

Cellular Immune Responses to SARS-CoV-2 in Exposed Seronegative Individuals
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of Study Participants and Sample Collection
2.2. Serological Testing
2.3. SARS-CoV-2 Pseudo-Neutralization ELISA
2.4. Measurement of Interferon-α
2.5. ELISpot Assay
2.6. In Vitro Stimulation with SARS-CoV-2 Peptides
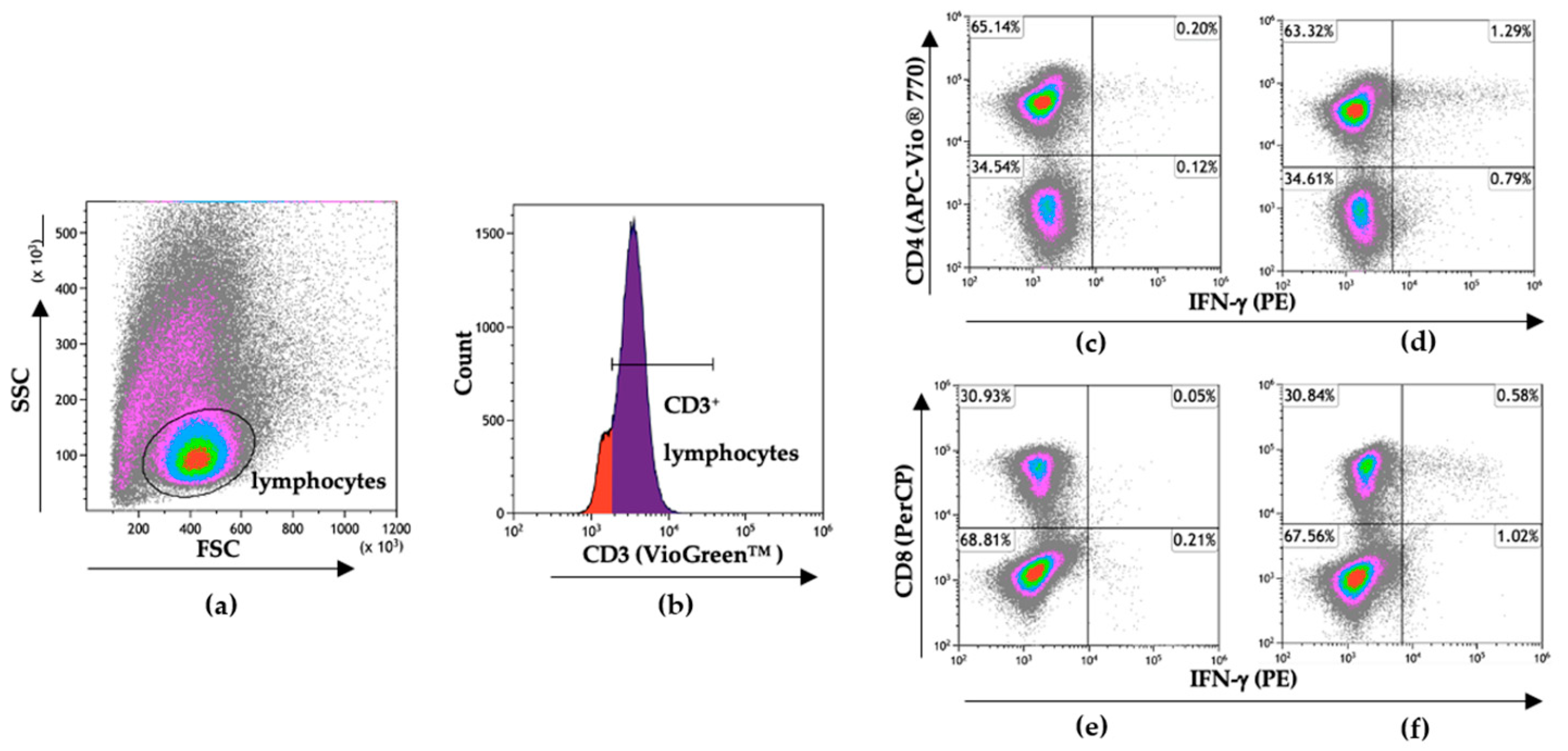
2.7. Flow Cytometry
2.8. Statistical Analysis
3. Results
3.1. Selection of SARS-CoV-2 Exposed Uninfected Persons
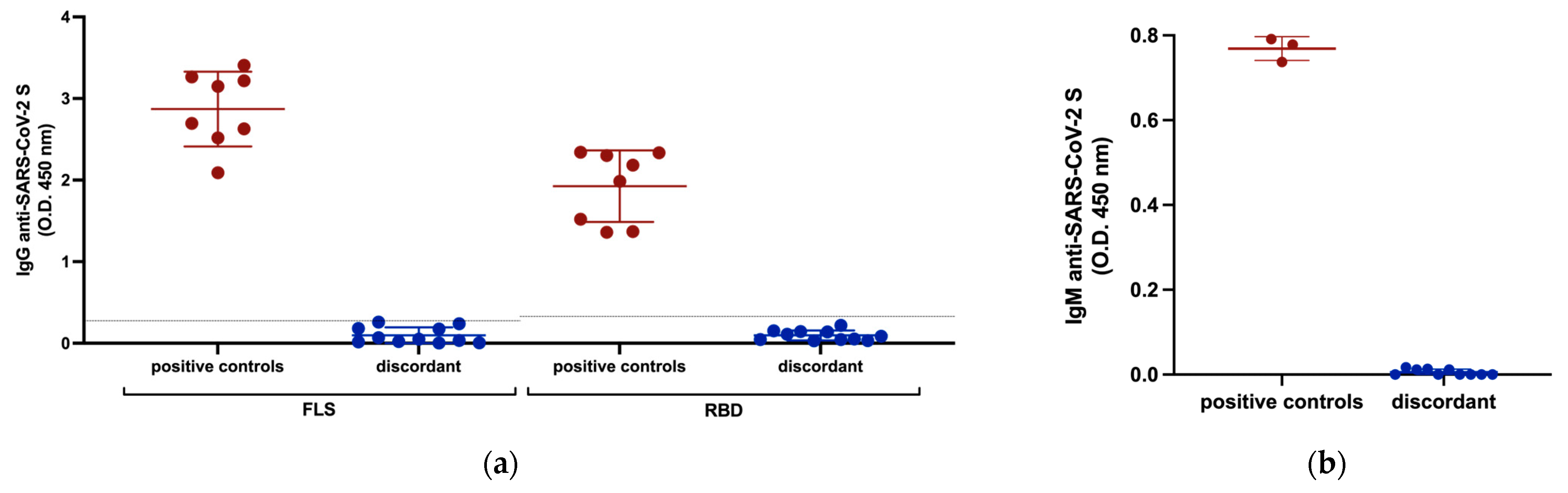
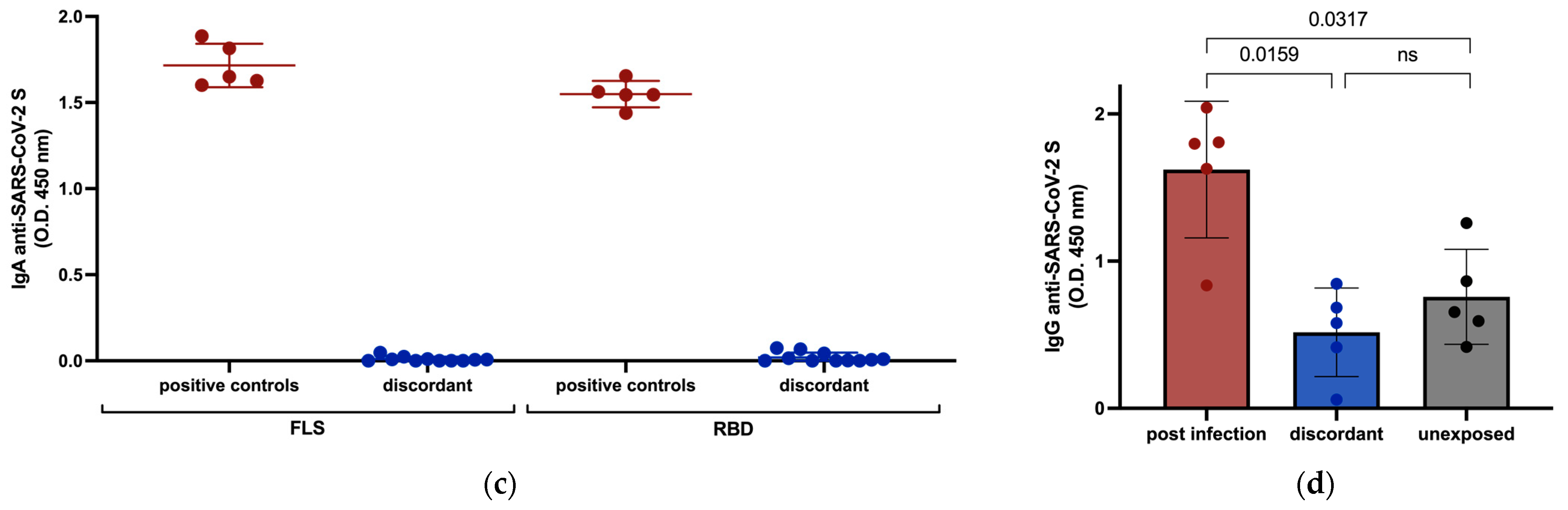
3.2. Anti-SARS-CoV-2 Serology
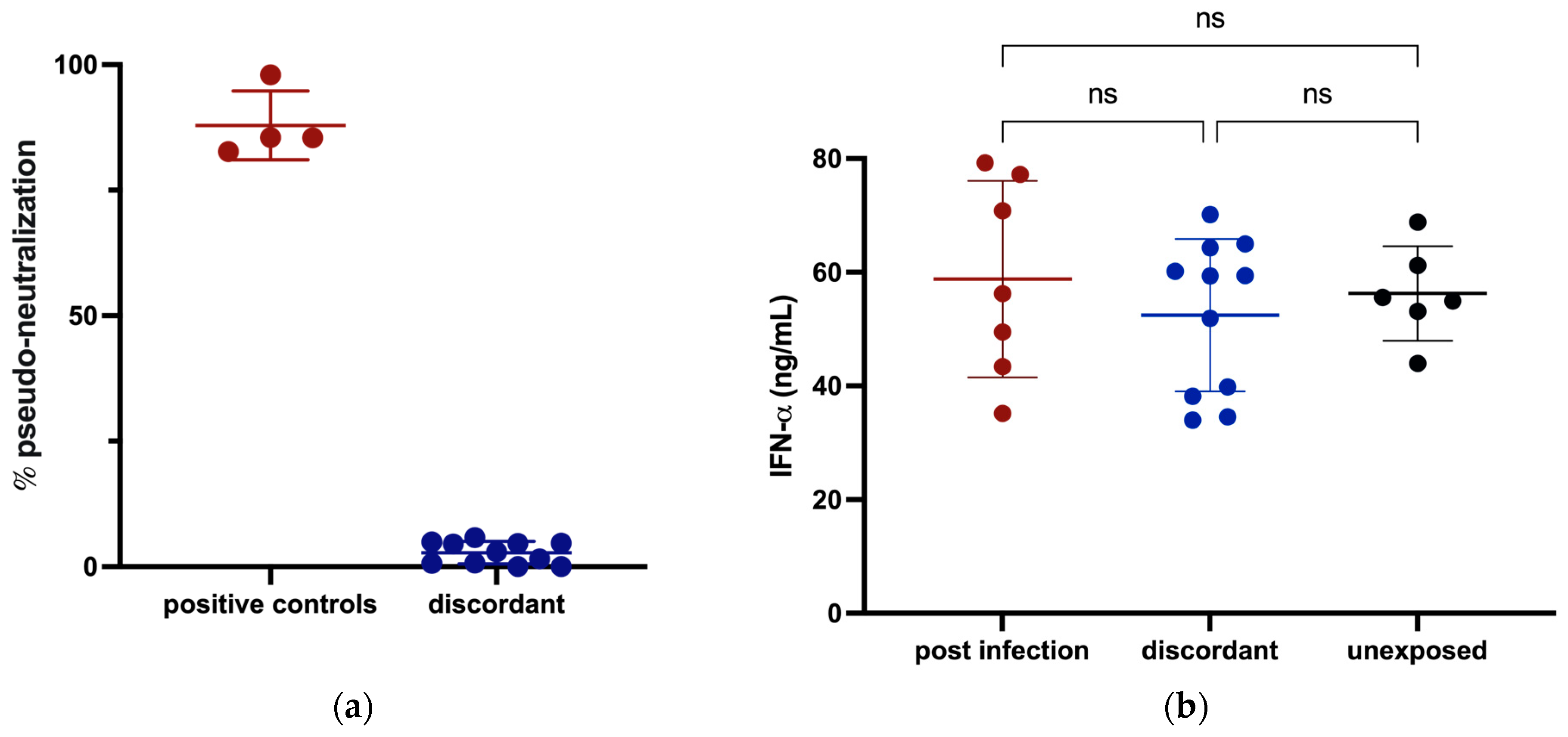
3.3. Innate Immunity against SARS-CoV-2
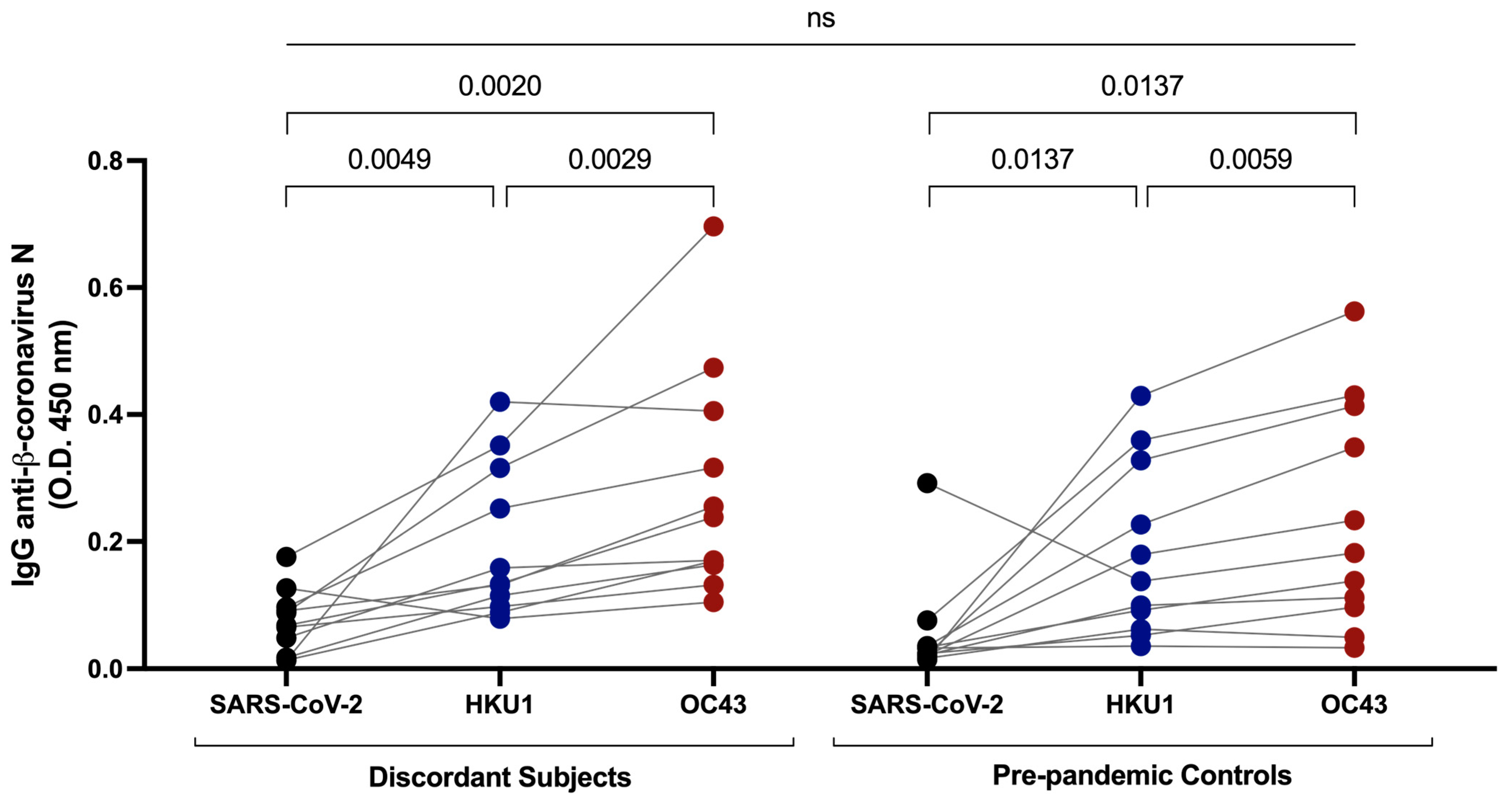
3.4. Cross-Reactive Immunity with Common β-Coronaviruses
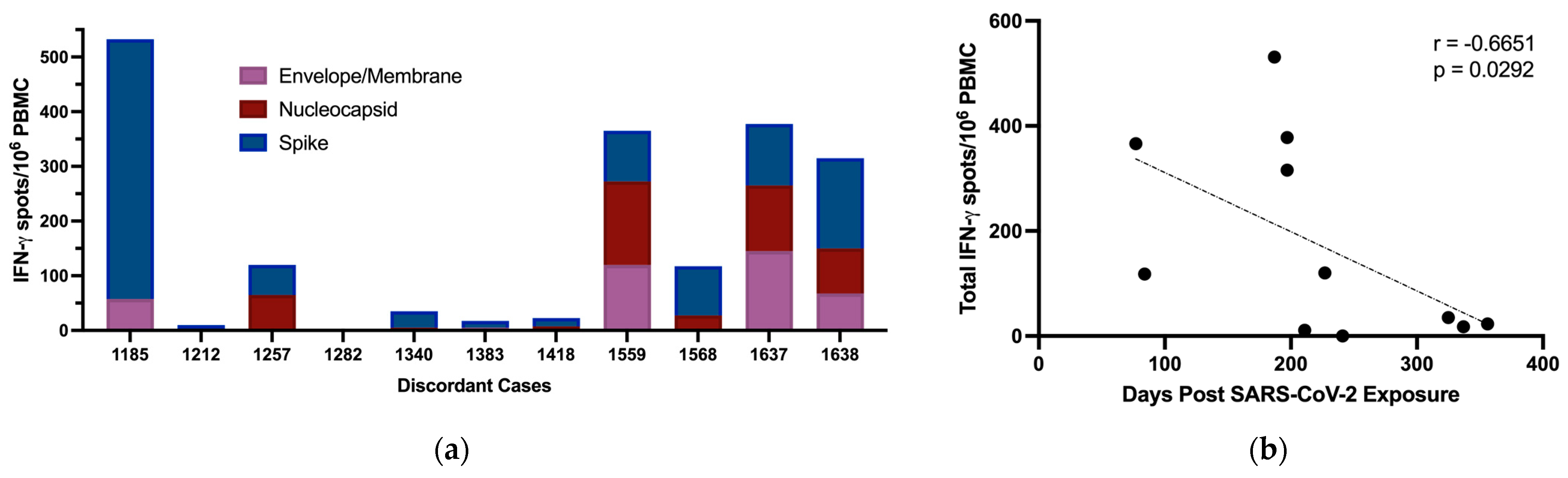
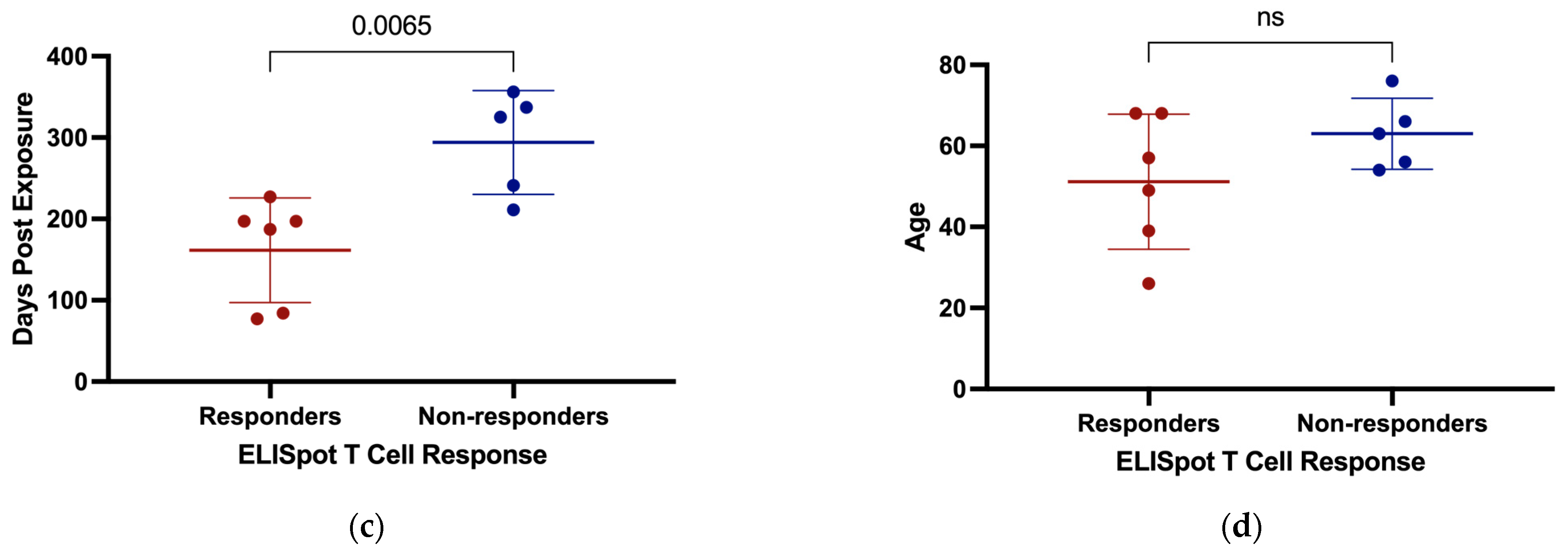
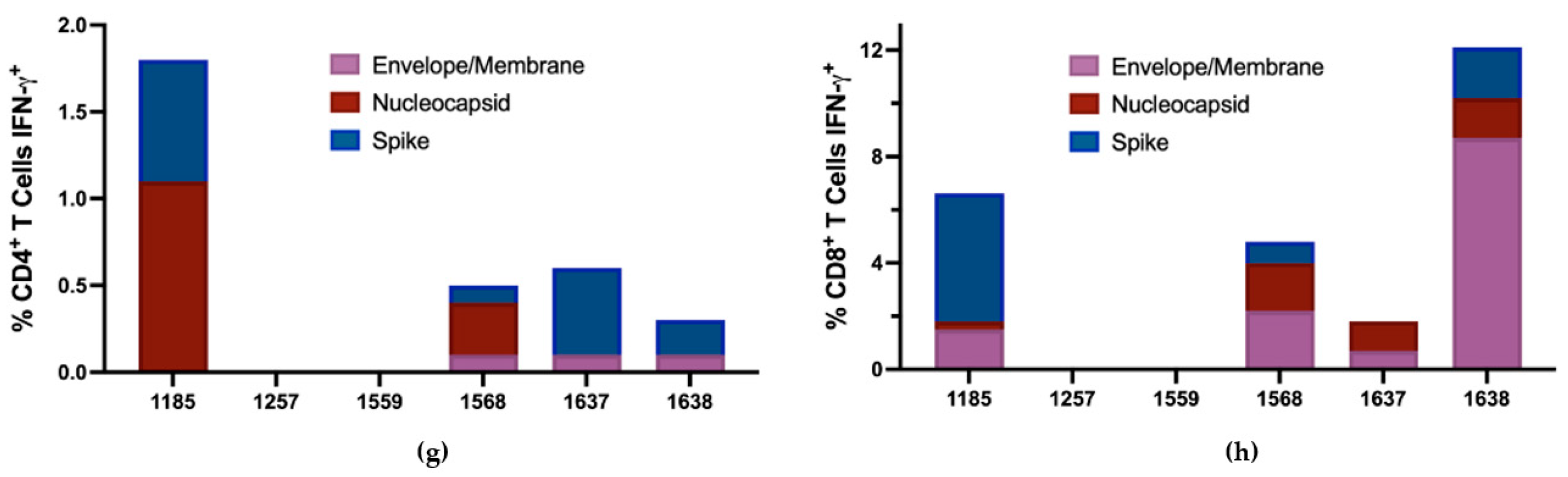
3.5. T Cell Responses to SARS-CoV-2
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swadling, L.; Diniz, M.O.; Schmidt, N.M.; Amin, O.E.; Chandran, A.; Shaw, E.; Pade, C.; Gibbons, J.M.; le Bert, N.; Tan, A.T.; et al. Pre-existing polymerase-specific T cells expand in abortive seronegative SARS-CoV-2. Nature 2021, 601, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Thurairajah, P.H.; Hegazy, D.; Chokshi, S.; Shaw, S.; Demaine, A.; Kaminski, E.R.; Naoumov, N.V.; Cramp, M.E. Hepatitis C virus (HCV)-Specific T cell responses in injection drug users with apparent resistance to HCV infection. J. Infect. Dis. 2008, 198, 1749–1755. [Google Scholar] [CrossRef]
- Heller, T.; Werner, J.M.; Rahman, F.; Mizukoshi, E.; Sobao, Y.; Gordon, A.M.; Sheets, A.; Sherker, A.H.; Kessler, E.; Bean, K.S.; et al. Occupational exposure to hepatitis C virus: Early T-cell responses in the absence of seroconversion in a longitudinal cohort study. J. Infect. Dis. 2013, 208, 1020–1025. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Pinho, M.P.; Buckley, P.R.; Woodhouse, I.B.; Ogg, G.; Simmons, A.; Napolitani, G.; Koohy, H. Potential CD8+ T Cell Cross-Reactivity against SARS-CoV-2 Conferred by Other Coronavirus Strains. Front. Immunol. 2020, 11, 579480. [Google Scholar] [CrossRef]
- Kundu, R.; Narean, J.S.; Wang, L.; Fenn, J.; Pillay, T.; Fernandez, N.D.; Conibear, E.; Koycheva, A.; Davies, M.; Tolosa-Wright, M.; et al. Cross-reactive memory T cells associate with protection against SARS-CoV-2 infection in COVID-19 contacts. Nat. Commun. 2022, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef]
