

Booster Dose of SARS-CoV-2 mRNA Vaccine in Kidney Transplanted Patients Induces Wuhan-Hu-1 Specific Neutralizing Antibodies and T Cell Activation but Lower Response against Omicron Variant
 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
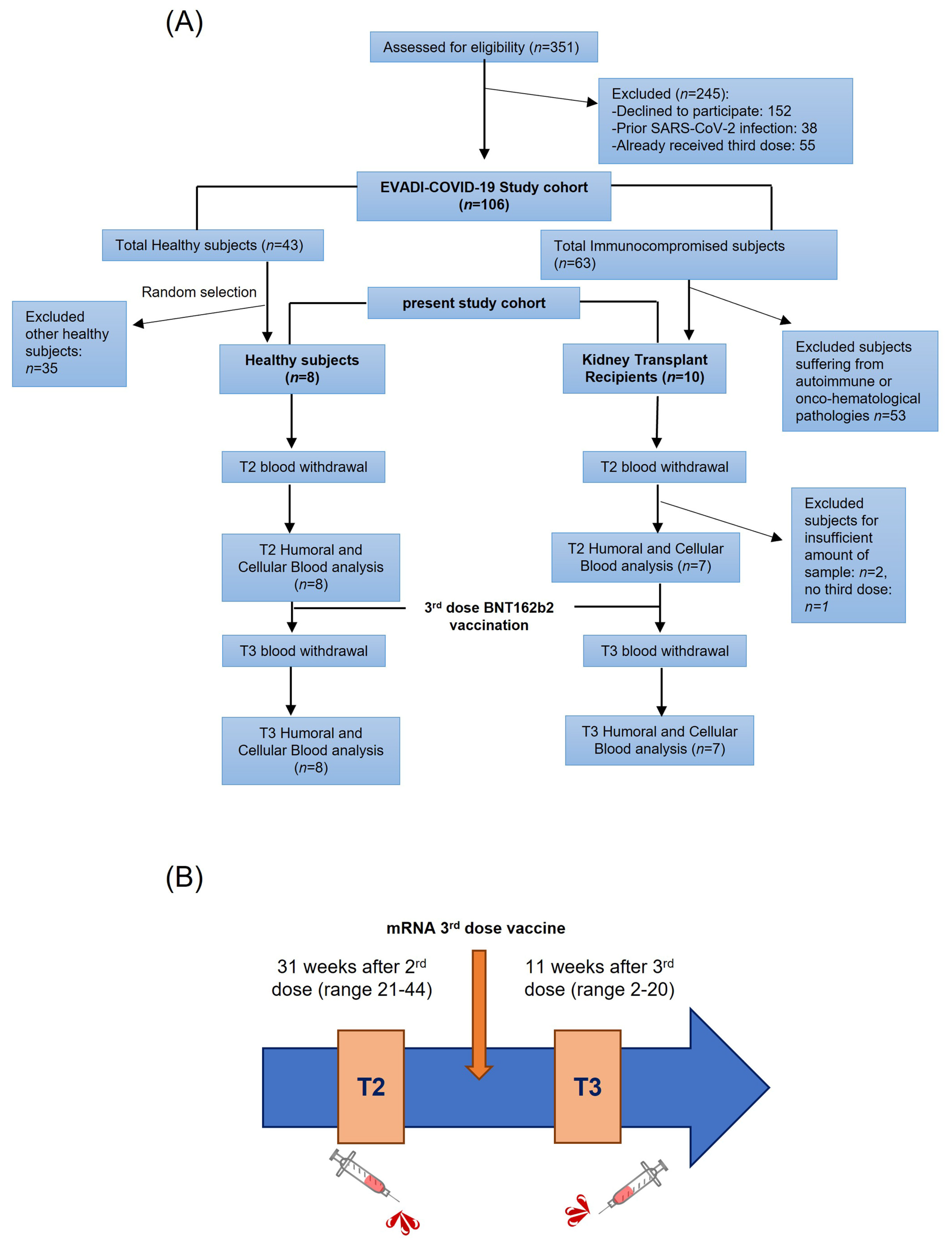
2.1. Cohort Description and Sample Collection
2.2. Total Anti-SARS-CoV-2 Ig Detection
2.3. Neutralization Assay
2.4. Functional T Cell Assays: Activation Induced Markers (AIM) and IFN-γ ELISA
2.5. Responsivity Criteria and Statistical Analysis
3. Results
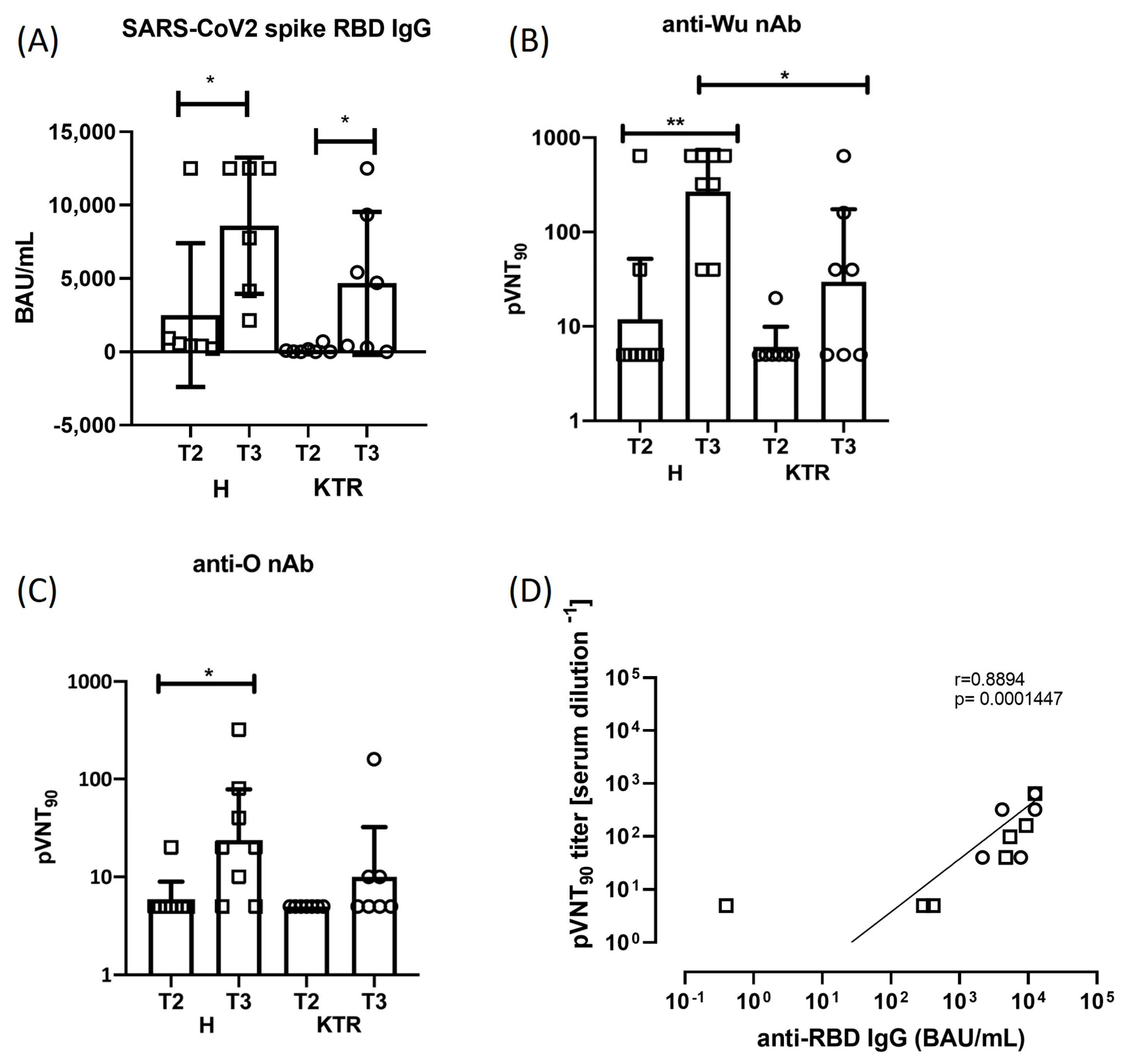
3.1. Total Antibody Response to SARS-CoV-2
3.2. Neutralizing Antibody Response against SARS-CoV-2 Wuhan-Hu-1 and Omicron Strains
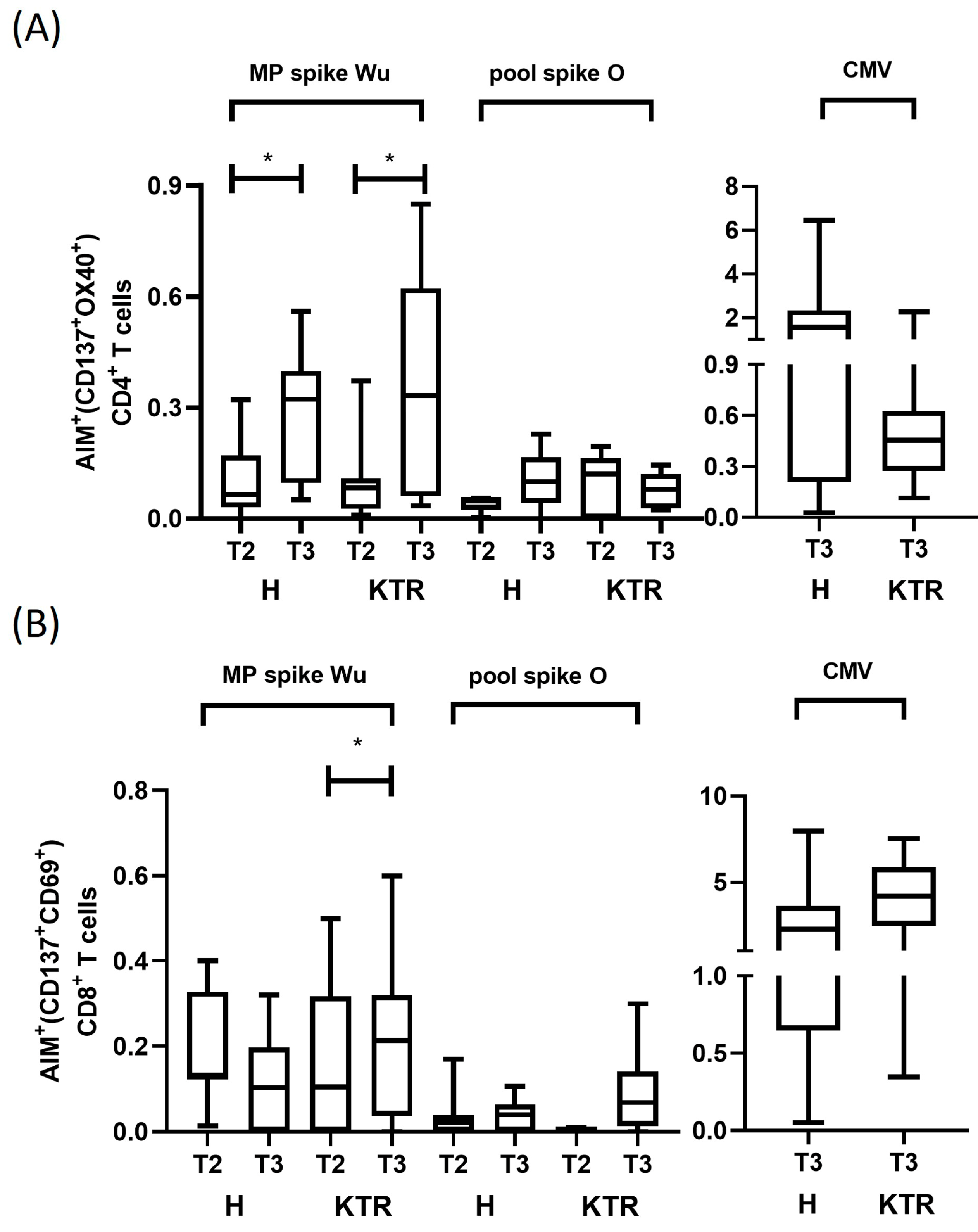
3.3. Analysis of Spike-Specific CD4+ and CD8+ T Cell Reactivity
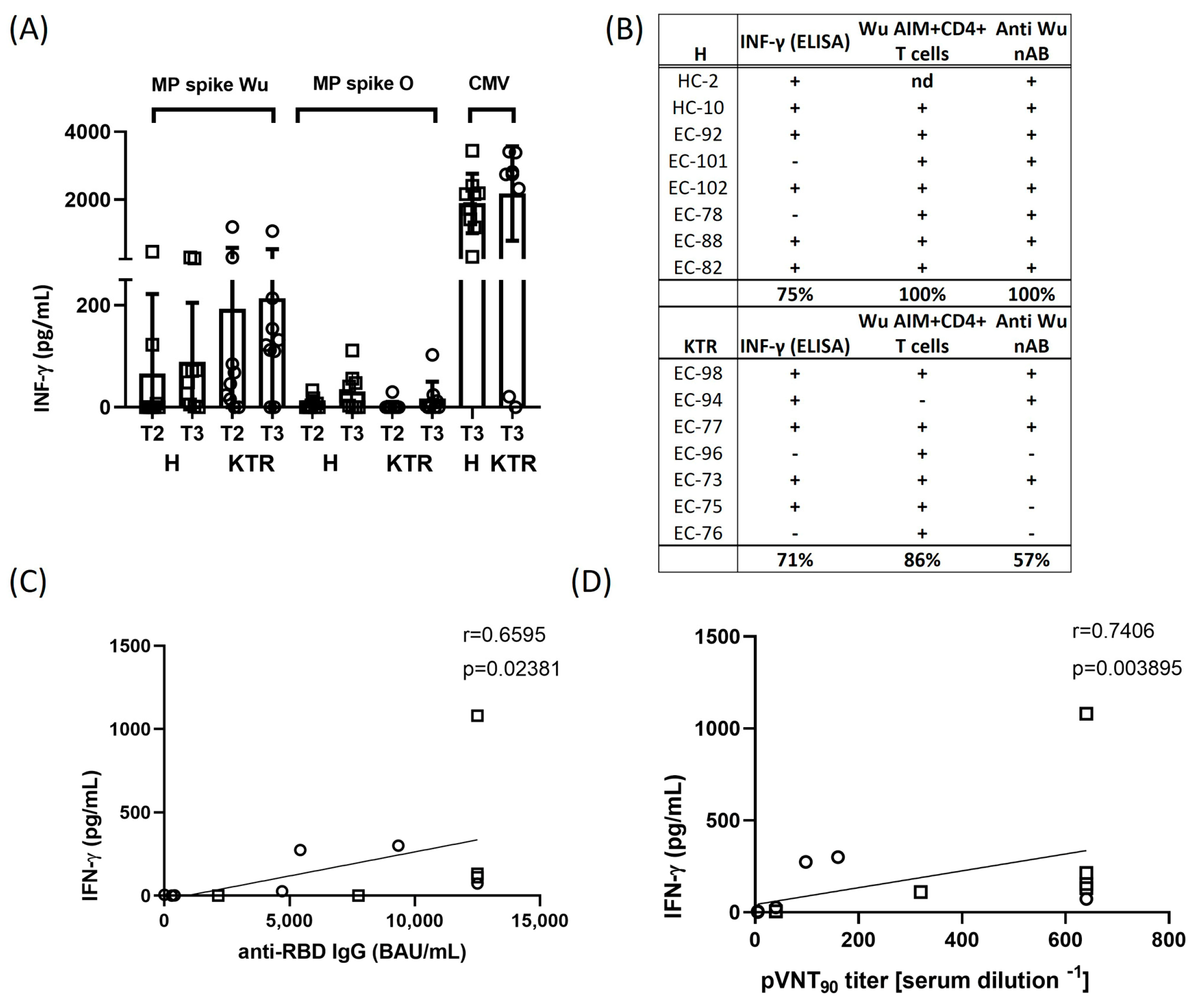
3.4. IFN-γ Production in Response to Spike Peptides and Correlation Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chu, L.; Vrbicky, K.; Montefiori, D.; Huang, W.; Nestorova, B.; Chang, Y.; Carfi, A.; Edwards, D.K.; Oestreicher, J.; Legault, H.; et al. Immune Response to SARS-CoV-2 after a Booster of MRNA-1273: An Open-Label Phase 2 Trial. Nat. Med. 2022, 28, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Geers, D.; Shamier, M.C.; Bogers, S.; den Hartog, G.; Gommers, L.; Nieuwkoop, N.N.; Schmitz, K.S.; Rijsbergen, L.C.; van Osch, J.A.T.; Dijkhuizen, E.; et al. SARS-CoV-2 Variants of Concern Partially Escape Humoral but Not T-Cell Responses in COVID-19 Convalescent Donors and Vaccinees. Sci. Immunol. 2021, 6, eabj1750. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, T.; Ferguson, N.M.; Nash, S.G.; Webster, H.H.; Flaxman, S.; Andrews, N.; Hinsley, W.; Bernal, J.L.; Kall, M.; Bhatt, S.; et al. Comparative Analysis of the Risks of Hospitalisation and Death Associated with SARS-CoV-2 Omicron (B.1.1.529) and Delta (B.1.617.2) Variants in England: A Cohort Study. Lancet 2022, 399, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Coelho, C.H.; Zhang, Z.; Dan, J.M.; Yu, E.D.; Methot, N.; Bloom, N.I.; Goodwin, B.; Phillips, E.; Mallal, S.; et al. SARS-CoV-2 Vaccination Induces Immunological T Cell Memory Able to Cross-Recognize Variants from Alpha to Omicron. Cell 2022, 185, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Mateus, J.; Dan, J.M.; Zhang, Z.; Moderbacher, C.R.; Lammers, M.; Goodwin, B.; Sette, A.; Crotty, S.; Weiskopf, D. Low-Dose MRNA-1273 COVID-19 Vaccine Generates Durable Memory Enhanced by Cross-Reactive T Cells. Science 2021, 374, eabj9853. [Google Scholar] [CrossRef] [PubMed]
