Abstract
The family Tymoviridae comprises positive-sense RNA viruses, which mainly infect plants. Recently, a few Tymoviridae-like viruses have been found in mosquitoes, which feed on vertebrate sources. We describe a novel Tymoviridae-like virus, putatively named, Guachaca virus (GUAV), isolated from Culex pipiens and Culex quinquefasciatus species of mosquitoes and collected in the rural area of Santa Marta, Colombia. After a cytopathic effect was observed in C6/36 cells, RNA was extracted and processed through the NetoVIR next-generation sequencing protocol, and data were analyzed through the VirMAP pipeline. Molecular and phenotypic characterization of the GUAV was achieved using a 5′/3′ RACE, transmission electron microscopy, amplification in vertebrate cells, and phylogenetic analysis. A cytopathic effect was observed in C6/36 cells three days post-infection. The GUAV genome was successfully assembled, and its polyadenylated 3′ end was corroborated. GUAV shared only 54.9% amino acid identity with its closest relative, Ek Balam virus, and was grouped with the latter and other unclassified insect-associated tymoviruses in a phylogenetic analysis. GUAV is a new member of a family previously described as comprising plant-infecting viruses, which seem to infect and replicate in mosquitoes. The sugar- and blood-feeding behavior of the Culex spp., implies a sustained contact with plants and vertebrates and justifies further studies to unravel the ecological scenario for transmission.
1. Introduction
The family Tymoviridae belongs to the order Tymovirales, a diverse group represented by viruses that infect plants. The family comprises three genera, Tymovirus, Marafivirus, and Maculavirus, which share their virus morphology with non-enveloped virions and icosahedral capsids of coat proteins that are approximately 30 nm in diameter [1]. These viruses are considered members of the alphavirus-like superfamily because of the genome organization, replication, and translation strategies [2]. The identified species contain positive-sense, single-stranded, and 5′capped RNA genomes of approximately 6–7.5 kb encoding at least a large polypeptide that is proteolytically processed by a virus-encoded endopeptidase into proteins with methyltransferase, NTPase/helicase, and RNA-dependent RNA polymerase activities, making up the replicase complex, and depending on the genus, a second protein (coat) translated from a subgenomic RNA at the 3′ region, and other overlapping proteins, such as the movement protein in Tymovirus, P16 and P31 in Maculavirus, and P21 in Marafivirus [1]. While the presence of a tRNA-like structure (TLS) is found in the genus Tymovirus, poly(A)-tailed genomes are characteristic of the genera Maculavirus and Marafivirus (except for Maize rayado fino virus) [3]. Their translation and RNA genome replication occur in the cytoplasm in virus-induced membrane structures at the periphery of chloroplasts and mitochondria [1,4].
Virus transmission in plants occurs through mechanical contact, infected plant material, or insect vectors [1,5]. Insect species from the families Chrysomelidae, Curculionidae, and Cicadellidae may act as vectors of these viruses [1]. While an ecological relationship between plant viruses and plant fluid-feeding insects can be directly inferred, and sustained virus exposure to the cellular environment of the insect vector could impose selection pressure for virus adaptation, there is little evidence of a biological role of insects in actively replicating and amplifying plant viruses at titers for a circulative and propagative transmission [6]. The male mosquito diet (and a complement for females) depends on obtaining sugars and other nutrients from plants [7], which is a plausible explanation for the increasing report of tymoviruses in insects detected through metagenomic studies [8,9,10,11,12,13,14,15,16]. Due to the sugar- and blood-feeding behavior of mosquitoes on plant and vertebrate sources, respectively, the recent isolation of Tymoviridae-like viruses in China, Mexico, and Brazil is of major interest [17,18,19], justifying studies of ecological interactions favoring transmission.
Here, we describe the discovery, using molecular and phenotypic characterization, of a new member of the family Tymoviridae, putatively named the Guachaca virus (GUAV), which was isolated from a mosquito pool of the genus Culex in the Sierra Nevada de Santa Marta (SNSM) and located in the north coast of Colombia, an ecosystem that comprises natural (sylvatic) and intervened (rural and urban) areas.
