
Tumor Necrosis Factor and Interleukin-1β Upregulate NRP2 Expression and Promote SARS-CoV-2 Proliferation
 , , , ,
, , , ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Next-Generation RNA Sequencing (RNA-seq)
2.3. Preparation of Primary Cultured Fibroblast-like Synoviocytes (FLS), the MH7A Cell Line and the Lung MRC5 Cell Line
2.4. RNA Isolation and Quantitative Real-Time PCR (RT–qPCR)
2.5. Immunohistochemistry (IHC) Staining for Formalin-Fixed Paraffin-Embedded (FFPE) Synovial Tissues
2.6. Luciferase Assay
2.7. Western Blotting
2.8. Plasmid Construction and Transfection into MH7A Cell Line
2.9. Synthesis of Recombinant S Protein
2.10. S Protein Binding In Vitro Assay
2.11. SARS-CoV-2 Infection of MH7A Cells
2.12. Statistical Analysis
3. Results
3.1. SARS-CoV-2 Host Receptor Gene Expression in Inflamed Synovial Tissues
3.2. Identification of Cytokines That Can Enhance NRP1 and NRP2 Expressions
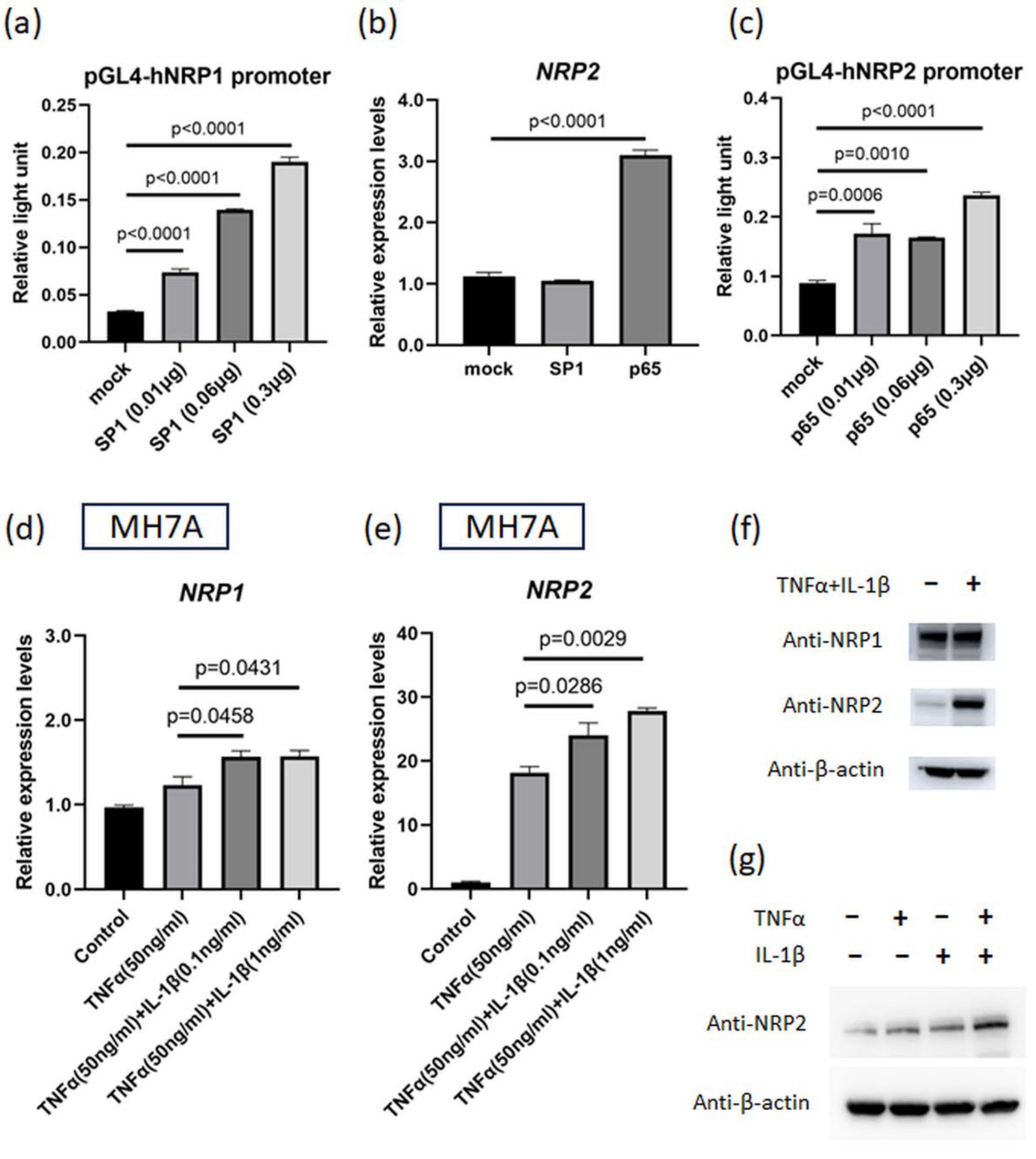
3.3. NF-κB p65 Transcription Factor Can Induce NRP2 Expression
3.4. NRP2 Expression Is Elevated by TNFα and IL-1β Co-Stimulation
3.5. SARS-CoV-2 Infection and Proliferation under Inflammatory Conditions
3.6. Proinflammatory Cytokine-Induced NRP2 Expression Contributes to Enhancement of Viral Proliferation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus–Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Mcgoogan, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72,314 Cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Clinical Trial Group. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Kalil, A.C.; Patterson, T.F.; Mehta, A.K.; Tomashek, K.M.; Wolfe, C.R.; Ghazaryan, V.; Marconi, V.C.; Ruiz-Palacios, G.M.; Hsieh, L.; Kline, S.; et al. ACTT-2 Study Group Members. Baricitinib plus Remdesivir for Hospitalized Adults with COVID-19. N. Engl. J. Med. 2021, 384, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Kinoshita, N.; Yamayoshi, S.; Sakai-Tagawa, Y.; Fujisaki, S.; Ito, M.; Iwatsuki-Horimoto, K.; Chiba, S.; Halfmann, P.; Nagai, H.; et al. Efficacy of Antibodies and Antiviral Drugs Against COVID-19 Omicron Variant. N. Engl. J. Med. 2022, 386, 995–998. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; Van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam Van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and Furin Are Both Essential for Proteolytic Activation of SARS-CoV-2 in Human Airway Cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef]
- Kumanogoh, A.; Kikutani, H. Immunological Functions of the Neuropilins and Plexins as Receptors for Semaphorins. Nat. Rev. Immunol. 2013, 13, 802–814. [Google Scholar] [CrossRef]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.E.; Williamson, M.K.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 Is a Host Factor for SARS-CoV-2 Infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An Inflammatory Cytokine Signature Predicts COVID-19 Severity and Survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The Cytokine Storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef]