- GeurtsvanKessel, C.H.; Geers, D.; Schmitz, K.S.; Mykytyn, A.Z.; Lamers, M.M.; Bogers, S.; Scherbeijn, S.; Gommers, L.; Sablerolles, R.S.G.; Nieuwkoop, N.N.; et al. Divergent SARS-CoV-2 Omicron-Reactive T and B Cell Responses in COVID-19 Vaccine Recipients. Sci. Immunol. 2022, 7, eabo2202. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against Type I IFNs in Patients with Life-Threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, L. Risk of COVID-19 for Patients with Cancer. Lancet Oncol. 2020, 21, e181. [Google Scholar] [CrossRef]
- Gao, Y.-D.; Ding, M.; Dong, X.; Zhang, J.-J.; Azkur, A.K.; Azkur, D.; Gan, H.; Sun, Y.-L.; Fu, W.; Li, W.; et al. Risk Factors for Severe and Critically Ill COVID-19 Patients: A Review. Allergy Eur. J. Allergy Clin. Immunol. 2021, 76, 428–455. [Google Scholar] [CrossRef]
- Yu, E.D.; Narowski, T.M.; Wang, E.; Garrigan, E.; Mateus, J.; Frazier, A.; Weiskopf, D.; Grifoni, A.; Premkumar, L.; da Silva Antunes, R.; et al. Immunological Memory to Common Cold Coronaviruses Assessed Longitudinally over a Three-Year Period. SSRN Electron. J. 2022, 30, 1269–1278. [Google Scholar] [CrossRef]
- Banerjee, D.; Popoola, J.; Shah, S.; Ster, I.C.; Quan, V.; Phanish, M. COVID-19 Infection in Kidney Transplant Recipients. Kidney Int. 2020, 97, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Rincon-Arevalo, H.; Choi, M.; Stefanski, A.L.; Halleck, F.; Weber, U.; Szelinski, F.; Jahrsdörfer, B.; Schrezenmeier, H.; Ludwig, C.; Sattler, A.; et al. Impaired Humoral Immunity to SARS-CoV-2 BNT162b2 Vaccine in Kidney Transplant Recipients and Dialysis Patients. Sci. Immunol. 2021, 6, eabj1031. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, J.; Tonnus, W.; Paliege, A.; Rettig, R.; Steglich, A.; Gembardt, F.; Kessel, F.; Kröger, H.; Arndt, P.; Sradnick, J.; et al. Cellular and Humoral Immune Responses after 3 Doses of BNT162b2 MRNA SARS-CoV-2 Vaccine in Kidney Transplant. Transplantation 2021, 105, e267–e269. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Abravanel, F.; Marion, O.; Couat, C.; Izopet, J.; Del Bello, A. Three Doses of an MRNA Covid-19 Vaccine in Solid-Organ Transplant Recipients. N. Engl. J. Med. 2021, 385, 661–662. [Google Scholar] [CrossRef] [PubMed]