2. Materials and Methods
2.1. Study Area and Entomological Collection
The viral metagenomic study in hematophagous mosquitoes was carried out in the natural ecosystem of the Sierra Nevada de Santa Marta, Santa Marta, Magdalena, Colombia. The SNSM is an isolated mountain range in northern Colombia, which is separate from the Andes range that runs through the north of the country. Reaching an elevation of 5700 m (18,700 ft) just 42 km (26 mi) from the Caribbean coast, SNSM is the highest coastal range in the tropics, and one of the highest coastal ranges in the world. SNSM encompasses about 17,000 km2 (6600 sq mi) and serves as the source of 36 rivers. The range is in the Departments of Magdalena, Cesar, and La Guajira. A pool containing six specimens identified through classic taxonomy as belonging to the genus Culex (Culex pipiens, Culex quinquefasciatus) was collected on 8 June 2018, using a CO2-baited CDC light trap in a rural setting of Quebrada Valencia (latitude 1,124,300,000 and longitude −7,380,511,111) near the populated center of Puerto Nuevo-Guachaca area. The mosquito pool was transported in liquid nitrogen from the field station to the molecular biology laboratory at the Universidad Cooperativa de Colombia, where it was stored at −80 °C until processing.
2.2. Virus Isolation and Preparation of Viral Stocks
C6/36 cells (CRL-1660, obtained from ATCC), derived from whole Aedes albopictus larvae were cultured in Eagle’s minimal essential medium (MEM), supplemented with 10% and 2% fetal bovine serum (FBS) for growth and maintenance, respectively, and incubated in an atmosphere at 95% relative humidity and 5% CO2 at 28 °C. Vero cells (CCL-81, obtained from ATCC) derived from the kidney of the African green monkey Cercopithecus aethiops were cultured in MEM with 8% and 2% FBS for growing and maintenance, respectively, and incubated in an atmosphere at 95% relative humidity and 5% CO2 at 37 °C.
The mosquito pool was mechanically homogenized for 25 s and 1500 rpm in the BeadBug instrument (Benchmark Scientific Inc., Sayreville, NJ, USA) with the use of ceramic Magna lyser green beads (Roche, Mannheim, Germany) in 1.3 mL of Dulbecco’s phosphate-buffered saline (DPBS) supplemented with 10% of FBS and 1% of penicillin/streptomycin. Subsequently, it was centrifuged at 14,000 rpm for 15 min and filtered with the use of a 0.4 µm syringe filter. A 100 aliquot of the supernatant was used for inoculation on C6/36 and Vero cell cultures growing in 24-well plates, and the adhesion phase was carried out for 1 h, at 28 °C and 37 °C, respectively. Subsequently, 800 µL of MEM supplemented with 2% FBS and antibiotics were added, and the cultures were incubated at the appropriate temperature. Negative (mock infection) control was included in each multi-well plate. Supernatant of the virus isolation was collected on the third day post-infection when cytopathic effect was evidenced. Subsequently, a second passage was carried out in the same conditions. Each supernatant was collected, cleared, aliquoted, and stored at −70 °C.
Virus stocks were obtained from the inoculation of two million C6/36 cells in T25 flasks with a 1/10 dilution of the supernatant of the second passage virus isolation. The inoculation was carried out with 125 µL of the supernatant and 1125 µL of MEM with 2% SFB and 5% tryptose phosphate during one hour at 28 °C. Then, MEM was added to a final volume of 5 mL, and the cell culture was incubated until the cytopathic effect was observed in more than 50% of the cell monolayers around the first three days post-infection. The presence of the virus was confirmed by real-time RT-PCR using custom-made primers as described below.
2.3. RNA Purification
RNA purification from the supernatant of mosquito pool homogenization was performed using the QIAamp Viral RNA mini kit (Qiagen, Hilden, Germany), according to the manufacturer instructions, followed by RNase-free RQ1 DNase treatment to degrade double- and single-stranded DNA (Promega, Dübendorf, Switzerland). The RNA extract was stored at −80 °C.
2.4. Metagenomic Sequencing, Assembly, and Virus Discovery
The RNA extract was used for whole transcriptome amplification through the WTA2 kit (Merck KGaA, Darmstadt, Germany) following the NetoVIR protocol [20]. Subsequently, dsDNA was quantified by fluorometry, and 3 ng was used for library preparation with the Nextera XT DNA library prep kit (Illumina, San Diego, CA, USA). Next-generation sequencing (NGS) was performed in a MiSeq instrument with the MiSeq reagent kit version 3 of 600 cycles (Illumina, San Diego, CA, USA). Resulting sequencing reads were processed with VirMAP, which employs a combination of de novo and reference-guides assemblies and taxonomic classification using GenBank databases “gbvrl” and “gbphage” [21].