- Anderson, E.M.; Goodwin, E.C.; Verma, A.; Arevalo, C.P.; Bolton, M.J.; Weirick, M.E.; Gouma, S.; McAllister, C.M.; Christensen, S.R.; Weaver, J.E.; et al. Seasonal human coronavirus antibodies are boosted upon SARS-CoV-2 infection but not associated with protection. Cell 2021, 184, 1858–1864.e10. [Google Scholar] [CrossRef]
- Fraley, E.; LeMaster, C.; Banerjee, D.; Khanal, S.; Selvarangan, R.; Bradley, T. Cross-reactive antibody immunity against SARS-CoV-2 in children and adults. Cell. Mol. Immunol. 2021, 18, 1826–1828. [Google Scholar] [CrossRef] [PubMed]
- Dobaño, C.; Santano, R.; Jiménez, A.; Vidal, M.; Chi, J.; Rodrigo Melero, N.; Popovic, M.; López-Aladid, R.; Fernández-Barat, L.; Tortajada, M.; et al. Immunogenicity and crossreactivity of antibodies to the nucleocapsid protein of SARS-CoV-2: Utility and limitations in seroprevalence and immunity studies. Transl. Res. 2021, 232, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Che, X.-Y.; Qiu, L.-W.; Liao, Z.-Y.; Wang, Y.-D.; Wen, K.; Pan, Y.-X.; Hao, W.; Mei, Y.-B.; Cheng, V.C.C.; Yuen, K.-Y. Antigenic Cross-Reactivity between Severe Acute Respiratory Syndrome-Associated Coronavirus and Human Coronaviruses 229E and OC43. J. Infect. Dis. 2005, 191, 2033–2037. [Google Scholar] [CrossRef]