- Hod, T.; Ben-David, A.; Olmer, L.; Scott, N.; Ghinea, R.; Mor, E.; Levy, I.; Indenbaum, V.; Lustig, Y.; Grossman, E.; et al. BNT162b2 Third Booster Dose Significantly Increases the Humoral Response Assessed by Both RBD IgG and Neutralizing Antibodies in Renal Transplant Recipients. Transpl. Int. 2022, 35, 10239. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Luo, D.; Mei, B.; Du, J.; Liu, X.; Xie, H.; Liu, L.; Su, S.; Mai, G. Immunogenicity of COVID-19 Vaccines in Solid Organ Transplant Recipients: A Systematic Review and Meta-Analysis. Clin. Microbiol. Infect. 2022, 29, 441–456. [Google Scholar] [CrossRef]
- Lyke, K.E.; Atmar, R.L.; Islas, C.D.; Posavad, C.M.; Szydlo, D.; Paul Chourdhury, R.; Deming, M.E.; Eaton, A.; Jackson, L.A.; Branche, A.R.; et al. Rapid Decline in Vaccine-Boosted Neutralizing Antibodies against SARS-CoV-2 Omicron Variant. Cell Rep. Med. 2022, 3, 100679. [Google Scholar] [CrossRef]
- Callaghan, C.J.; Curtis, R.M.K.; Mumford, L.; Whitaker, H.; Pettigrew, G.; Gardiner, D.; Marson, L.; Thorburn, D.; White, S.; Parmar, J.; et al. Vaccine Effectiveness against the SARS-CoV-2 B.1.1.529 Omicron Variant in Solid Organ and Islet Transplant Recipients in England: A National Retrospective Cohort Study. Transplantation 2023, 107, 1124–1135. [Google Scholar] [CrossRef]
- Kumar, D.; Ferreira, V.H.; Hall, V.G.; Hu, Q.; Samson, R.; Ku, T.; Ierullo, M.; Majchrzak-Kita, B.; Tomlinson, G.; Gingras, A.C.; et al. Neutralization of SARS-CoV-2 Variants in Transplant Recipients after Two and Three Doses of MRNA-1273 Vaccine. Ann. Intern. Med. 2022, 175, 226–233. [Google Scholar] [CrossRef]
- Benning, L.; Morath, C.; Bartenschlager, M.; Kim, H.; Reineke, M.; Beimler, J.; Buylaert, M.; Nusshag, C.; Kälble, F.; Reichel, P.; et al. Neutralizing Antibody Response against the B.1.617.2 (Delta) and the B.1.1.529 (Omicron) Variants after a Third MRNA SARS-CoV-2 Vaccine Dose in Kidney Transplant Recipients. Am. J. Transpl. 2022, 22, 1873–1883. [Google Scholar] [CrossRef]
- Takai, S.; Nishida, H.; Ito, H.; Fukuhara, H.; Nawano, T.; Narisawa, T.; Kanno, H.; Yagi, M.; Yamagishi, A.; Sakurai, T.; et al. Humoral and Cellular Immune Response and the Safety of Third SARS-CoV-2 MRNA Vaccine with Longer Interval after the Second Vaccination in Kidney Transplant Recipients. Front. Immunol. 2022, 13, 1050211. [Google Scholar] [CrossRef] [PubMed]
- Panizo, N.; Giménez, E.; Albert, E.; Zulaica, J.; Rodríguez-Moreno, A.; Rusu, L.; Giménez-Civera, E.; Puchades, M.J.; D’Marco, L.; Gandía-Salmerón, L.; et al. SARS-CoV-2-Spike Antibody and T-Cell Responses Elicited by a Homologous Third MRNA COVID-19 Dose in Hemodialysis and Kidney Transplant Recipients. Microorganisms 2022, 10, 2275. [Google Scholar] [CrossRef] [PubMed]