2.5. Primer Design for Virus Detection by RT-qPCR
Primer design was carried out for the consensus sequence assembled for the GUAV identified in the present study (Table 1) using the Geneious software v9.1.8 (Biomatters Ltd., San Diego, CA, USA). Real-time RT-PCR was carried out with annealing temperature of 55 °C and the SuperScript™ III Platinum One-Step qRT-PCR kit (Thermo Fisher Scientific, Carlsbad, CA, USA), according to the manufacturer instructions, in the CFX96 Real-Time PCR system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).

Table 1.
Primers and probe used for amplification of the viral helicase gene of the putative Guachaca virus.
2.6. 5′ and 3′ UTR Amplification Using RACE
For 5′ UTR amplification, the 5′/3′ RACE kit second generation (Roche Diagnostics GmbH, Mannheim, Germany) was used. After cDNA synthesis and poly(A) tailing, PCR amplification steps were performed to the poly(A)-tailed cDNA, using the oligo dT-anchor primer and the designed reverse primers, Tymoviridae-like5UTR-R2 and Tymoviridae-like5UTR-R1 (Table 1), according to the manufacturer’s instructions.
To corroborate the presence of the poly(A) tail at the 3′ end of the viral genome previously evidenced by the NGS virus assembly, an assay of cDNA synthesis was carried out with random hexamers or Oligo(dT) primer, followed by PCR amplification with the designed Scheme_20_RIGHT_2 (5′-ttacaaaccaggaacctctgact-3′) and the Oligo(dT) primers. The amplification was performed with final concentration of 250 nM of each primer, 1X PCR buffer, 2 mM MgCl2, 400 µM of each dNTP, 1.25 U of Platinum Taq DNA polymerase (Invitrogen, Carlsbad, CA, USA), and 5 µL of the polyadenylated cDNA in a final reaction volume of 25 µL. The thermal profile was as follows: 95°C for 4 min, 40 cycles (95 °C for 20 s, 55 °C for 30 s, and 72 °C for 30 s), and a final extension at 72 °C for 5 min. The purification of PCR amplicons was performed using the QIAquick PCR purification kit (Qiagen Inc., Chatsworth, CA, USA). DNA sequencing was performed through the Sanger method (Macrogen Inc., Seoul, Republic of Korea). The DNA sequences were edited and assembled into the Geneious software v9.1.8 (Biomatters Ltd., San Diego, CA, USA).
2.7. Complete Genome Annotation
Consensus sequences of the viral genome obtained from NGS and that of the 3′ UTR obtained by the RACE approach and direct sequencing were assembled through Geneious v9.1.8 (Biomatters Ltd., San Diego, CA, USA), and motifs, secondary structures, and antigenic domains were predicted and annotated. InterPro server from ELIXIR infrastructure was used for functional domain prediction [22].
2.8. Evaluation of Virus Growth in Mosquito and Permissiveness of Mammalian Cells
C6/36 cells growing in T25 flasks were inoculated with a 1/10 dilution of the viral stock and incubated under standard conditions, as described above. Supernatants were collected at 3-, 6-, 9-, 12-, 15-, 18-, 24-, 36-, 48-, and 72 h post-infection, and triplicates were evaluated using real-time RT-PCR. Cell cultures were examined every day for the presence and magnitude of cytopathic effect during the assay.
To assess the ability of GUAV to infect human cells, HEK293 (human embryonic kidney), HeLa (adenocarcinoma human cervix epithelial), A549 (adenocarcinoma human alveolar basal epithelial), and U937 (pro monocyte from histiocytic lymphoma) were cultured in T25 flasks at 37 °C with 5% CO2. Each cell line was grown in the following specific medium conditions: HEK293 in DMEM 10% FBS, HeLa and A549 in MEM 8% FBS, and U937 in RPMI 5% FBS and stimulated with PMA at final concentration of 25 µM. Confluent monolayers or suspensions were inoculated with 500 μL of a 1/10 dilution of the viral stock, following BSL-3 practices. The adhesion phase was carried out at 37 °C for 1 h and then each flask was rinsed twice with 5 mL of DPBS, and 5 mL of fresh culture medium supplemented with 2% FBS were added. Cell culture supernatants were collected at 5, 7, 9, 11, and 13 days post-infection, aliquoted, and stored at −70 °C for subsequent RNA purification and real-time RT-PCR.