- Sagar, M.; Reifler, K.; Rossi, M.; Miller, N.S.; Sinha, P.; White, L.F.; Mizgerd, J.P. Recent endemic coronavirus infection is associated with less-severe COVID-19. J. Clin. Investig. 2021, 131, e143380. [Google Scholar] [CrossRef] [PubMed]
- Ings, D.P.; Hatfield, K.M.; Fifield, K.E.; Harnum, D.O.A.; Holder, K.A.; Russell, R.S.; Grant, M.D. Few SARS-CoV-2 infections detected in Newfoundland and Labrador in the absence of Public Health Laboratory-based confirmation. PLoS ONE 2022, 17, e0262957. [Google Scholar] [CrossRef]
- Available online: https://www.bcchr.ca/sites/default/files/group-can-asc/CAN-ASC_PBMC%20isolation%2C%20cryopreservation%20and%20thawing_20190321.pdf (accessed on 29 November 2021).
- Holder, K.A.; Ings, D.P.; Harnum, D.O.A.; Russell, R.S.; Grant, M.D. Moderate to severe SARS-CoV-2 infection primes vaccine-induced immunity more effectively than asymptomatic or mild infection. NPJ Vaccines 2022, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Fagyas, M.; Fejes, Z.; Sütő, R.; Nagy, Z.; Székely, B.; Pócsi, M.; Ivády, G.; Bíró, E.; Bekő, G.; Nagy, A.; et al. Circulating ACE2 activity predicts mortality and disease severity in hospitalized COVID-19 patients. Int. J. Infect. Dis. 2022, 115, 8–16. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Zhang, F.; Gan, R.; Zhen, Z.; Hu, X.; Li, X.; Zhou, F.; Liu, Y.; Chen, C.; Xie, S.; Zhang, B.; et al. Adaptive immune responses to SARS-CoV-2 infection in severe versus mild individuals. Signal Transduct. Target. Ther. 2020, 5, 156. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.O.; Cardoso, C.C.; Santos-Pirath, Í.M.; Costa, H.Z.; Gartner, R.; Werle, I.; Mohr, E.T.B.; da Rosa, J.S.; Felisberto, M.; Kretzer, I.F.; et al. The relationship between peripheral immune response and disease severity in SARS-CoV-2-infected subjects: A cross-sectional study. Immunology 2022, 165, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Strålin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef]
- Cameron, B.; Galbraith, S.; Li, H.; Lloyd, A. Correlates and characteristics of hepatitis C virus-specific T-cell immunity in exposed uninfected high-risk prison inmates. J. Viral Hepat. 2013, 20, e96–e106. [Google Scholar] [CrossRef]
- Truong, L.X.; Luong, T.T.; Scott-Algara, D.; Versmisse, P.; David, A.; Perez-Bercoff, D.; Nguyen, N.V.; Tran, H.K.; Cao, C.T.; Fontanet, A.; et al. CD4 cell and CD8 cell-mediated resistance to HIV-1 infection in exposed uninfected intravascular drug users in Vietnam. AIDS 2003, 17, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raya, B.; Kollmann, T.R.; Marchant, A.; MacGillivray, D.M. The immune system of HIV-exposed uninfected infants. Front. Immunol. 2016, 7, 383. [Google Scholar] [CrossRef]
