Abstract
The metagenomic analysis of mosquitoes allows for the genetic characterization of mosquito-associated viruses in different regions of the world. This study applied a metagenomic approach to identify novel viral sequences in seven species of mosquitoes collected from the Novosibirsk region of western Siberia. Using NGS sequencing, we identified 15 coding-complete viral polyproteins (genomes) and 15 viral-like partial sequences in mosquitoes. The complete sequences for novel viruses or the partial sequences of capsid proteins, hypothetical viral proteins, and RdRps were used to identify their taxonomy. The novel viral sequences were classified within the orders Tymovirales and Picornavirales and the families Partitiviridae, Totiviridae, Tombusviridae, Iflaviridae, Nodaviridae, Permutotetraviridae, and Solemoviridae, with several attributed to four unclassified RNA viruses. Interestingly, the novel putative viruses and viral sequences were mainly associated with the mosquito Coquillettidia richardii. This study aimed to increase our understanding of the viral diversity in mosquitoes found in the natural habitats of Siberia, which is characterized by very long, snowy, and cold winters.
1. Introduction
Mosquitoes are well known to transmit numerous arboviruses that cause viral infections in animals and humans, such as the West Nile virus (WNV) and the Zika, Japanese encephalitis, Chikungunya, and dengue viruses [1,2,3,4]. In recent years, the development of metagenomic approaches has led to the discovery of many novel viruses in invertebrates [5,6,7,8,9,10]. Studying the viromes of different species of mosquitoes has revealed new viruses referred to as insect-specific viruses (ISVs). The viral interference within the ISV group and pathogenic viruses may dramatically change the viral biodiversity in mosquitoes and, thereby, predetermine the transmission of pathogenic viruses from mosquitoes to animals and humans [11,12,13,14,15,16]. These are viruses belonging to the families Peribunyaviridae, Flaviviridae, Reoviridae, and Togaviridae, and each is a potential source of viral biodiversity for viruses, with dual tropism for invertebrate and vertebrate hosts [7,17,18].
Comparing the viromes of mosquitoes from different geographical regions has revealed their biodiversity, providing new insights into the phylogeography of mosquito-borne viruses [5,6,19,20]. Generally, such studies are conducted in countries characterized by warm or tropical climates, such as China, Australia, Mozambique, and the USA, where mosquito-borne viral infections are not uncommon. The Sindbis, Inco, and West Nile viruses are usually detected and isolated from mosquitoes in the southern regions of Russia [21,22]. No systemic information is available for western Siberia, where it is also possible for mosquito-borne viruses to circulate. This region has a continental climate with long winters and short summers that may limit the biodiversity of mosquito species and mosquito-associated viruses. Only a few studies have reported the detection of WNV markers in bird and human cases of West Nile fever in western Siberia [22,23].
In this study, we sought to investigate the biodiversity of mosquito-borne viruses from different mosquito species collected in western Siberia using metagenomic approaches.
2. Materials and Methods
2.1. Mosquito Samples
For this study, 3910 mosquitoes were collected in the Novosibirsk region during the spring–summer periods of 2017–2018. The collection sites were situated in typical mosquito habitats in western Siberia (Figure 1): deciduous and mixed forests with well-developed grassy cover, deforestations with the natural resumption of hardwoods, and the banks of streams. The mosquitoes were collected using a light trap (https://survinat.ru/2011/09/metodika-sborov-xraneniya-i-izucheniya-komarov/, accessed on 23 June 2023). Capture was conducted after sunset. The mosquitoes were transported in a thermal bag on a damp napkin at a temperature of 4 °C and stored at minus 18–24 °C. Fragments of the 16S rRNA and the COI gene of the mitogenome were sequenced via the Sanger method to determine the mosquito species [11]. Pools of 10–40 mosquitoes were formed according to the morphological features of mosquito species, and fragments of the 16S rRNA and the COI gene from individual mosquitoes were selectively sequenced to determine their species.
Figure 1.
The location from which mosquitoes were collected for this study, with the upper picture showing the world and the lower picture showing the Novosibirsk region.
2.2. Sample Preparation
All the mosquitoes were washed with 70% ethanol and then rinsed twice with water, followed by homogenization to remove potential surface microorganisms. The homogenization of the samples was performed mechanically by grinding them in a mortar with 300 µL of sterile saline. The homogenates were centrifuged at 8000× g for 5 min at 4 °C, and the supernatants were used for the analysis. The total RNA was extracted using an Extract RNA reagent (Eurogen, Moscow, Russia) according to the manufacturer’s protocol and purified on Cleanup Mini spin columns (Eurogen, Moscow, Russia). The pools were then processed using benzonaze [24]. The first cDNA chain was synthesized using the NEBNext Ultra Direction module. The second cDNA chain was synthesized using the UMI Second Strand Synthesis module (Illumina, San Diego, CA, USA, Lexogen, Wien, Austria).
2.3. NGS Sequencing and Phylogenetic Analysis
The dsDNA libraries were prepared and analyzed via NGS on MiSeq using Illumina technology. Cutadapt (version 1.18) and SAMtools (version 0.1.18) were used to remove the Illumina adapters and re-read. The contigs were assembled de novo using MIRA assembly (version 4.9.6). The experimentally determined sequences were deposited in GenBank. The phylogenetic analysis was performed using RNA-dependent RNA polymerase (RdRp) sequences from GenBank with an amino acid identity > 20%. The sequences were aligned, and phylogenetic trees were built in Vector NTI Advance 11, MEGA 7/10 (PSU, USA), and Lasergen 7 (Invitrogen, Waltham, MA, USA). The resulting viral sequences and sequence read archive (SRA) were deposited in GenBank. The phylogenetic trees were calculated via the maximum likelihood method, using 500 replicates for bootstrap values.