2.9. Transmission Electron Microscopy (TEM)
C6/36 cells infected with the Tymoviridae-like virus were fixed in 3% glutaraldehyde prepared in phosphate buffer pH 7.2, centrifuged at 3000 rpm for 5 min, resuspended, and washed three times with the phosphate buffer pH 7.2. Subsequently, post-fixation with 1% osmium tetroxide was carried out for one hour, followed by three washes, as described above. The cells were dehydrated by treatment with ethanol solutions in ascending concentrations (50, 70, 80, 95, and 100%) for 15 min each, followed by propylene oxide for 20 min. For the pre-imbibition, the cell pellet was treated with a mixture of propylene oxide and resin Epon-Araldite in a 2:1 ratio followed by another 1:1 mixture, for one hour. To complete the imbibition, pure resin was added, and the cell pellet was left at 4 °C. The next day, the resin was replaced and the was transferred to an oven at 65 °C for 24 h to obtain the polymerized block. In an ultramicrotome, semi-fine sections (0.5 microns) and ultra-fine sections (60 nanometers) were obtained. The sections were contrasted with uranyl acetate and lead citrate for observation in an electron microscope EM109 (Zeiss, Jena, Germany).
2.10. Phylogenetic Analysis
A nucleotide sequence dataset was generated through the identification of the ORF1 in previously released sequences of members of the family Tymoviridae, followed by a multiple sequence alignment (MSA) at the nucleotide and amino acid levels using MAFFT v.7 [23]. Accounting for the high genetic distance between some species of the family, a well-represented and unambiguous region of 3480 nt was selected for phylogenetic analysis. The nucleotide substitution model was estimated for the dataset through ModelFinder [24] and the maximum likelihood method was used for tree reconstruction with IQ-TREE [25]. Branch support was estimated by UltraFast Bootstrap with 1000 replicates [26].
3. Results
3.1. Cytopathic Effect Is Suggestive of Productive Viral Infection in C6/36 Cells
C6/36 cells inoculated with a supernatant of a homogenized mosquito pool of Culex spp. (CIST0019) displayed a cytopathic effect that was characterized by the gradual detachment of the cell monolayer by the third day and expanded to the whole culture by the fifth day post-infection. A second passage of a 1/10 dilution of the cell supernatant in fresh C6/36 cells induced the same effect over the following three days, demonstrating the active replication of a transmissible agent (Figure 1).
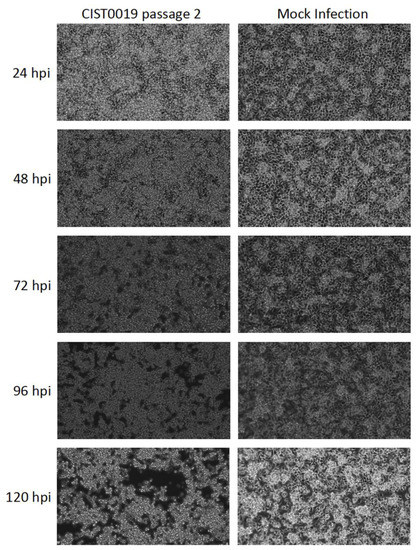
Figure 1.
Cytopathic effect induced in C6/36 cells after infection of the sample CIST0019 passage 2. Morphological changes in the cell culture inoculated with the supernatant of the viral infection with mosquito sample CIST0019, characterized by cell aggregation, were observed from the third day post-infection, and increased during the fourth and fifth day. Mock infection: Uninfected C6/36 cells. Magnification: 200×.
3.2. A Tymoviridae-like Virus Identified through Metagenomic Next-Generation Sequencing
A total of 92,939 paired-end reads were obtained with a Q-score threshold of ≥30. Viral metagenomic analysis using VirMAP [21] enabled the identification of 81,989 viral reads. A total of 81,165 reads (99%) mapped to Ek Balam virus with a genome coverage of 96.2% and a depth of 1824X. Other viral species were identified in the same sample, accounting for an extremely low proportion: Dipteran brevihamaparvovirus (768 reads, 0.9%), Rinkaby virus (43 reads, 0.05%), Culex flavivirus (10 reads, 0.01%), and Atrato Gouko-like virus (3 reads, 0.004%). Low nucleotide and amino acid identities of 57.8% and 54.9%, respectively, to Ek Balam virus were observed, supporting its proposal as a novel species within the family Tymoviridae, putatively named the Guachaca virus (GUAV).