- Fowke, K.R.; Nagelkerke, N.J.D.; Kimani, J.; Anzala, A.O.; Bwayo, J.J.; MacDonald, K.S.; Ngugi, E.N.; Plummer, F.A. Resistance to HIV-1 infection among persistently seronegative prostitutes in Nairobi, Kenya. Lancet 1996, 348, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Shim, C.H.; Cho, S.; Shin, Y.M.; Choi, J.M. Emerging role of bystander T cell activation in autoimmune diseases. BMB Rep. 2022, 55, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Bailey-Bucktrout, S.L.; Jeker, L.T.; Penaranda, C.; Martínez-Llordella, M.; Ashby, M.; Nakayama, M.; Rosenthal, W.; Bluestone, J.A. Instability of the transcription factor Foxp3 leads to the generation of pathogenic memory T cells in vivo. Nat. Immunol. 2009, 10, 1000–1007. [Google Scholar] [CrossRef]
- Kim, T.S.; Shin, E.C. The activation of bystander CD8+ T cells and their roles in viral infection. Exp. Mol. Med. 2019, 51, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Johansson, A.M.; Malhotra, U.; Kim, Y.G.; Gomez, R.; Krist, M.P.; Wald, A.; Koelle, D.M.; Kwok, W.W. Cross-reactive and mono-reactive SARS-CoV-2 CD4+ T cells in prepandemic and COVID-19 convalescent individuals. PLoS Pathog. 2021, 17, e1010203. [Google Scholar] [CrossRef]