- Charmetant, X.; Espi, M.; Benotmane, I.; Barateau, V.; Heibel, F.; Buron, F.; Gautier-Vargas, G.; Delafosse, M.; Perrin, P.; Koenig, A.; et al. Infection or a Third Dose of MRNA Vaccine Elicits Neutralizing Antibody Responses against SARS-CoV-2 in Kidney Transplant Recipients. Sci. Transl. Med. 2022, 14, eabl6141. [Google Scholar] [CrossRef] [PubMed]
- Lederer, K.; Bettini, E.; Parvathaneni, K.; Painter, M.M.; Agarwal, D.; Lundgreen, K.A.; Weirick, M.; Muralidharan, K.; Castaño, D.; Goel, R.R.; et al. Germinal Center Responses to SARS-CoV-2 MRNA Vaccines in Healthy and Immunocompromised Individuals. Cell 2022, 185, 1008–1024. [Google Scholar] [CrossRef] [PubMed]
- Cucchiari, D.; Egri, N.; Bodro, M.; Herrera, S.; Del Risco-Zevallos, J.; Casals-Urquiza, J.; Cofan, F.; Moreno, A.; Rovira, J.; Banon-Maneus, E.; et al. Cellular and Humoral Response after MRNA-1273 SARS-CoV-2 Vaccine in Kidney Transplant Recipients. Am. J. Transpl. 2021, 21, 2727–2739. [Google Scholar] [CrossRef] [PubMed]
- Massa, F.; Cremoni, M.; Gérard, A.; Grabsi, H.; Rogier, L.; Blois, M.; Couzin, C.; Ben Hassen, N.; Rouleau, M.; Barbosa, S.; et al. Safety and Cross-Variant Immunogenicity of a Three-Dose COVID-19 MRNA Vaccine Regimen in Kidney Transplant Recipients. EBioMedicine 2021, 73, 103679. [Google Scholar] [CrossRef]
- Yang, J.; Lee, K.W.; Baek, J.Y.; Bae, S.; Lee, Y.H.; Kim, H.; Huh, K.; Cho, S.Y.; Kang, C.-I.; Chung, D.R.; et al. Augmented Humoral and Cellular Immunity against Severe Acute Respiratory Syndrome Coronavirus 2 after Breakthrough Infection in Kidney Transplant Recipients Who Received 3 Doses of Coronavirus Disease 2019 Vaccine. Am. J. Transpl. 2023, 23, 565–572. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501. [Google Scholar] [CrossRef]
- Kidney Disease: Improving Global Outcomes (KDIGO) Transplant Work Group. Special Issue: KDIGO Clinical Practice Guideline for the Care of Kidney Transplant Recipients. Am. J. Transpl. 2009, 9, S1–S155. [Google Scholar] [CrossRef]
- Neerukonda, S.N.; Vassell, R.; Herrup, R.; Liu, S.; Wang, T.; Takeda, K.; Yang, Y.; Lin, T.L.; Wang, W.; Weiss, C.D. Establishment of a Well-Characterized SARS-CoV-2 Lentiviral Pseudovirus Neutralization Assay Using 293T Cells with Stable Expression of ACE2 and TMPRSS2. PLoS ONE 2021, 16, e0248348. [Google Scholar] [CrossRef]