3.3. Putative Guachaca Virus Genome Organization
The full-genome sequence of the putative GUAV was reconstructed from the NGS data plus 3′ UTR corroboration (Figure 2a). The 5′ UTR amplification attempts were unsuccessful; however, the RNA secondary structure of the available sequence at the 5′ end allowed the prediction of a highly stable region (dG = −3819 kcal/mol) with a long and perfectly matched stem-loop structure of 60 nucleotides, similar to those found in other RNA viruses [27], resembling a viral internal ribosomal entry site (IRES) (Figure 2b) [28], and suggesting a selective pressure for a functional role [29]. Extending from the nucleotide position 154 to 5388, a large ORF encoding for a 1744 residues polyprotein was predicted (ORF1). The functional domain’s composition of this ORF was similar to other members of the alphavirus-like superfamily and members of the family Tymoviridae, with the conserved methyltransferase, endopeptidase, helicase, and RNA polymerase sequence motifs resembling a replicase complex (Supplementary Figure S1a). A second ORF (ORF2) was predicted from the nucleotide positions 5418 to 6149, with only 29 nucleotides downstream of the ORF1 stop codon, showing homology to the coat protein of several species belonging to the family Tymoviridae, and displaying a cleavage signal at the amino-terminal region and several antigenic domains distributed throughout the soluble portion of the polypeptide (Supplementary Figure S1b). At the 3′ UTR, a 104 nucleotides sequence was corroborated through RACE-PCR, which was followed by direct sequencing. This sequence was predicted to fold into a partially matched stem-loop structure, followed by a poly(A) tail of approximately 21 nucleotides (Supplementary Figure S2). GUAV genome reconstruction and annotations are available under GenBank accession number: OQ286121.
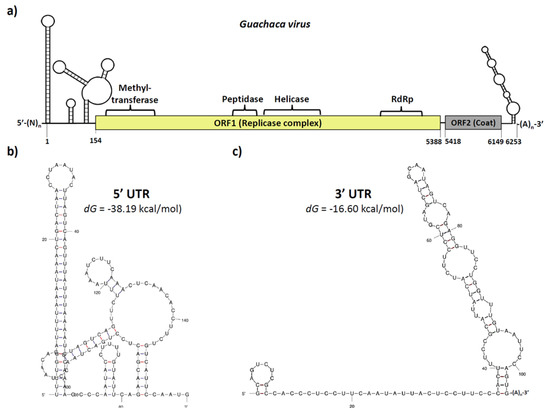
Figure 2.
Genomic organization of the putative Guachaca virus (GUAV). (a) The GUAV genome was approximately 6250 nucleotides in length and comprised a 5′ UTR, a large ORF (ORF1) encoding for the replicase complex, a short ORF (ORF2) encoding for the structural coat protein, a 3′ UTR, and a poly(A) tail. (b) The 5′ UTR of around 153 nucleotides in length contains a highly stable IRES-like structure. (c) The 3′ UTR of around 125 nt in length contains a long partially mismatched stem-loop structure, followed by a poly(A) tail.
3.4. Phylogenetic Inference Supports the Proposal of a New Virus Species and Suggest the Need for New Genera for Insect-Specific Viruses from the Family Tymoviridae
For phylogenetic analysis, an MSA of 2042 amino acids from a conserved region of the ORF1, ranging from amino acid positions 704 to 1723, was performed for 56 sequences, which were representative of the different genera within the family Tymoviridae, and five sequences of species belonging to the family Alphaflexiviridae, which were incorporated as an outgroup. The genera Tymovirus and Marafivirus were well represented, and most species belonging to these genera fell into corresponding monophyletic groups, except for Poinsettia mosaic virus, a species originally assigned the genus Tymovirus, which fell into the genus Marafivirus in our study, and Alfalfa virus F and Medicago sativa marafivirus 1, which were distantly related to the other species within the genus Marafivirus (Figure 3). The genus Maculavirus was represented by only two members of the same species (Grapevine Red Globe virus).
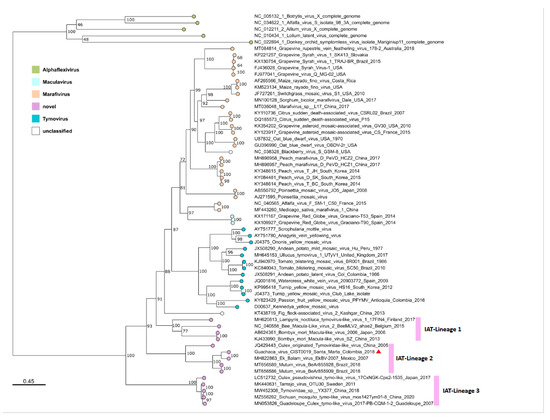
Figure 3.