- Laguna-Goya, R.; Utrero-Rico, A.; Talayero, P.; Lasa-Lazaro, M.; Ramirez-Fernandez, A.; Naranjo, L.; Segura-Tudela, A.; Cabrera-Marante, O.; Rodriguez De Frias, E.; Garcia-Garcia, R.; et al. IL-6–Based Mortality Risk Model for Hospitalized Patients with COVID-19. J. Allergy Clin. Immunol. 2020, 146, 799–807.e9. [Google Scholar] [CrossRef]
- Abani, O.; Abbas, A.; Abbas, F.; Abbas, M.; Abbasi, S.; Abbass, H.; Abbott, A.; Abdallah, N.; Abdelaziz, A.; Abdelfattah, M.; et al. Tocilizumab in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. Lancet 2021, 397, 1637–1645. [Google Scholar] [CrossRef]
- REMAP-CAP Investigators; Gordon, A.C.; Mouncey, P.R.; Al-Beidh, F.; Rowan, K.M.; Nichol, A.D.; Arabi, Y.M.; Annane, D.; Beane, A.; van Bentum-Puijk, W.; et al. Interleukin-6 Receptor Antagonists in Critically Ill Patients with COVID-19. N. Engl. J. Med. 2021, 384, 1491–1502. [Google Scholar] [CrossRef]
- Gladman, D.D.; Mease, P.J.; Ritchlin, C.T.; Choy, E.H.S.; Sharp, J.T.; Ory, P.A.; Perdok, R.J.; Sasso, E.H. Adalimumab for Long-Term Treatment of Psoriatic Arthritis: Forty-Eight Week Data from the Adalimumab Effectiveness in Psoriatic Arthritis Trial. Arthritis Rheum. 2007, 56, 476–488. [Google Scholar] [CrossRef]
- Reinisch, W.; Sandborn, W.J.; Hommes, D.W.; D’haens, G.; Hanauer, S.; Schreiber, S.; Panaccione, R.; Fedorak, R.N.; Tighe, M.B.; Huang, B.; et al. Adalimumab for Induction of Clinical Remission in Moderately to Severely Active Ulcerative Colitis: Results of a Randomised Controlled Trial. Gut 2011, 60, 780–787. [Google Scholar] [CrossRef]
- Sugimura, N.; Mizoshita, T.; Sugiyama, T.; Togawa, S.; Miyaki, T.; Suzuki, T.; Tanida, S.; Kataoka, H.; Sasaki, M. Real-World Efficacy of Adalimumab and Infliximab for Refractory Intestinal Behcet’s Disease. Dig. Liver Dis. 2019, 51, 967–971. [Google Scholar] [CrossRef]
- Van De Putte, L.B.A.; Atkins, C.; Malaise, M.; Sany, J.; Russell, A.S.; van Riel, P.L.; Settas, L.; Bijlsma, J.W.; Todesco, S.; Dougados, M.; et al. Efficacy and Safety of Adalimumab as Monotherapy in Patients with Rheumatoid Arthritis for Whom Previous Disease Modifying Antirheumatic Drug Treatment Has Failed. Ann. Rheum. Dis. 2004, 63, 508–516. [Google Scholar] [CrossRef]
- Van Der Heijde, D.; Kivitz, A.; Schiff, M.H.; Sieper, J.; Dijkmans, B.A.C.; Braun, J.; Dougados, M.; Reveille, J.D.; Wong, R.L.; Kupper, H.; et al. Efficacy and Safety of Adalimumab in Patients with Ankylosing Spondylitis: Results of a Multicenter, Randomized, Double-Blind, Placebo-Controlled Trial. Arthritis Rheum. 2006, 54, 2136–2146. [Google Scholar] [CrossRef]