3. Results
3.1. Mosquito Species
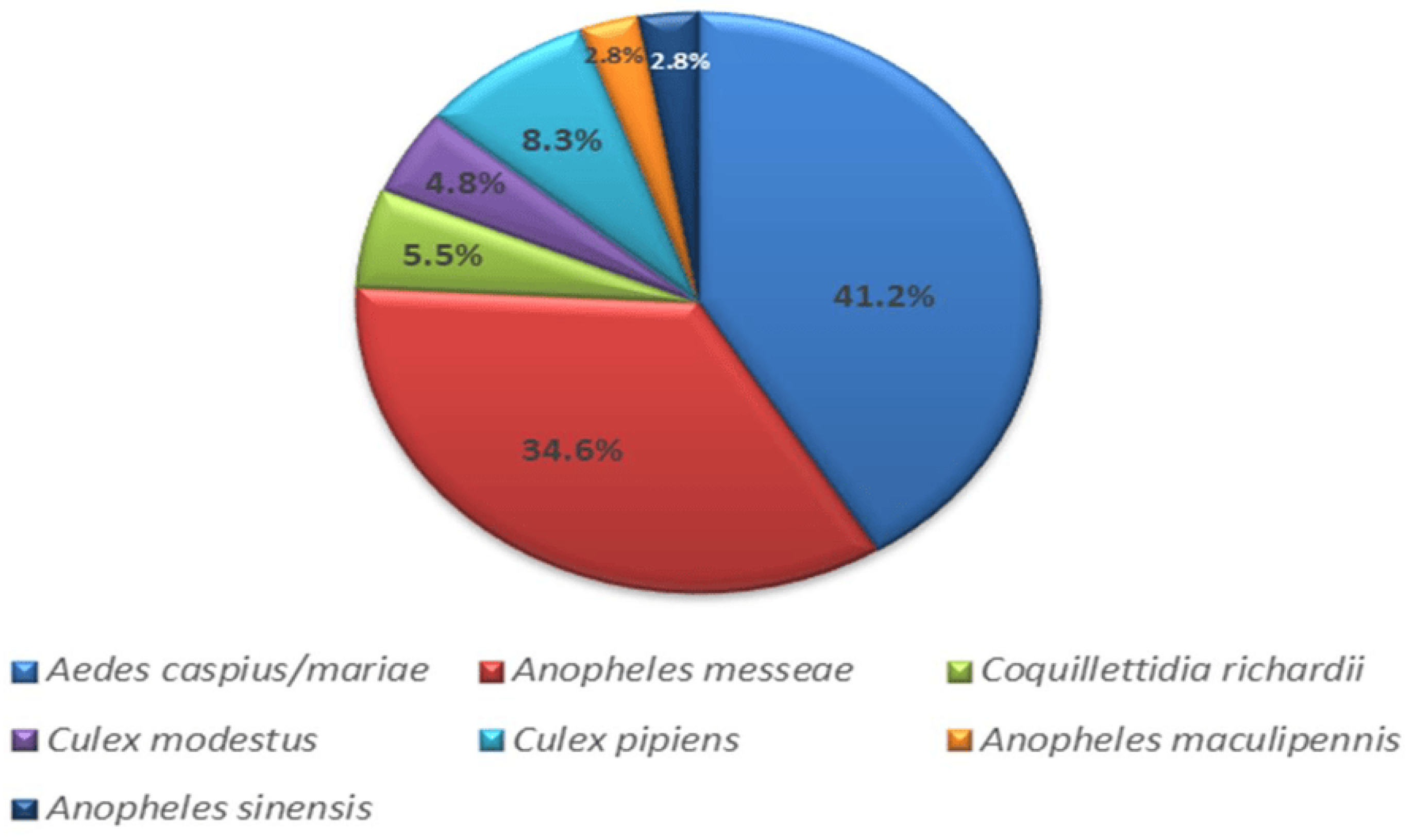
A total of 3910 mosquitoes were collected in the Novosibirsk suburbs and the rural district of the Novosibirsk region in 2017–2018. Pools of 10–40 mosquitoes were formed to identify the mosquito species for each collection point. Fragments of the 16S rRNA and the COI gene of the mitogenome were sequenced to determine the species of mosquitoes (Figure 2). Aedes caspius (Pallas, 1771) and Ae. mariae (Sergent and Sergent, 1903) were found to be the most abundant species, comprising 41.2% of the total. Anopheles messeae (Falleroni, 1926) accounted for 34.6%, Culex pipiens (Linnaeus, 1758) for 8.3%, Coquillettidia richardii (Ficalbi, 1889) for 5.5%, C. modestus (Ficalbi, 1889) for 4.8%, and An. maculipennis (Meigen, 1818) and An. sinensis (Wiedemann, 1828) for 2.8% each.
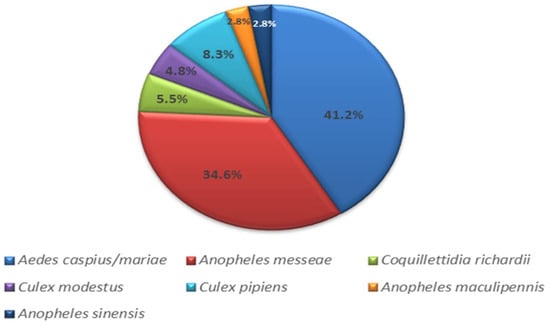
Figure 2.
Histogram of mosquito species collected in the southern part of western Siberia (Novosibirsk region).
3.2. NGS Sequencing
In total, 144 putative viral sequences were selected in the first step, and 30 of these sequences with lengths greater than 1239 bp were chosen. Eight sequences with a level of identity > 80% aa were previously described as mosquito-borne viruses (Table 1). These were Partitivirus-like 1 (dsRNA, Partitiviridae), Hammarskog tombus-like virus (ssRNA (+), Tombusviridae), Hammarskog picorna-like virus (ssRNA (+), Picornavirales, unclassified), Lymantria dispar iflavirus 1 (ssRNA (+) virus, Iflavirus), Wenzhou noda-like virus 6 (ssRNA (+) virus, unclassified), Mayapan virus (ssRNA (+), two segments), Sanxia permutotetra-like virus 1 (ssRNA (+) virus, unclassified), and Chaq virus-like 1 (RNA virus, unclassified). An additional 22 viral sequences were presented as putative mosquito-borne sequences, with a level of identity less than 79% for viral prototype sequences.

Table 1.
List of putative and novel mosquito-associated viruses detected in western Siberia.