Phylogeny of the family Tymoviridae. The tree was reconstructed for a partial replicase complex region (2042 amino acids) of 56 representative sequences of the family Tymoviridae and 5 sequences of the family Alphaflexiviridae as an outgroup for rooting. The maximum likelihood method was used with the estimated LG + F + I + G4 protein model and 1000 ultrafast bootstrap replicates. Putative GUAV is denoted by a red triangle. Pink bars denote the three insect-associated Tymoviridae lineages 1 to 3.
It is of special interest that an increasing number of recently described species, and especially those detected or isolated from insects, differ from the previously described genera. This well-supported monophyletic group contains at least three insect-associated Tymoviridae (IAT) lineages; one of them is represented by the proposed virus species infecting Bombyx mori, Andrena haemorrhoa, and Lampyris noctiluca, and the second and third IAT-lineages are represented by viruses infecting mosquitoes of the family Culicidae. The second IAT-lineage with a branch support of 100% comprised several proposed species, including the putative GUAV isolated in 2018 in Colombia from Culex spp. mosquitoes; Ek Balam virus, isolated in 2007 in Mexico from Culex quinquefasciatus; and Mutum virus, isolated in 2018 in Brazil from Mansonia sp. A third IAT-lineage of Culicidae-infecting viruses with a branch support of 100% comprised the unclassified Guadeloupe Culex tymo-like virus, Tarnsjo virus, Tymoviridae sp., Sichuan mosquito tymo-like virus, and Culex pseudovishnui tymo-like virus, detected or isolated from Culex spp., and unclassified mosquitoes from Japan, Guadeloupe, Sweden, and China (Figure 3). The evolutionary divergence estimated from the amino acid differences (p-distances) showed that within-lineage mean distances for the three suggested IAT-lineages were comparable to those obtained for the previously described genera (Maculavirus, Marafivirus, and Tymovirus), consistent with their monophyletic grouping (Table 2). The evolutionary divergence over the sequence pair between the groups showed lower values between the previously described genera (0.456–0.515) than between almost any of these genera and the suggested IAT-lineages (0.520–0.622), except for the Marafivirus-IAT_Lineage_3 (0.505) (Table 2).
Table 2.
Estimates of evolutionary divergence within and between genera and suggested lineages belonging to the family Tymoviridae.
3.5. Ultrastructural Features of Guachaca Virus
The TEM results showed complete icosahedral viral particles with a diameter of 40–50 nm in symmetrical arrangements in the cytoplasm, as well as associated with vesicles that were fused to the cell membrane in the cell periphery of C6/36 cells infected with GUAV 5 days post-infection (Figure 4).
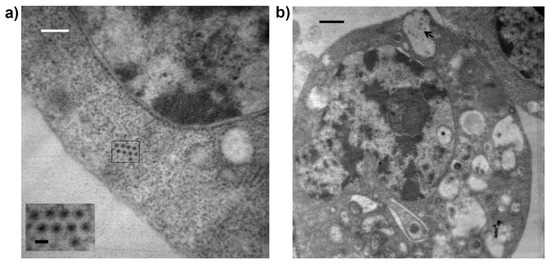
Figure 4.
TEM microphotography of C6/36 cells infected with the putative GUAV. (a) Complete icosahedral viral particles with a diameter of 40–50 nm were observed in the cytoplasm. Black bar: 50 nm. White bar 700 nm. (b) Virus particles observed into vacuoles (arrow). A vacuole was present in the nucleus (asterisk). Black bar 700 nm.
3.6. Vertebrate Cells Were Not Permissible for GUAV Infection
While GUAV successfully replicated in C6/36 cells, showing an increasing titer during the first 72 hpi (Supplementary Figure S2), this virus isolation was unable to replicate in vertebrate cells (Vero, HEK, HELA, A549, and U937). Cell cultures were observed every day without evidence of a cytopathic effect in vertebrate cells and without detection or accumulation of viral RNA by RT-qPCR (Supplementary Figure S3).
4. Discussion
The genus Culex of mosquitoes (Diptera: Culicidae) is of special relevance for public health, with several species serving as vectors for critical human and animal pathogens (e.g., Mayaro virus (MAYV), SLEV, West Nile virus (WNV), etc.). The anthropophilic behavior of this vector species highlights its proximity with humans which, certainly leads to a sustained exposure to the vector virome and the potential transmission of pathogens.