- Clerici, M.; Giorgi, J.V.; Chou, C.-C.; Gudeman, V.K.; Zack, J.A.; Gupta, P.; Ho, H.-N.; Nishanian, P.G.; Berzofsky, J.A.; Shearer, G.M. Cell-Mediated Immune Response to Human Immunodeficiency Virus (HIV) Type 1 in Seronegative Homosexual Men with Recent Sexual Exposure to HIV-1. J. Infect. Dis. 1992, 165, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- McChesney, M.B.; Collins, J.R.; Lu, D.; Lu, X.; Torten, J.; Ashley, R.L.; Cloyd, M.W.; Miller, C.J. Occult Systemic Infection and Persistent Simian Immunodeficiency Virus (SIV)-Specific CD4-T-Cell Proliferative Responses in Rhesus Macaques That Were Transiently Viremic after Intravaginal Inoculation of SIV. J. Virol. 1998, 72, 10029–10035. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject ID | Sex | Age | Date of Contact’s Confirmatory Test | Contact’s Symptoms | SARS-CoV-2 Exposure Strain | Days from Exposure to Sample Collection |
|---|---|---|---|---|---|---|
| 1185 | F | 68 | 25 March 2020 | Severe | Wuhan-Hu-1 | 187 |
| 1212 | M | 63 | 30 March 2020 | Moderate | Wuhan-Hu-1 | 211 |
| 1257 | M | 26 | 31 March 2020 to 4 April 2020 1 | Moderate (×1) Mild (×3) 1 | Wuhan-Hu-1 | 227 |
| 1282 | M | 76 | 3 April 2020 | Moderate | Wuhan-Hu-1 | 241 |
| 1340 | F | 56 | 15 March 2020 | Severe | Wuhan-Hu-1 | 325 |
| 1383 | F | 54 | 31 March 2020 | Moderate | Wuhan-Hu-1 | 337 |