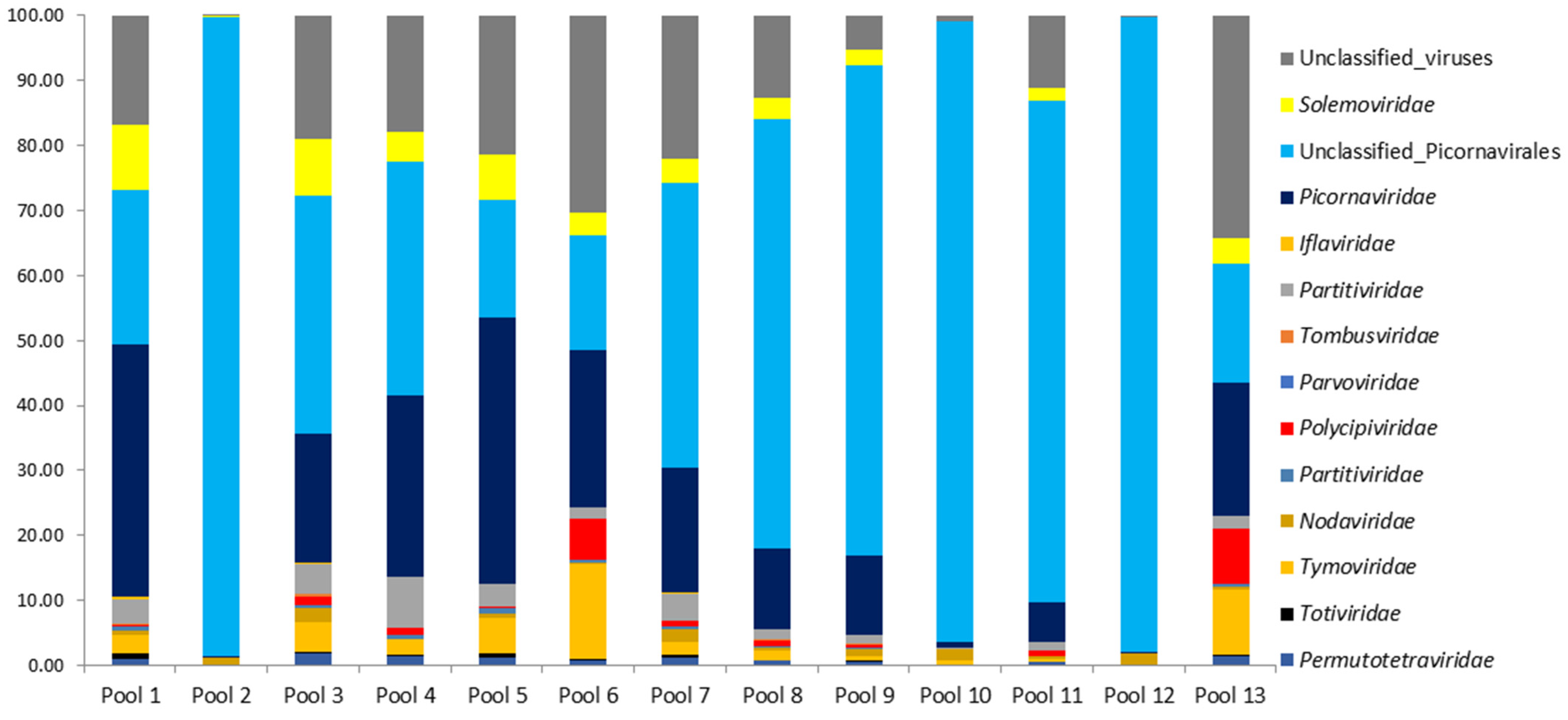
The proportions for classified and unclassified viral reads are presented for 13 mosquito pools of Cq. richardii (Figure 3). The prevalence of unclassified and classified picornaviruses was detected in practically all the pools studied. Unclassified viral sequences were also analyzed, ranging from 0.13% to 34.27% for the different pools.
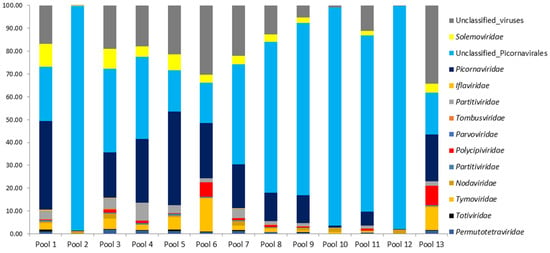
Figure 3.
Annotation of the taxonomy for the viral reads in the different pools of the Cq. richardii mosquito. Proportions for classified and unclassified sequences are in %.
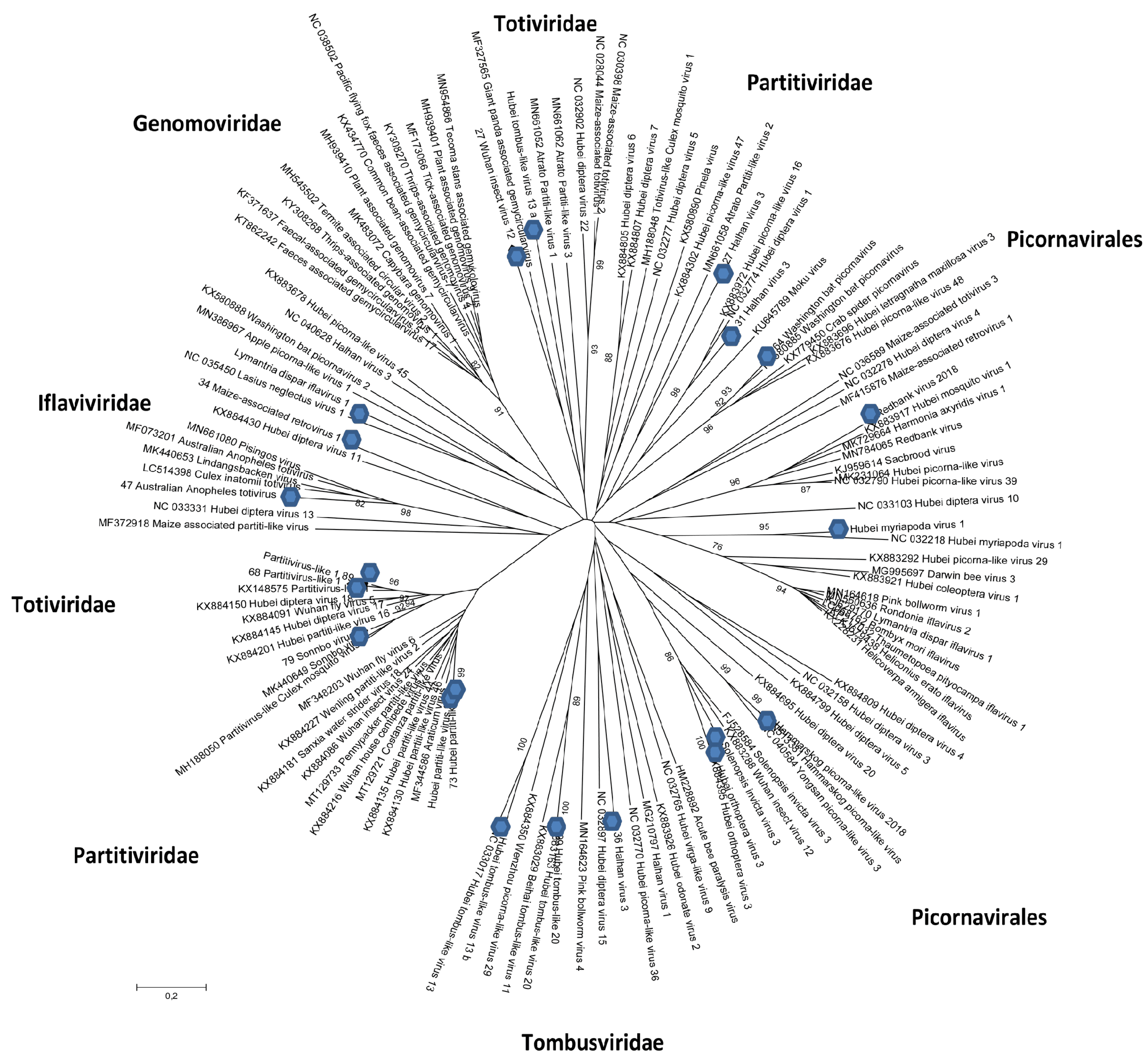
The phylogenetic analysis results for the sequences obtained from the mosquitoes are presented in Figure 4. This phylogeny was based on the amino acid sequences of RdRps, with these data confirming the biodiversity of the mosquito viruses found in nature.
