramigna, G.; Agrati, C.; Linardos, G.; et al. Persistent B cell memory after SARS-CoV-2 vaccination is functional during breakthrough infections. Cell Host Microbe 2022, 30, 400–408.e4. [Google Scholar] [CrossRef]
- Edridge, A.W.D.; Kaczorowska, J.; Hoste, A.C.R.; Bakker, M.; Klein, M.; Loens, K.; Jebbink, M.F.; Matser, A.; Kinsella, C.M.; Rueda, P.; et al. Seasonal coronavirus protective immunity is short-lasting. Nat. Med. 2020, 26, 1691–1693. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Schulien, I.; Kemming, J.; Oberhardt, V.; Wild, K.; Seidel, L.M.; Killmer, S.; Sagar, N.; Daul, F.; Salvat Lago, M.; Decker, A.; et al. Characterization of pre-existing and induced SARS-CoV-2-specific CD8(+) T cells. Nat. Med. 2021, 27, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Thevarajan, I.; Nguyen, T.H.O.; Koutsakos, M.; Druce, J.; Caly, L.; van de Sandt, C.E.; Jia, X.; Nicholson, S.; Catton, M.; Cowie, B.; et al. Breadth of concomitant immune responses prior to patient recovery: A case report of non-severe COVID-19. Nat. Med. 2020, 26, 453–455. [Google Scholar] [CrossRef] [PubMed]
- McMahan, K.; Yu, J.; Mercado, N.B.; Loos, C.; Tostanoski, L.H.; Chandrashekar, A.; Liu, J.; Peter, L.; Atyeo, C.; Zhu, A.; et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021, 590, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Habel, J.R.; Nguyen, T.H.; van de Sandt, C.E.; Juno, J.A.; Chaurasia, P.; Wragg, K.; Koutsakos, M.; Hensen, L.; Jia, X.; Chua, B.; et al. Suboptimal SARS-CoV-2-specific CD8(+) T cell response associated with the prominent HLA-A*02:01 phenotype. Proc. Natl. Acad. Sci. USA 2020, 117, 24384–24391. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Strålin, K.; Gorin, J.-B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Felce, S.L.; Dong, D.; Penkava, F.; Mentzer, A.J.; Yao, X.; Liu, G.; Yin, Z.; Chen, J.-L.; Lu, Y.; et al. An immunodominant NP105-113-B*07:02 cytotoxic T cell response controls viral replication and is associated with less severe COVID-19 disease. Nat. Immunol. 2022, 23, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Joosten, S.A.; Sullivan, L.C.; Ottenhoff, T.H. Characteristics of HLA-E Restricted T-Cell Responses and Their Role in Infectious Diseases. J. Immunol. Res. 2016, 2016, 2695396. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Gehre, M.N.; Qin, K.; Sterrett, S.; Ali, A.; Dang, Y.; Abraham, S.; Costanzo, M.C.; Venegas, L.A.; Tang, J.; et al. HLA-E-restricted HIV-1-specific CD8+ T cell responses in natural infection. J. Clin. Investig. 2021, 131, e148979. [Google Scholar] [CrossRef]
- Sharpe, H.R.; Bowyer, G.; Brackenridge, S.; Lambe, T. HLA-E: Exploiting pathogen-host interactions for vaccine development. Clin. Exp. Immunol. 2019, 196, 167–177. [Google Scholar] [CrossRef]
- Kraemer, T.; Blasczyk, R.; Bade-Doeding, C. HLA-E: A novel player for histocompatibility. J. Immunol. Res. 2014, 2014, 352160. [Google Scholar] [CrossRef]
- Caccamo, N.; Sullivan, L.C.; Brooks, A.G.; Dieli, F. Harnessing HLA-E-restricted CD8 T lymphocytes for adoptive cell therapy of patients with severe COVID-19. Br. J. Haematol. 2020, 190, e185–e187. [Google Scholar] [CrossRef] [PubMed]
- Yuling, W.; Boquan, J.; Na, L.; Ying, T.; Yun, D.; Qi, L.; Ying, M.; Ran, Z. Preparation and identification of monoclonal antibodies against structural proteins of SARS-CoV-2. J. Air Force Med. Univ. 2023, 44, 595–601. [Google Scholar]
- Sahin, U.; Muik, A.; Vogler, I.; Derhovanessian, E.; Kranz, L.M.; Vormehr, M.; Quandt, J.; Bidmon, N.; Ulges, A.; Baum, A.; et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature 2021, 595, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.J.; Pade, C.; Gibbons, J.M.; Otter, A.D.; Lin, K.M.; Muñoz Sandoval, D.; Pieper, F.P.; Butler, D.K.; Liu, S.; Joy, G.; et al. Immune boosting by B.1.1.529 (Omicron) depends on previous SARS-CoV-2 exposure. Science 2022, 377, eabq1841. [Google Scholar] [CrossRef]