- Ni, L.; Ye, F.; Cheng, M.L.; Feng, Y.; Deng, Y.Q.; Zhao, H.; Wei, P.; Ge, J.; Gou, M.; Li, X.; et al. Detection of SARS-CoV-2-Specific Humoral and Cellular Immunity in COVID-19 Convalescent Individuals. Immunity 2020, 52, 971–977. [Google Scholar] [CrossRef] [PubMed]
- D’Apice, L.; Trovato, M.; Gramigna, G.; Colavita, F.; Francalancia, M.; Matusali, G.; Meschi, S.; Lapa, D.; Bettini, A.; Mizzoni, K.; et al. Comparative Analysis of the Neutralizing Activity against SARS-CoV-2 Wuhan-Hu-1 Strain and Variants of Concern: Performance Evaluation of a Pseudovirus-Based Neutralization Assay. Front. Immunol. 2022, 13, 981693. [Google Scholar] [CrossRef] [PubMed]
- Vocca, I.; Canani, R.B.; Camarca, A.; Ruotolo, S.; Nocerino, R.; Radano, G.; Del Mastro, A.; Troncone, R.; Gianfrani, C. Peripheral Blood Immune Response Elicited by Beta-Lactoglobulin in Childhood Cow’s Milk Allergy. Pediatr. Res. 2011, 70, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, K.; Kato, H.; Ohtake, N.; Stanleyraj Jeremiah, S.; Ryo, A. Enhancement of Humoral and Cellular Immunity against SARS-CoV-2 by a Third Dose of BNT162b2 Vaccine in Japanese Healthcare Workers. J. Infect. Dis. 2022, 227, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mateus, J.; Coelho, C.H.; Dan, J.M.; Moderbacher, C.R.; Gálvez, R.I.; Cortes, F.H.; Grifoni, A.; Tarke, A.; Chang, J.; et al. Humoral and Cellular Immune Memory to Four COVID-19 Vaccines. Cell 2022, 185, 2434–2451. [Google Scholar] [CrossRef]
- Petrone, L.; Picchianti-Diamanti, A.; Sebastiani, G.D.; Aiello, A.; Laganà, B.; Cuzzi, G.; Vanini, V.; Gualano, G.; Grifoni, A.; Ferraioli, M.; et al. Humoral and Cellular Responses to Spike of δ SARS-CoV-2 Variant in Vaccinated Patients with Immune-Mediated Inflammatory Diseases. Int. J. Infect. Dis. 2022, 121, 24–30. [Google Scholar] [CrossRef]
- Heldman, M.R.; Limaye, A.P. SARS-CoV-2 Vaccines in Kidney Transplant Recipients: Will They Be Safe and Effective and How Will We Know? J. Am. Soc. Nephrol. 2021, 32, 1021–1024. [Google Scholar] [CrossRef] [PubMed]
- Deepak, P.; Kim, W.; Paley, M.A.; Yang, M.; Carvidi, A.B.; Demissie, E.G.; El-Qunni, A.A.; Haile, A.; Huang, K.; Kinnett, B.; et al. Effect of Immunosuppression on the Immunogenicity of Mrna Vaccines to SARS-Cov-2 a Prospective Cohort Study. Ann. Intern. Med. 2021, 174, 1572–1585. [Google Scholar] [CrossRef]
- Aslam, S.; Adler, E.; Mekeel, K.; Little, S.J. Clinical Effectiveness of COVID-19 Vaccination in Solid Organ Transplant Recipients. Transpl. Infect. Dis. 2021, 23, e13705. [Google Scholar] [CrossRef]