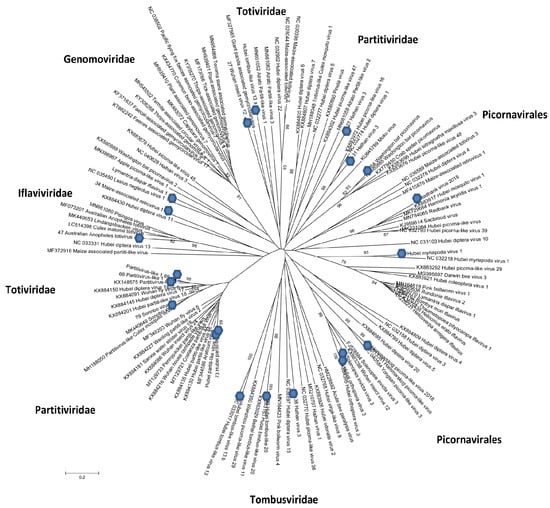
Figure 4.
Schematic phylogenetic tree for RNA viruses belonging to different families found in mosquitoes collected in western Siberia in 2017–2018. Maximum likelihood phylogenetic tree based on the amino acid sequences of RdRps. Branch numbers indicate bootstrap support in percentage out of 1000 replicates. The sequences from this study are marked with circles. Tymovirales (MW251314; MW251315); Partitiviridae (MW251327; MW389552; MW389553; MW389554; MW389555; MW251328; MW251329; MW251330; MW251325; MW389559; MW389560; MW389561); Totiviridae (MW251336); Tombusviridae (MW251332; MW251324); Picornavirales (MT753151; MW251320; MW251321; MW251322; MW251316; MW251317; MW251323; MW251313); Iflaviridae (MW251319; MT753152; MT753153; MW389556; MW389557); Permutotetraviridae (MT753154; MW389558).
3.2.1. Tymovirales (Positive ssRNA)
The complete viral genome of a tymovirus-like sequence with an identity level of 60% (according to its amino acid sequence) with a previously described insect-associated tymovirus 1 in Mexico (MN203215) was detected in the Cq. richardii mosquitoes. This virus has been designated as an Inya insect-associated virus 1 (MW251313; MW251314). The genomic positive ssRNA of the Inya insect-associated virus was identified as comprising 6526 bp and three ORF-encoding proteins (Figure S1). The ORF MP of Inya insect-associated virus 1 was an RdRp, and this ORF contained a highly conservative “tymobox” near the 5′-end [25]. The tymobox sequence had 16 nucleotides that were likely part of the subgenomic promoter for the third ORF encoding the coat protein (CP). Previously, the Tymovirales were well-known as plant viruses [26]. Inya insect-associated virus 1 can presumably be identified taxonomically as belonging to the order Tymovirales, unclassified Tymovirales.
3.2.2. Partitiviridae (dsRNA)
Partitivirus-like 1 was detected in Cq. richardii and demonstrated an identity level of 89% with an isolate from Anopheles gambiae that was collected in Liberia (KX148575). Another novel putative partitivirus was detected in the pool of Cq. richardii mosquitoes (Figure S2) and was designated as Krahall insect-associated virus 1 and 2, with an identity level of 60–63% shared with a previously described Atrato partiti-like virus 2, which was isolated earlier from Anopheles darlingi in Colombia (MN661058). In addition, seven suspected partitiviruses were found in the pool of Cq. richardii mosquitoes. These are the novel insect Talaya 1 and 2 viruses (MW251327–MW251330), with aa sequence identity levels of 68–71% identity with a previously described Partitivirus-like 1 (Liberia, KX148575); the Tarbrook virus (MW251325), with 79% homology with a previously described Sonnbo virus in Sweden (MK440649); and Zeyabrook partiti-like_viruses 1 and 2, with similarities of 38–42% with a Beihai partiti-like virus 2 (NC_032500) from China. All the prototype sequences were isolated earlier from invertebrates (mollusks, octopuses, mosquitoes, and odonatos). These partitiviruses were preliminarily taxonomically identified as belonging to the family Partitiviridae, unclassified Partitiviridae.
3.2.3. Totiviridae (Monopartite dsRNA)
We have found a novel putative totivirus, designated Zyryana toti-like virus 2, with the prototype Fitzroy Crossing toti-like virus 1 isolated earlier from Culex annulirostris in Australia (MT498830) (Table 1). The Zyryana toti-like virus 2, with an identity level of 51% with the prototype sequence, was detected in the pools of An. Messeae and Cq. richardii mosquitoes. The length of the nucleotide sequence of the Zyryana toti-like virus 2 was over 5863 bp, and the ORFs encoded two proteins, 1112 aa and 807 aa, of the CP and RdRp. The putative genome organization schemes for these totiviruses are presented in Figure S3.