- Rodda, L.B.; Morawski, P.A.; Pruner, K.B.; Fahning, M.L.; Howard, C.A.; Franko, N.; Logue, J.; Eggenberger, J.; Stokes, C.; Golez, I.; et al. Imprinted SARS-CoV-2-specific memory lymphocytes define hybrid immunity. Cell 2022, 185, 1588–1601.e14. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Dolan, C.; Barouch, D.H. COVID-19 Vaccines: Adenoviral Vectors. Annu. Rev. Med. 2022, 73, 41–54. [Google Scholar] [CrossRef]
- Sakurai, F.; Tachibana, M.; Mizuguchi, H. Adenovirus vector-based vaccine for infectious diseases. Drug Metab. Pharmacokinet. 2022, 42, 100432. [Google Scholar] [CrossRef]
- Bortolotti, D.; Gentili, V.; Rizzo, S.; Rotola, A.; Rizzo, R. SARS-CoV-2 Spike 1 Protein Controls Natural Killer Cell Activation via the HLA-E/NKG2A Pathway. Cells 2020, 9, 1975. [Google Scholar] [CrossRef]
- Hammer, Q.; Dunst, J.; Christ, W.; Picarazzi, F.; Wendorff, M.; Momayyezi, P.; Huhn, O.; Netskar, H.K.; Maleki, K.T.; García, M.; et al. SARS-CoV-2 Nsp13 encodes for an HLA-E-stabilizing peptide that abrogates inhibition of NKG2A-expressing NK cells. Cell Rep. 2022, 38, 110503. [Google Scholar] [CrossRef]
- Grant, E.J.; Nguyen, A.T.; Lobos, C.A.; Szeto, C.; Chatzileontiadou, D.S.; Gras, S. The unconventional role of HLA-E: The road less traveled. Mol. Immunol. 2020, 120, 101–112. [Google Scholar] [CrossRef]
- Tennøe, S.; Gheorghe, M.; Stratford, R.; Clancy, T. The T Cell Epitope Landscape of SARS-CoV-2 Variants of Concern. Vaccines 2022, 10, 1123. [Google Scholar] [CrossRef] [PubMed]
- Weingarten-Gabbay, S.; Klaeger, S.; Sarkizova, S.; Pearlman, L.R.; Chen, D.Y.; Gallagher, K.M.; Bauer, M.R.; Taylor, H.B.; Dunn, W.A.; Tarr, C.; et al. Profiling SARS-CoV-2 HLA-I peptidome reveals T cell epitopes from out-of-frame ORFs. Cell 2021, 184, 3962–3980.e17. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples ID | Gender | Age (y) | Variety of Vaccine Received | Manufacturer | Inoculation Times | Days Since Latest Inoculation |
|---|---|---|---|---|---|---|
| No.1 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 58 |
| No.2 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 127 |
| No.3 | M | 23 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 54 |
| No.4 | M | 31 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 124 |
| No.5 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 59 |
| No.6 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 120 |
| No.7 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 128 |
| No.8 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 28 |
| No.9 | M | 21 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 56 |
| No.10 | M | 22 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 56 |
| No.11 | M | 32 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 27 |
| No.12 | M | 22 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 90 |
| No.13 | F | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 164 |
| No.14 | M | 23 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 95 |
| No.15 | F | 30 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 159 |
| No.16 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 95 |
| No.17 | M | 30 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 164 |
| No.18 | M | 25 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 96 |
| No.19 | M | 22 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 91 |
| No.20 | M | 32 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 166 |
| No.21 | F | 30 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 93 |
| No.22 | M | 21 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 92 |
| No.23 | M | 21 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 92 |