- Gianfrancesco, M.; Hyrich, K.L.; Al-Adely, S.; Carmona, L.; Danila, M.I.; Gossec, L.; Izadi, Z.; Jacobsohn, L.; Katz, P.; Lawson-Tovey, S.; et al. COVID-19 Global Rheumatology Alliance. Characteristics Associated with Hospitalisation for COVID-19 in People with Rheumatic Disease: Data from the COVID-19 Global Rheumatology Alliance Physician-Reported Registry. Ann. Rheum. Dis. 2020, 79, 859–866. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; De Sá, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Gonçalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes Are Activated in Response to SARS-CoV-2 Infection and Are Associated with COVID-19 Severity in Patients. J. Exp. Med. 2021, 218, e20201707. [Google Scholar] [CrossRef]
- Mokuda, S.; Tokunaga, T.; Masumoto, J.; Sugiyama, E. Angiotensin-Converting Enzyme 2, a SARS-CoV-2 Receptor, Is Upregulated by Interleukin 6 through STAT3 Signaling in Synovial Tissues. J. Rheumatol. 2020, 47, 1593–1595. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Mokuda, S.; Tokunaga, T.; Kohno, H.; Ishitoku, M.; Araki, K.; Sugimoto, T.; Yoshida, Y.; Yamamoto, T.; Matsumoto, M.; et al. Expression of Factor XIII Originating from Synovial Fibroblasts and Macrophages Induced by Interleukin-6 Signaling. Inflamm. Regen. 2023, 43, 2. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, T.; Mokuda, S.; Kohno, H.; Yukawa, K.; Kuranobu, T.; Oi, K.; Yoshida, Y.; Hirata, S.; Sugiyama, E. TGFβ1 Regulates Human RANKL-Induced Osteoclastogenesis via Suppression of NFATc1 Expression. Int. J. Mol. Sci. 2020, 21, 800. [Google Scholar] [CrossRef] [PubMed]
- Mokuda, S.; Miyazaki, T.; Ubara, Y.; Kanno, M.; Sugiyama, E.; Takasugi, K.; Masumoto, J. CD1a+ Survivin+ Dendritic Cell Infiltration in Dermal Lesions of Systemic Sclerosis. Arthritis Res. Ther. 2015, 17, 275. [Google Scholar] [CrossRef]
- Nomura, T.; Nazmul, T.; Yoshimoto, R.; Higashiura, A.; Oda, K.; Sakaguchi, T. Ethanol Susceptibility of SARS-CoV-2 and Other Enveloped Viruses. Biocontrol Sci. 2021, 26, 177–180. [Google Scholar] [CrossRef]
- Rossignol, M.; Pouysségur, J.; Klagsbrun, M. Characterization of the neuropilin-1 Promoter; Gene Expression Is Mediated by the Transcription Factor Sp1. J. Cell. Biochem. 2003, 88, 744–757. [Google Scholar] [CrossRef]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H.; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef]
- Lukassen, S.; Chua, R.L.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-CoV-2 Receptor ACE2 and TMPRSS2 Are Primarily Expressed in Bronchial Transient Secretory Cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef]
- Jobe, A.; Vijayan, R. Neuropilins: C-End Rule Peptides and Their Association with Nociception and COVID-19. Comput. Struct. Biotechnol. J. 2021, 19, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Lin, B.; Pathak, J.L.; Gao, H.; Young, A.J.; Wang, X.; Liu, C.; Wu, K.; Liu, M.; Chen, J.M.; et al. ACE2 and Furin Expressions in Oral Epithelial Cells Possibly Facilitate COVID-19 Infection via Respiratory and Fecal-Oral Routes. Front. Med. 2020, 7, 580796. [Google Scholar] [CrossRef] [PubMed]