3.2.4. Tombusviridae (Positive ssRNA)
Tombusviridae (Tolivirales, Tombusviridae) are single-stranded RNA (+) genomes between 3.7 and 4.8 kb in length, currently regarded as plant viruses with a relatively limited selection of hosts, and they usually require a subviral RNA for replication [27,28,29]. These viruses are frequently found in insects, and the results show that tombusviruses were first detected in mosquitoes in western Siberia. We present the complete polyprotein (4317 bp) for the Hammarskog tombus-like virus, which had 90% identity with a similar virus that was detected in Sweden (MN513379) and isolated from Cq. Richiardii in 2017 [30]. The 4166 bp partial polyprotein contained three ORF-encoded hypothetical polypeptides, 397 aa, 482 aa, and 409 aa, which differed for Hubei tombus-like 20 (Figure S4). In addition, we found a novel Oyosh tombus-like virus with 62% identity with Hubei tombus-like virus 13 (NC033017), which was isolated from house centipedes in China. Four polypeptides were encoded by a prototype genome (5904 bp). The RdRp for Tombusviridae was translated using a potential alternative mechanism to suppress the stop-codon-reading mechanism with the formation of a full-size protein with an elongated ORF1 C-end [6].
3.2.5. Picornavirales (Positive ssRNA)
Most members of the order Picornavirales have a single molecule of positive-sense RNA ranging in length between 7000 and 12,500 nt. The viral RNA is infectious and serves as a template for replication and mRNA [31]. Six different picorna-like viruses were identified as mosquito-associated viruses in western Siberia (Table 1). We assembled a complete genome for the Hammarskog picorna-like virus (11,507 bp) from the Cq. richardii mosquitoes that had five OFRs encoding 175 aa, 156 aa, 121aa, 376 aa, and 2424 aa polypeptides with 98% aa identity with a previously described Hammarskog picorna-like virus (MN513381) isolated from Cq. richiardii in Sweden (Figure S5). In addition, other novel picorno-like viruses were found in the Cq. richardii mosquitoes collected in the Novosibirsk region. These are Miltyush picorna-like viruses 1 and 2, the Isses picorna-like virus, the Ichacreek insect virus, and the polycipiviridae associated with Ora rivulet insects.
The Miltyush picorna-like virus was found to have only 36% identity with a previously detected Halhan virus 3 from Haliotis discus hannai in Korea (NC040628). Isses picorna-like virus 1 was found to have 64% identity with a Washington bat picornavirus previously discovered in the USA (KX580885). Ichacreek insect virus 3 was identified to have a 44% level of identity with a previously detected Solenopsis invicta virus 3 (GU017972) from Solenopsis invicta in Argentina. Ora rivulet insect-associated polycipiviridae were identified to have a 34% identity level with previously discovered Polycipivirida sp. isolated from Pteropus lylei in Cambodia (MK161350). The preliminary taxonomic identification for these six viruses is Picornavirales, unclassified Picornavirales.
3.2.6. Iflaviridae (Positive ssRNA)
The order Picornavirales also includes some iflaviruses that were found in Cq. richardii mosquitoes in this study. Lymantria dispar iflavirus 1 was detected, with its sequence having 99% identity with already-known viral isolates from the USA and Russia (KJ629170, MN938851). The alignment and phylogenetic analysis revealed a high level of sequence identity with the representatives of Iflavirus, the family Iflaviridae.
3.2.7. Nodaviridae (Bi-Partite Positive-Sense, ssRNA, Sometimes Associated with Subviral RNA Molecule Capable of Replicating Itself)
We assembled practically the whole genome for Wenzhou noda-like virus 6 from the Cq. richardii mosquitoes, as well as the Mayapan virus, with identity levels of 87% and 90%, respectively. Previously, the Wenzhou noda-like virus 6 sequence was identified in Channeled applesnail in China (KX883260), and the Mayapan virus (MH719096) was isolated from the Psorophora ferox mosquito in Mexico (Figure S6). Other novel nodaviruses were also found in the Cq. richardii mosquitoes collected in the Novosibirsk region. These were Mayzas noda-like virus RNA 2, with a prototype Mayapan virus RNA2 segment (MH719097), which had a 43% identity with that isolated from the Psorophora ferox mosquito in Mexico, and the insect-associated Uzakla virus with a prototype Mosinovirus (KJ632942), which had a 36% identity with that isolated from Culicidae spp. in the Cote d’Ivoire.
3.2.8. Permutotetraviridae (dsRNA)
The genomic RNA for permutotetraviruses is 4582 bp long and encodes three ORFs overlapping in a short region (Figure S7). The longest ORF (1028 aa) encoding the RdRp overlaps for 106 nucleotides with a small ORF (199 aa), which presumably encodes the capsid protein. Like all permutotetraviruses, the sequences from the Cq. richardii mosquito pools showed the presence of the virus with a 93% identity with Sanxia permutotetra-like virus 1, which was previously detected in water striders in China (KX883450). The Uzakla mosquito-associated permutotetra-like virus, with a 53% identity with Vespa velutina permutotetra-like virus 2 in France (MN5650551; MN565052), was described early on as belonging to the group of unclassified Permutotetraviridae.
3.2.9. Other Viruses and Viral Sequences
Two variants of Chaq virus-like 1, Tartas insect associate virus, ZeyaBrook chaq-like virus 2, Kamenka insect-associated virus, and the Uzakla insect virus were detected in Cq. richardii. The Chaq virus-like 1 had 82% identity with an earlier described unclassified sequence from Anopheles gambiae in Liberia (KX148554). The ZeyaBrook chaq-like virus 2, Kamenka insect-associated virus, and Uzakla insect virus had identity levels of 33–47% with previously unclassified putative viral sequences (KX148556, KX883594, and NC032218, respectively) isolated from invertebrates. Only the Tartas insect associate virus may be classified as unclassified Solemoviridae, having an identity level of 61% with prototype Atrato Sobemo-like virus 6 (MN661101), which was detected in Wyeomyia spp. mosquitoes in Colombia. The Solemoviridae have a relatively small (4–4.6 kb) positive-sense, single-stranded, monopartite RNA genome with 4–5 ORFs, and they are usually associated with plant viruses.
4. Discussion
The application of the metagenomic approach offers novel opportunities for virome analysis [5,7,20]. This approach has provided new insights into the evolution of viruses of clinical importance and has allowed new viruses to be discovered from different viral families, such as Peribunyaviridae [5], Rhabdoviridae [32], Orthomyxoviridae [7,33], Flaviviridae [34], and Reoviridae [35], as well as the unclassified Chuvirus [7] and Negevirus [36]. Recent metagenomic studies have also confirmed the presence of the dengue virus, Zika virus, and Japanese encephalitis virus in mosquitoes in China [37,38].