- Werbel, W.A.; Boyarsky, B.J.; Ou, M.T.; Massie, A.B.; Tobian, A.A.R.; Garonzik-Wang, J.M.; Segev, D.L. Safety and Immunogenicity of a Third Dose of Sars-Cov-2 Vaccine in Solid Organ Transplant Recipients: A Case Series. Ann. Intern. Med. 2021, 174, 1330–1332. [Google Scholar] [CrossRef]
- Hall, V.G.; Ferreira, V.H.; Ku, T.; Ierullo, M.; Majchrzak-Kita, B.; Chaparro, C.; Selzner, N.; Schiff, J.; McDonald, M.; Tomlinson, G.; et al. Randomized Trial of a Third Dose of MRNA-1273 Vaccine in Transplant Recipients. N. Engl. J. Med. 2021, 385, 1244–1246. [Google Scholar] [CrossRef] [PubMed]
- Saharia, K.K.; Husson, J.S.; Niederhaus, S.V.; Iraguha, T.; Avila, S.V.; Yoo, Y.J.; Hardy, N.M.; Fan, X.; Omili, D.; Crane, A.; et al. Humoral Immunity against SARS-CoV-2 Variants Including Omicron in Solid Organ Transplant Recipients after Three Doses of a COVID-19 MRNA Vaccine. Clin. Transl. Immunol. 2022, 11, e1391. [Google Scholar] [CrossRef] [PubMed]
- Karaba, A.H.; Johnston, T.S.; Aytenfisu, T.Y.; Akinde, O.; Eby, Y.; Ruff, J.E.; Abedon, A.T.; Alejo, J.L.; Blankson, J.N.; Cox, A.L.; et al. A Fourth Dose of COVID-19 Vaccine Does Not Induce Neutralization of the Omicron Variant Among Solid Organ Transplant Recipients with Suboptimal Vaccine Response. Transplantation 2022, 106, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeier, E.; Rincon-Arevalo, H.; Stefanski, A.L.; Potekhin, A.; Staub-Hohenbleicher, H.; Choi, M.; Bachmann, F.; Prob, V.; Hammett, C.; Schrezenmeier, H.; et al. B and t Cell Responses after a Third Dose of SARS-Cov-2 Vaccine in Kidney Transplant Recipients. J. Am. Soc. Nephrol. 2021, 32, 3027–3033. [Google Scholar] [CrossRef] [PubMed]
- Reinscheid, M.; Luxenburger, H.; Karl, V.; Graeser, A.; Giese, S.; Ciminski, K.; Reeg, D.B.; Oberhardt, V.; Roehlen, N.; Lang-Meli, J.; et al. COVID-19 MRNA Booster Vaccine Induces Transient CD8+ T Effector Cell Responses While Conserving the Memory Pool for Subsequent Reactivation. Nat. Commun. 2022, 13, 4631. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ramos, A.A.; Poindessous, V.; Marchetti-Laurent, C.; Pallet, N.; Loriot, M.A. The Effect of Immunosuppressive Molecules on T-Cell Metabolic Reprogramming. Biochimie 2016, 127, 23–36. [Google Scholar] [CrossRef]