After inoculation of a C6/36 cells culture with a supernatant of Culex spp. that were pool collected in the Sierra Nevada de Santa Marta, an isolated mountain range in the north of Colombia, the cytopathic effect was observed from 3 dpi, gradually extended to the whole monolayer, and was reproduced after a second passage of a diluted inoculum. This finding suggested the presence of a transmissible agent, which was further studied and proposed as a new virus species.
Metagenomic approaches for virus discovery have been refined during the last decade, with the publication of several experimental protocols for virus enrichment and pipelines for taxonomic assignment. In our study, the VirMAP pipeline was successfully applied, allowing the identification of a viral contig that is distantly related to all previously reported viruses, but most closely related to the Ek Balam virus. The metagenomic approach is powerful for the identification of viral signatures, whose genome coverage depends on the viral abundance in a sample. Our results were successful in obtaining the complete genome with high coverage of a new member of the family Tymoviridae, putatively named the Guachaca virus (GUAV). Similar findings have been previously reported in China, Mexico, and Brazil [17,18,19], where genetically related members of the family Tymoviridae were isolated from Culex spp., suggesting the existence of an unclassified group of virus species in this family.
The Inference of the genome organization of GUAV demonstrated that this virus differed from the three previously defined genera, with the absence of a tRNA-like structure and a movement protein characteristic of the genus Tymovirus, and the absence of the accessory proteins present in the genera Maculavirus and Marafivirus.
The RACE strategy for the 5′ end characterization of the GUAV genome was unsuccessful, but the genome assembly from the NGS data enabled the prediction of secondary RNA structures in that region, starting with a long, perfectly matching stem-loop, followed by a structure of major complexity. The RACE strategy applied to the 3′ end enabled the corroboration of the presence of a poly(A) tail. It is of interest that a highly structured 3′ end with a long stem-loop and internal bulges was successfully identified upstream of the poly(A) tail. Tymoviruses have explored several solutions for the 3′ end stability and function, including unstructured polyadenylated and non-polyadenylated tails, tRNA-like structures [3], and the here described structured and polyadenylated end.
While the family Tymoviridae is mainly represented by plant-infecting viruses, the phylogenetic analysis presented here and in some previously reported studies [17,18,19], support the emergence of monophyletic groups conformed by insect-infecting viruses, which could justify the need for new genera proposals and reclassification of the virus species inside the family.
With the design of a molecular approach, the productive infection in C6/36 cells was demonstrated as an increase in viral genome abundance, which was inversely proportional to the Ct values that gradually decreased between the first- and third-day post-infection. This evidence of infection in mosquito cells indirectly suggests that GUAV could be infecting natural populations of Culex mosquitoes, which is also supported by previous studies with closely related viruses [18,19,30].
The TEM findings evidenced the presence of virus-like particles in the cytoplasm of C6/36 cells and particles associated with intracellular membranes, as has been evidenced for other members of the family Tymoviridae in which peripheral vesicles in chloroplasts and/or mitochondria have been reported [1].
All tested mammalian cells were not permissive for GUAV replication. This finding, and the inferred speciation from a family of plant- and insect-associated viruses suggest that the virus circulates in natural cycles, without published evidence of public health implications.
As part of their ecological interactions warranting virus transmission, several insect species have been incriminated as mechanical or biological vectors. Several insect-specific viruses have been shown to dominate the insect viromes [11], considered essential components of ecosystems [31], and ae able to maintain long-term virus-vector interactions. The successful replication of GUAV in mosquito cells and its inferred ability to infect a natural population of mosquitoes supposes an interaction of the virus with other coinfecting insect-specific viruses and arboviruses, leading to unexpected consequences that deserve study. Additional research on Culex spp. vector competence, and virus adaptation to mammalian cells in controlled experimental conditions would help elucidate the molecular and structural basis for receptor binding, host shift, and viral emergence.
5. Conclusions
A novel virus, putatively named the Guachaca virus (GUAV), was discovered and successfully characterized at molecular and phenotypic levels. This virus was unable to replicate in mammalian cells, which suggests its maintenance as an insect-associated virus. While the GUAV biology and transmission cycle remain unraveled, its natural presence in mosquitoes in rural areas, which are in frequent contact with plants and vertebrates as feeding sources, justifies further studies to unravel the ecological scenario for transmission. Phylogenetic evidence warrants a revision of the current taxonomy of the family Tymoviridae to better represent the recently described insect-associated viruses.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/v15040953/s1, Figure S1: Functional domains predicted for the coding regions of the putative Guachaca virus (GUAV). Figure S2: Growth curve of the putative GUAV in C6/36 cells through RT-qPCR. Figure S3: Growth curve of the putative GUAV in human cells through RT-qPCR.