Numerous genetically diverse viruses have also been detected via NGS sequencing in plants, invertebrates, and vertebrates in tropical countries [7,8]. Phylogenetic analyses have demonstrated that it is possible for all host species and viruses to co-evolve by changing hosts. Mosquitoes are among the most common and important viral vectors of the Zika, dengue, yellow fever, and West Nile viruses and are associated with unprecedented global outbreaks of these infectious diseases in tropical countries [39,40]. In addition, mosquitoes are also known to carry insect-specific viruses. Although they do not directly affect humans and animals, these viruses can modulate the transmission of pathogenic viruses to vertebrates [41,42]. The growth of tourism and trade has also led to intensive exchanges of viral pathogens and their vectors in different geographic regions. Together with the rapid growth of large cities in tropical countries, these are the basis for outbreaks or/and epidemics of mosquito-borne infections among animals and humans, creating environments that can maintain the transmission of zoonotic infections [43]. In addition, viruses have extraordinary evolutionary potential for generating new pathogenic isolates that can cause severe diseases in humans and/or animals.
The south of the Western Siberian Plain is characterized by a continental climate, with short, warm summers and long winters; uniform humidity; and rather abrupt changes in all weather components over relatively short periods of time [44]. This region has experienced characteristic negative mean annual temperatures during the last century, with the maximum variation in the mean annual temperature being 3.6 °C over the observation period. The activity season for different species of mosquitoes begins when the ambient temperature rises above 0 °C (early May) and ends in late August or early September, depending on the year. The maximum duration of the period of mosquito activity is approximately four or five months. Seventeen species of mosquitoes were previously found in the forest–steppe and steppe zones of the region [45]. The composition of mosquito species from different foci can drastically vary. For example, the concentrations of Cq. richardii can vary from 1.7 to 99.5%, with this species usually dominating in the main forest–steppe and steppe landscapes of the rural part of the Novosibirsk region.
In this study, we used a metagenomic-sequencing method to identify the viromes in seven mosquito species collected in the vicinity of Novosibirsk. The metagenomic approach was used to identify the viral diversity in randomly collected mosquitoes. We identified 30 coding-complete viral genomes and viral-like partial sequences of capsid proteins and/or RdRps from the mosquitoes (Table 1). These sequences were classified as putative members of the orders Tymovirales and Picornavirales; the families Partitiviridae, Totiviridae, Tombusviridae, Iflaviridae, Nodaviridae, Permutotetraviridae, and Solemoviridae; and four unclassified RNA-viruses. The previously described Partitivirus-like 1, Hammarskog tombus-like virus, Hammarskog picorna-like virus, Lymantria dispar iflavirus 1, Wenzhou noda-like virus 6, Mayapan virus, Sanxia permutotetra-like virus 1, and Chaq virus-like 1 were identified as practically complete genomes with levels of identity of 82–99% with the Cq. richardii mosquito. These viruses were found earlier in Liberia (west Africa), Sweden (north Europe), the USA (America), China (Asia), and Mexico (Central America). These findings allowed us to hypothesize that these viruses may be widely distributed on a global scale.
Some novel putative viruses and viral sequences had prototype viral sequences with identity levels from 31% to 79%, with these prototypes also being found in invertebrates from almost all continents. Some of them were associated with different species of mosquitoes. In our study, the majority of the novel viruses were associated with the Cq. richardii mosquito, a species that is widespread in the south of western Siberia [45]. The role of this mosquito species in the spread of human viral infections in Siberia has been virtually uninvestigated, suggesting that our knowledge concerning mosquito-associated viruses in north Eurasia is very limited and requires further study.
5. Conclusions
We identified novel and known viral genomes and viral-like partial sequences in mosquitoes collected in the Novosibirsk region of western Siberia. They were classified as novel putative viruses via a bioinformatics analysis of partial sequences of capsid proteins and RdRps or whole polyproteins (genomes) within the orders Tymovirales and Picornavirales; the families Partitiviridae, Totiviridae, Tombusviridae, Iflaviridae, Nodaviridae, Permutotetraviridae, and Solemoviridae; and four unclassified RNA viruses. We believe that these virus identifications can enhance our understanding of the transmission of RNA viruses by mosquitoes in north Asia. We hope that the discovery and observation of these mosquito-borne viruses can help prevent future outbreaks of viral infections in the region under study.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/v15091896/s1, Figure S1: Andean potato mild mosaic virus (APMMV) as novel Inya insect-associated virus 1 and 2; Figure S2: A novel virus with 51% identity with Atrato partiti-like virus 2 was detected in the Anopheles messeae pool and referred to as the Krahall insect-associated virus; Figure S3: The scheme of genome organization for Fitzroy Crossing toti-like virus 1; Figure S4: The scheme of genome organization for Tombus-like virus; Figure S5: The genome organization for the picorna-like virus; Figure S6: The genome organization for nodaviruses (ssRNA); Figure S7: The genome organization for permutotetraviruses.