| No.24 | M | 22 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 100 |
| No.25 | M | 22 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 100 |
| No.26 | M | 23 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 100 |
| No.27 | F | 23 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 165 |
| No.28 | M | 21 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 101 |
| No.29 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 101 |
| No.30 | M | 23 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 101 |
| No.31 | M | 21 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 101 |
| No.32 | M | 24 | inactivated vaccine | Sinovac Biotech | 1 | 100 |
| No.33 | F | 25 | inactivated vaccine | Sinovac Biotech | 1 | 87 |
| No.34 | F | 24 | inactivated vaccine | Sinovac Biotech | 1 | 128 |
| No.35 | M | 29 | adenovirus-vectored vaccine | CanSino Biologics | 1 | 171 |
| No.36 | M | 26 | inactivated vaccine | Sinovac Biotech | 1 | 92 |
| No.37 | F | 23 | inactivated vaccine | Sinovac Biotech | 1 | 104 |
| No.38 | M | 25 | inactivated vaccine | Sinovac Biotech | 1 | 89 |
| No.39 | M | 39 | inactivated vaccine | China National Biotec Group Company | 1 | 220 |
| No.40 | M | 23 | recombinant protein subunit vaccine | Zhifei Biological | 1 | 27 |
| No.41 | M | 24 | recombinant protein subunit vaccine | Zhifei Biological | 1 | 27 |
| No.42 | F | 24 | recombinant protein subunit vaccine | Zhifei Biological | 1 | 64 |
| No.43 | F | 26 | recombinant protein subunit vaccine | Zhifei Biological | 1 | 68 |
| No.44 | F | 25 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 3 |
| No.45 | M | 32 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 3 |
| No.46 | F | 23 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 2 |
| No.47 | F | 24 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 4 |
| No.48 | M | 23 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 4 |
| No.49 | M | 23 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 4 |
| No.50 | M | 25 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 5 |
| No.51 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 5 |
| No.52 | M | 23 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 5 |
| No.53 | M | 24 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 5 |
| No.54 | M | 22 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 6 |
| No.55 | M | 22 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 5 |
| No.56 | F | 31 | adenovirus-vectored vaccine | CanSino Biologics | 2 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Yang, L.; Tang, K.; Zhang, Y.; Zhang, C.; Zhang, Y.; Jin, B.; Zhang, Y.; Zhuang, R.; Ma, Y. Ad5-nCoV Vaccination Could Induce HLA-E Restricted CD8+ T Cell Responses Specific for Epitopes on Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein. Viruses 2024, 16, 52. https://doi.org/10.3390/v16010052
Wang Y, Yang L, Tang K, Zhang Y, Zhang C, Zhang Y, Jin B, Zhang Y, Zhuang R, Ma Y. Ad5-nCoV Vaccination Could Induce HLA-E Restricted CD8+ T Cell Responses Specific for Epitopes on Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein. Viruses. 2024; 16(1):52. https://doi.org/10.3390/v16010052
Chicago/Turabian StyleWang, Yuling, Lu Yang, Kang Tang, Yusi Zhang, Chunmei Zhang, Yun Zhang, Boquan Jin, Yuan Zhang, Ran Zhuang, and Ying Ma. 2024. "Ad5-nCoV Vaccination Could Induce HLA-E Restricted CD8+ T Cell Responses Specific for Epitopes on Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein" Viruses 16, no. 1: 52. https://doi.org/10.3390/v16010052
APA StyleWang, Y., Yang, L., Tang, K., Zhang, Y., Zhang, C., Zhang, Y., Jin, B., Zhang, Y., Zhuang, R., & Ma, Y. (2024). Ad5-nCoV Vaccination Could Induce HLA-E Restricted CD8+ T Cell Responses Specific for Epitopes on Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein. Viruses, 16(1), 52. https://doi.org/10.3390/v16010052