- Stein, S.R.; Ramelli, S.C.; Grazioli, A.; Chung, J.Y.; Singh, M.; Yinda, C.K.; Winkler, C.W.; Sun, J.; Dickey, J.M.; Ylaya, K.; et al. SARS-CoV-2 Infection and Persistence in the Human Body and Brain at Autopsy. Nature 2022, 612, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, L.; Mescia, F.; Turner, L.; Hanson, A.L.; Kotagiri, P.; Dunmore, B.J.; Ruffieux, H.; De Sa, A.; Huhn, O.; Morgan, M.D.; et al. Longitudinal Analysis Reveals That Delayed Bystander CD8+ T Cell Activation and Early Immune Pathology Distinguish Severe COVID-19 from Mild Disease. Immunity 2021, 54, 1257–1275.e8. [Google Scholar] [CrossRef]
- Haga, S.; Yamamoto, N.; Nakai-Murakami, C.; Osawa, Y.; Tokunaga, K.; Sata, T.; Yamamoto, N.; Sasazuki, T.; Ishizaka, Y. Modulation of TNF-α-Converting Enzyme by the Spike Protein of SARS-CoV and ACE2 Induces TNF-α Production and Facilitates Viral Entry. Proc. Natl. Acad. Sci. USA 2008, 105, 7809–7814. [Google Scholar] [CrossRef]
- Zipeto, D.; Palmeira, J.D.F.; Argañaraz, G.A.; Argañaraz, E.R. ACE2/ADAM17/TMPRSS2 Interplay May Be the Main Risk Factor for COVID-19. Front. Immunol. 2020, 11, 576745. [Google Scholar] [CrossRef]
- Séguin, C.A.; Pilliar, R.M.; Madri, J.A.; Kandel, R.A. TNF-alpha Induces MMP2 Gelatinase Activity and MT1-MMP Expression in an In Vitro Model of Nucleus Pulposus Tissue Degeneration. Spine 2008, 33, 356–365. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishitoku, M.; Mokuda, S.; Araki, K.; Watanabe, H.; Kohno, H.; Sugimoto, T.; Yoshida, Y.; Sakaguchi, T.; Masumoto, J.; Hirata, S.; et al. Tumor Necrosis Factor and Interleukin-1β Upregulate NRP2 Expression and Promote SARS-CoV-2 Proliferation. Viruses 2023, 15, 1498. https://doi.org/10.3390/v15071498
Ishitoku M, Mokuda S, Araki K, Watanabe H, Kohno H, Sugimoto T, Yoshida Y, Sakaguchi T, Masumoto J, Hirata S, et al. Tumor Necrosis Factor and Interleukin-1β Upregulate NRP2 Expression and Promote SARS-CoV-2 Proliferation. Viruses. 2023; 15(7):1498. https://doi.org/10.3390/v15071498
Chicago/Turabian StyleIshitoku, Michinori, Sho Mokuda, Kei Araki, Hirofumi Watanabe, Hiroki Kohno, Tomohiro Sugimoto, Yusuke Yoshida, Takemasa Sakaguchi, Junya Masumoto, Shintaro Hirata, and et al. 2023. "Tumor Necrosis Factor and Interleukin-1β Upregulate NRP2 Expression and Promote SARS-CoV-2 Proliferation" Viruses 15, no. 7: 1498. https://doi.org/10.3390/v15071498
APA StyleIshitoku, M., Mokuda, S., Araki, K., Watanabe, H., Kohno, H., Sugimoto, T., Yoshida, Y., Sakaguchi, T., Masumoto, J., Hirata, S., & Sugiyama, E. (2023). Tumor Necrosis Factor and Interleukin-1β Upregulate NRP2 Expression and Promote SARS-CoV-2 Proliferation. Viruses, 15(7), 1498. https://doi.org/10.3390/v15071498