Author Contributions
V.A.T., A.N.S., M.Y.K., E.P.P., A.V.G. and V.B.L. designed the experiments and analyzed the data; A.N.S., M.Y.K., E.P.P., N.L.T., R.B.B., A.V.G., T.P.M., Y.A.K. and T.V.T. performed the experiments; V.A.T. and V.B.L. managed the investigation and obtained funds; V.A.T. and V.B.L. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Ministry of Science and Higher Education of the Russian Federation (Agreement No. 075-15-2019-1665) and as part of a state assignment (9/21) for the State Scientific Center of Virology and Biotechnology «Vector» of the Federal Service for Surveillance of Consumer Rights Protection and Human Welfare.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors have no conflict of interest related to this study. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Rizzoli, A.; Jimenez-Clavero, M.A.; Barzon, L.; Cordioli, P.; Figuerola, J.; Koraka, P.; Martina, B.; Moreno, A.; Nowotny, N.; Pardigon, N.; et al. The challenge of West Nile virus in Europe: Knowledge gaps and research priorities. Euro Surveill. 2015, 20, 21135. [Google Scholar] [CrossRef] [PubMed]
- Fauci, A.S.; Morens, D.M. Zika Virus in the Americas—Yet Another Arbovirus Threat. N. Engl. J. Med. 2016, 374, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.A.; Dermody, T.S. Chikungunya virus: Epidemiology, replication, disease mechanisms, and prospective intervention strategies. J. Clin. Investig. 2017, 127, 737–749. [Google Scholar] [CrossRef]
- Messina, J.P.; Brady, O.J.; Scott, T.W.; Zou, C.; Pigott, D.M.; Duda, K.A.; Bhatt, S.; Katzelnick, L.; Howes, R.E.; Battle, K.E.; et al. Global spread of dengue virus types: Mapping the 70 year history. Trends Microbiol. 2014, 22, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Sadeghi, M.; Altan, E.; Deng, X.; Barker, C.M.; Fang, Y.; Coey, L.L.; Delwart, E. Virome of >12 thousand Culex mosquitoes from throughout California. Virology 2018, 523, 74–88. [Google Scholar] [CrossRef]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- Frey, K.G.; Biser, T.; Hamilton, T.; Santos, C.J.; Pimentel, G.; Mokashi, V.P.; Bishop-Lilly, K.A. Bioinformatic Characterization of Mosquito Viromes within the Eastern United States and Puerto Rico: Discovery of Novel Viruses. Evol. Bioinform. 2016, 12, 1–12. [Google Scholar] [CrossRef]
- Junglen, S.; Drosten, C. Virus discovery and recent insights into virus diversity in arthropods. Curr. Opin. Microbiol. 2013, 16, 507–513. [Google Scholar] [CrossRef]
- Webster, C.L.; Longdon, B.; Lewis, S.H.; Obbard, D.J. Twenty-Five New Viruses Associated with the Drosophilidae (Diptera). Evol. Bioinform. 2016, 12, 13–25. [Google Scholar] [CrossRef]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Yam, A.W.; Lu, J.W.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; et al. A new insect-specific flavivirus from northern Australia suppresses replication of West Nile virus and Murray Valley encephalitis virus in co-infected mosquito cells. PLoS ONE 2013, 8, e56534. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.L.; Solberg, O.D.; Langevin, S.A.; Brault, A.C. Characterization of a novel insect-specific flavivirus from Brazil: Potential for inhibition of infection of arthropod cells with medically important flaviviruses. J. Gen. Virol. 2014, 95, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Goenaga, S.; Kenney, J.L.; Duggal, N.K.; Delorey, M.; Ebel, G.D.; Zhang, B.; Levis, S.C.; Enria, D.A.; Brault, A.C. Potential for Co-Infection of a Mosquito-Specific Flavivirus, Nhumirim Virus, to Block West Nile Virus Transmission in Mosquitoes. Viruses 2015, 7, 5801–5812. [Google Scholar] [CrossRef]
- Nasar, F.; Erasmus, J.H.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. Eilat virus induces both homologous and heterologous interference. Virology 2015, 484, 51–58. [Google Scholar] [CrossRef]
- Hall-Mendelin, S.; McLean, B.J.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Hall, R.A.; van den Hurk, A.F. The insect-specific Palm Creek virus modulates West Nile virus infection in and transmission by Australian mosquitoes. Parasit. Vectors 2016, 9, 414. [Google Scholar] [CrossRef]
- Marklewitz, M.; Zirkel, F.; Kurth, A.; Drosten, C.; Junglen, S. Evolutionary and phenotypic analysis of live virus isolates suggests arthropod origin of a pathogenic RNA virus family. Proc. Natl. Acad. Sci. USA 2015, 112, 7536–7541. [Google Scholar] [CrossRef]
- Crochu, S.; Cook, S.; Attoui, H.; Charrel, R.N.; De Chesse, R.; Belhouchet, M.; Lemasson, J.J.; de Micco, P.; de Lamballerie, X. Sequences of flavivirus-related RNA viruses persist in DNA form integrated in the genome of Aedes spp. mosquitoes. J. Gen. Virol. 2004, 85, 1971–1980. [Google Scholar] [CrossRef]
- Cholleti, H.; Hayer, J.; Abilio, A.P.; Mulandane, F.C.; Verner-Carlsson, J.; Falk, K.I.; Fafetine, J.M.; Berg, M.; Blomstrom, A.L. Discovery of Novel Viruses in Mosquitoes from the Zambezi Valley of Mozambique. PLoS ONE 2016, 11, e0162751. [Google Scholar] [CrossRef]
- Shi, M.; Neville, P.; Nicholson, J.; Eden, J.S.; Imrie, A.; Holmes, E.C. High-Resolution Metatranscriptomics Reveals the Ecological Dynamics of Mosquito-Associated RNA Viruses in Western Australia. J. Virol. 2017, 91, e00680-17. [Google Scholar] [CrossRef]
- Ternovoĭ, V.A.; Shchelkanov, M.I.; Shestopalov, A.M.; Aristova, V.A.; Protopopova, E.V.; Gromashevskiĭ, V.L.; Druziaka, A.V.; Slavskiĭ, A.A.; Zolotykh, S.I.; Loktev, V.B.; et al. Detection of West Nile virus in birds in the territories of Baraba and Kulunda lowlands (West Siberian migration way) during summer-autumn of 2002. Vopr. Virusol. 2004, 49, 52–56. (In Russian) [Google Scholar] [PubMed]
- Korobitsyn, I.G.; Moskvitina, N.S.; Tyutenkov, O.Y.; Gashkov, S.I.; Kononova, Y.; Moskvitin, S.S.; Romanenko, V.N.; Mikryukova, T.P.; Protopopova, E.V.; Kartashov, M.Y.; et al. Detection of tick-borne pathogens in wild birds and their ticks in Western Siberia and high level of their mismatch. Folia Parasitol. 2021, 68, 024. [Google Scholar] [CrossRef] [PubMed]