- Kemlin, D.; Gemander, N.; Depickère, S.; Olislagers, V.; Georges, D.; Waegemans, A.; Pannus, P.; Lemy, A.; Goossens, M.E.; Desombere, I.; et al. Humoral and Cellular Immune Correlates of Protection against COVID-19 in Kidney Transplant Recipients. Am. J. Transpl. 2023, 23, 649–658. [Google Scholar] [CrossRef]
- Bonazzetti, C.; Tazza, B.; Gibertoni, D.; Pasquini, Z.; Caroccia, N.; Fanì, F.; Fornaro, G.; Pascale, R. Relationship between Immune Response to Severe Acute Respiratory Syndrome Coronavirus 2 Vaccines and Development of Breakthrough Infection in Solid Organ Transplant Recipients: The CONTRAST Cohort. Clin. Infect. Dis. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Mistry, P.; Barmania, F.; Mellet, J.; Peta, K.; Strydom, A.; Viljoen, I.M.; James, W.; Gordon, S.; Pepper, M.S. SARS-CoV-2 Variants, Vaccines, and Host Immunity. Front. Immunol. 2022, 12, 809244. [Google Scholar] [CrossRef]
- Goldberg, Y.; Mandel, M.; Bar-On, Y.M.; Bodenheimer, O.; Freedman, L.S.; Ash, N.; Alroy-Preis, S.; Huppert, A.; Milo, R. Protection and Waning of Natural and Hybrid Immunity to SARS-CoV-2. N. Engl. J. Med. 2022, 386, 2201–2212. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Age (ys)/ Sex | Volunteer Condition | Age at Transplant | Years after Transplant | Underlying Disease | T2 Serum Creatinine (mg/dL) | T2 Creatinine Clearance (mL/min) | T2 CKD Stage | T3 Serum Creatinine (mg/dL) | T3 Creatinine Clearance (mL/min) | T3 CKD Stage | T2 (Weeks from 2nd Dose) | T3 (Weeks from 3rd Dose) | Weeks between T2 and T3 | Immunosuppressive Treatment |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EC-73 | 44 ys/F | KTR | 28 | 16 | arterial hypertension | 0.87 | 84 | 2-T | 0.86 | 85 | 2-T | 22 | 17 | 2.4 | TAC 8mg/day |
| EC-75 | 73 ys/M | KTR | 58 | 15 | arterial hypertension | 1.39 | 50.5 | 3a-T | 1.34 | 50.6 | 3a-T | 21 | 20 | 2.8 | TAC 4 mg/day; MMF 500 mg/day |
| EC-76 | 30 ys/M | KTR | 24 | 6 | IgA nephritis | 1.54 | 62 | 2-T | 1.76 | 66.5 | 2-T | 26 | 15 | 2.1 | CsA 60 mg/day; MMF 900 mg/day; PDN 5 mg/day |
| EC-77 | 20 ys/F | KTR | 13 | 7 | nephritis | 0.68 | 127 | 1-T | 0.83 | 103 | 1-T | 22 | 13 | 1.8 | TAC 4 mg/day; MMF 500 mg/day |
| EC-94 | 54 ys/M | KTR | 36 | 18 | obstructive renal failure | 2.22 | 34 | 3b-T | 2.32 | 33 | 3b-T | 27 | 7 | 1.0 | SIR 1 mg/day; mPRED 4 mg/day |
| EC-96 | 30 ys/F | KTR | 23 | 7 | ADPKD | 1.66 | 42 | 3b-T | 1.65 | 41 | 3b-T | 29 | 13 | 1.8 | CsA 170 mg/day; MMF 720 mg/day; mPRED 4 mg/day |
| EC-98 | 57 ys/F | KTR | 45 | 12 | arterial hypertension | 1.2 | 53 | 3a-T | 1.06 | 61 | 2-T | 31 | 5 | 0.7 | TAC 1.5 mg/day; MMF 1000 mg/day; PDN 5 mg/day |