| 1418 | F | 66 | 24 March 2020 | Moderate | Wuhan-Hu-1 | 356 |
| 1559 | F | 57 | 10 February 2021 1 | Moderate (×2) 1 | B.1.1.7 | 77 |
| 1568 | M | 49 | 11 February 2021 | Moderate | B.1.1.7 | 84 |
| 1637 | F | 39 | 15 February 2021 2 | Mild 2 | B.1.1.7 | 197 |
| 1638 | M | 68 | B.1.1.7 | 197 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norton, N.J.; Holder, K.A.; Ings, D.P.; Harnum, D.O.A.; Russell, R.S.; Grant, M.D. Cellular Immune Responses to SARS-CoV-2 in Exposed Seronegative Individuals. Viruses 2023, 15, 996. https://doi.org/10.3390/v15040996
Norton NJ, Holder KA, Ings DP, Harnum DOA, Russell RS, Grant MD. Cellular Immune Responses to SARS-CoV-2 in Exposed Seronegative Individuals. Viruses. 2023; 15(4):996. https://doi.org/10.3390/v15040996
Chicago/Turabian StyleNorton, Natasha J., Kayla A. Holder, Danielle P. Ings, Debbie O. A. Harnum, Rodney S. Russell, and Michael D. Grant. 2023. "Cellular Immune Responses to SARS-CoV-2 in Exposed Seronegative Individuals" Viruses 15, no. 4: 996. https://doi.org/10.3390/v15040996
APA StyleNorton, N. J., Holder, K. A., Ings, D. P., Harnum, D. O. A., Russell, R. S., & Grant, M. D. (2023). Cellular Immune Responses to SARS-CoV-2 in Exposed Seronegative Individuals. Viruses, 15(4), 996. https://doi.org/10.3390/v15040996