- Ternovoi, V.A.; Shvalov, A.N.; Kartashov, M.Y.; Ponomareva, E.P.; Tupota, N.L.; Khoroshavin, Y.A.; Bayandin, R.B.; Gladysheva, A.V.; Mikryukova, T.P.; Tregubchak, T.V. Cases of West Nile fever in Novosibirsk region in 2004, and the genotyping of its viral pathogen. Vestn. Ross. Akad. Med. Nauk. 2007, 1, 21–26. (In Russian) [Google Scholar]
- Rodgers, M.A.; Wilkinson, E.; Vallari, A.; McArthur, C.; Sthreshley, L.; Brennan, C.A.; Cloherty, G.; de Oliveira, T. Sensitive Next-Generation Sequencing Method Reveals Deep Genetic Diversity of HIV-1 in the Democratic Republic of the Congo. J. Virol. 2017, 91, e01841-16. [Google Scholar] [CrossRef]
- Ding, S.W.; Howe, J.; Keese, P.; Mackenzie, A.; Meek, D.; Osorio-Keese, M.; Skotnicki, M.; Srifah, P.; Torronen, M.; Gibbs, A. The tymobox, a sequence shared by most tymoviruses: Its use in molecular studies of tymoviruses. Nucleic Acids Res. 1990, 18, 1181–1187. [Google Scholar] [CrossRef]
- Kreuze, J.; Koenig, R.; De Souza, J.; Vetten, H.J.; Muller, G.; Flores, B.; Ziebell, H.; Cuellar, W. The complete genome sequences of a Peruvian and a Colombian isolate of Andean potato latent virus and partial sequences of further isolates suggest the existence of two distinct potato-infecting tymovirus species. Virus Res. 2013, 173, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Carino, E.; Bera, S.; Gao, F.; May, J.P.; Simon, A.E. Structural Analysis and Whole Genome Mapping of a New Type of Plant Virus Subviral RNA: Umbravirus-Like Associated RNAs. Viruses 2021, 13, 646. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, S.; Käfer, S.; Zirkel, F.; Donath, A.; Petersen, M.; Liu, S.; Zhou, X.; Drosten, C.; Misof, B.; Junglen, S. Viromics of extant insect orders unveil the evolution of the flavi-like superfamily. Virus Evol. 2021, 7, veab030. [Google Scholar] [CrossRef]
- Bera, S.; Ilyas, M.; Mikkelsen, A.A.; Simon, A.E. Conserved Structure Associated with Different 3’CITEs Is Important for Translation of Umbraviruses. Viruses 2023, 15, 638. [Google Scholar] [CrossRef]
- Öhlund, P.; Hayer, J.; Lundén, H.; Hesson, J.C.; Blomström, A.L. Viromics Reveal a Number of Novel RNA Viruses in Swedish Mosquitoes. Viruses 2019, 11, 1027. [Google Scholar] [CrossRef]
- Sanborn, M.A.; Klein, T.A.; Kim, H.-C.; Fung, C.K.; Figueroa, K.L.; Yang, Y.; Asafo-Adjei, E.A.; Jarman, R.G.; Hang, J. Metagenomic Analysis Reveals Three Novel and Prevalent Mosquito Viruses from a Single Pool of Aedes vexans nipponii Collected in the Republic of Korea. Viruses 2019, 11, 222. [Google Scholar] [CrossRef]
- Walker, P.J.; Firth, C.; Widen, S.G.; Blasdell, K.R.; Guzman, H.; Wood, T.G.; Paradkar, P.N.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N. Evolution of genome size and complexity in the Rhabdoviridae. PLoS Pathog. 2015, 11, e1004664. [Google Scholar] [CrossRef]
- Presti, R.M.; Zhao, G.; Beatty, W.L.; Mihindukulasuriya, K.A.; da Rosa, A.P.T.; Popov, V.L.; Wang, D. Quaranfil, Johnston Atoll, and Lake Chad viruses are novel members of the family Orthomyxoviridae. J. Virol. 2009, 83, 11599–11606. [Google Scholar] [CrossRef]
- Xia, H.; Wang, Y.; Atoni, E.; Zhang, B.; Yuan, Z. Mosquito-associated viruses in China. Virol. Sin. 2018, 33, 5–20. [Google Scholar] [CrossRef]
- Attoui, H.; Jaafar, F.M.; Belhouchet, M.; Tao, S.; Chen, B.; Liang, G.; de Lamballerie, X. Liao ning virus, a new Chinese seadornavirus that replicates in transformed and embryonic mammalian cells. J. Gen. Virol. 2006, 87, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Forrester, N.L.; Palacios, G.; Nasar, F.; Savji, N.; Rossi, S.L.; Guzman, H.; Wood, T.G.; Popov, V.; Gorchakov, R.; et al. Negevirus: A proposed new taxon of insect-specific viruses with wide geographic distribution. J. Virol. 2013, 87, 2475–2488. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Han, J.; Zhang, Y.; Li, C.; Guo, X.; Wen, S.; Tian, M.; Li, Y.; Wang, M.; Liu, H.; et al. Metagenomic Analysis of Flaviviridae in Mosquito Viromes Isolated from Yunnan Province in China Reveals Genes from Dengue and Zika Viruses. Front. Cell. Infect. Microbiol. 2018, 8, 359. [Google Scholar] [CrossRef]
- Klungthong, C.; Gibbons, R.V.; Thaisomboonsuk, B.; Nisalak, A.; Kalayanarooj, S.; Thirawuth, V.; Jarman, R.G. Dengue virus detection using whole blood for reverse transcriptase PCR and virus isolation. J. Clin. Microbiol. 2007, 45, 2480–2485. [Google Scholar] [CrossRef]
- LaBeaud, A.D.; Bashir, F.; King, C.H. Measuring the burden of arboviral diseases: The spectrum of morbidity and mortality from four prevalent infections. Popul. Health Metr. 2011, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.A.; Bielefeldt-Ohmann, H.; McLean, B.J.; O’Brien, C.A.; Colmant, A.M.; Piyasena, T.B.; Harrison, J.J.; Newton, N.D.; Barnard, R.T.; Prow, N.A.; et al. Commensal Viruses of Mosquitoes: Host Restriction, Transmission, and Interaction with Arboviral Pathogens. Evol. Bioinform. 2017, 12, 35–44. [Google Scholar] [CrossRef]
- Vasilakis, N.; Tesh, R.B. Insect-specific viruses and their potential impact on arbovirus transmission. Curr. Opin. Virol. 2015, 15, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; de Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Powers, A.M.; Waterman, S.H. A decade of arboviral activity—Lessons learned from the trenches. PLoS Negl. Trop. Dis. 2017, 11, e0005421. [Google Scholar] [CrossRef]
- Joganzen, B.G.; Petkevich, A.N. Acclimatization of fish in Western Siberia. Proc. Baraba Branch VNIORKH 1951, 5, 3–204. [Google Scholar]
- IuV, K.; Mirzaeva, A.G.; IuV, S.; Protopopova, E.V.; Dupal, T.A.; Ternovoĭ, V.A.; IuA, I.; Shestopalov, A.M.; Loktev, V.B. Species composition of mosquitoes (Diptera, Culicidae) and possibility of the West Nile virus natural foci formation in the South of Western Siberia. Parazitologiia 2007, 41, 459–470. (In Russian) [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).