| EC-78 | 68 ys/M | HC | - | - | none | - | - | - | - | - | - | 37 | 17 | 2.4 | none |
| EC-82 | 63 ys/M | HC | - | - | none | - | - | - | - | - | - | 32 | 8 | 1.1 | none |
| EC-88 | 43 ys/M | HC | - | - | none | - | - | - | - | - | - | 39 | 17 | 2.4 | none |
| EC-92 | 37 ys/M | HC | - | - | none | - | - | - | - | - | - | 41 | 10 | 1.4 | none |
| EC-101 | 53 ys/F | HC | - | - | none | - | - | - | - | - | - | 44 | 6 | 0.9 | none |
| EC-102 | 33 ys/F | HC | - | - | none | - | - | - | - | - | - | 43 | 6 | 0.8 | none |
| HC-2 | 41 ys/F | HC | - | - | none | - | - | - | - | - | - | 24 | 4 | 0.6 | none |
| HC-10 | 49 ys/F | HC | - | - | none | - | - | - | - | - | - | 25 | 2 | 0.3 | none |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Mastro, A.; Picascia, S.; D’Apice, L.; Trovato, M.; Barba, P.; Di Biase, I.; Di Biase, S.; Laccetti, M.; Belli, A.; Amato, G.; et al. Booster Dose of SARS-CoV-2 mRNA Vaccine in Kidney Transplanted Patients Induces Wuhan-Hu-1 Specific Neutralizing Antibodies and T Cell Activation but Lower Response against Omicron Variant. Viruses 2023, 15, 1132. https://doi.org/10.3390/v15051132
Del Mastro A, Picascia S, D’Apice L, Trovato M, Barba P, Di Biase I, Di Biase S, Laccetti M, Belli A, Amato G, et al. Booster Dose of SARS-CoV-2 mRNA Vaccine in Kidney Transplanted Patients Induces Wuhan-Hu-1 Specific Neutralizing Antibodies and T Cell Activation but Lower Response against Omicron Variant. Viruses. 2023; 15(5):1132. https://doi.org/10.3390/v15051132
Chicago/Turabian StyleDel Mastro, Andrea, Stefania Picascia, Luciana D’Apice, Maria Trovato, Pasquale Barba, Immacolata Di Biase, Sebastiano Di Biase, Marco Laccetti, Antonello Belli, Gerardino Amato, and et al. 2023. "Booster Dose of SARS-CoV-2 mRNA Vaccine in Kidney Transplanted Patients Induces Wuhan-Hu-1 Specific Neutralizing Antibodies and T Cell Activation but Lower Response against Omicron Variant" Viruses 15, no. 5: 1132. https://doi.org/10.3390/v15051132
APA StyleDel Mastro, A., Picascia, S., D’Apice, L., Trovato, M., Barba, P., Di Biase, I., Di Biase, S., Laccetti, M., Belli, A., Amato, G., Di Muro, P., Credendino, O., Picardi, A., De Berardinis, P., Del Pozzo, G., & Gianfrani, C. (2023). Booster Dose of SARS-CoV-2 mRNA Vaccine in Kidney Transplanted Patients Induces Wuhan-Hu-1 Specific Neutralizing Antibodies and T Cell Activation but Lower Response against Omicron Variant. Viruses, 15(5), 1132. https://doi.org/10.3390/v15051132