Author Contributions
Conceptualization, K.L.-D. and J.A.U.-C.; methodology, K.L.-D., C.G., L.S., O.T.-F., A.R.-M., G.P.-H. and J.A.U.-C.; software, H.A.R., M.C.W. and N.J.A.; validation, K.L.-D. and J.A.U.-C.; formal analysis, K.L.-D., H.A.R., M.C.W., N.J.A. and J.A.U.-C.; investigation, K.L.-D., E.P.-B., G.P.-H. and J.A.U.-C.; resources, K.L.-D. and J.A.U.-C.; data curation, H.A.R., M.C.W. and N.J.A.; writing—original draft preparation, K.L.-D. and J.A.U.-C.; writing—review and editing, K.L.-D., S.J., M.-C.N., G.P.-H. and J.A.U.-C.; visualization, K.L.-D., H.A.R. and J.A.U.-C.; supervision, J.A.U.-C.; project administration, D.P.-C. and J.A.U.-C.; funding acquisition, K.L.-D. and J.A.U.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Ministry of Science, Technology, and Innovation (Minciencias), Colombia (Grant number: 210477757671 to JAU-C and DP-C).
Institutional Review Board Statement
This study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board (CEMIN, Comité de Ética y Metodologías de Investigación) of the Instituto Nacional de Salud, Colombia (CEMIN-6-2017 from 2 June 2017). Sampling in the urban, rural, and sylvatic areas was framed as a molecular ecology and systematics activity focused on public health and ratified by the Ministry of Environment (Official letter, 8201-2-095, from 11 March 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
The full-length genome of GUAV has been deposited in GenBank under Accession Number OQ286121.
Acknowledgments
The authors would like to thank to the vector-borne disease intervention group at the health secretary of the Santa Marta district, Colombia, for their assistance in the fieldwork and community activities. Additionally, to the families living in the rural areas where mosquitoes were the study was performed.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martelli, G.P.; Sabanadzovic, S.; Abou-Ghanem Sabanadzovic, N.; Edwards, M.C.; Dreher, T. The family Tymoviridae. Arch. Virol. 2002, 147, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, P. Parallels among positive-strand RNA viruses, reverse-transcribing viruses and double-stranded RNA viruses. Nat. Rev. Microbiol. 2006, 4, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Digiaro, M.; Martelli, G.P. Complete sequence of Fig fleck-associated virus, a novel member of the family Tymoviridae. Virus Res. 2011, 161, 198–202. [Google Scholar] [CrossRef]
- Dreher, T.W. Turnip yellow mosaic virus: Transfer RNA mimicry, chloroplasts and a C-rich genome. Mol. Plant Pathol. 2004, 5, 367–375. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Ammar, E.-D.; Whitfield, A.E.; Redinbaugh, M.G. Insect Vector Interactions with Persistently Transmitted Viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [PubMed]
- Dietzgen, R.; Mann, K.; Johnson, K. Plant Virus–Insect Vector Interactions: Current and Potential Future Research Directions. Viruses 2016, 8, 303. [Google Scholar] [CrossRef]
- Foster, W.A. Mosquito Sugar Feeding and Reproductive Energetics. Annu. Rev. Entomol. 1995, 40, 443–474. [Google Scholar] [CrossRef]
- Batson, J.; Dudas, G.; Haas-Stapleton, E.; Kistler, A.L.; Li, L.M.; Logan, P.; Ratnasiri, K.; Retallack, H. Single mosquito metatranscriptomics identifies vectors, emerging pathogens and reservoirs in one assay. Elife 2021, 10, e68353. [Google Scholar] [CrossRef]
- Faizah, A.N.; Kobayashi, D.; Isawa, H.; Amoa-Bosompem, M.; Murota, K.; Higa, Y.; Futami, K.; Shimada, S.; Kim, K.S.; Itokawa, K.; et al. Deciphering the Virome of Culex vishnui Subgroup Mosquitoes, the Major Vectors of Japanese Encephalitis, in Japan. Viruses 2020, 12, 264. [Google Scholar] [CrossRef]
- Frey, K.G.; Biser, T.; Hamilton, T.; Santos, C.J.; Pimentel, G.; Mokashi, V.P.; Bishop-Lilly, K.A. Bioinformatic Characterization of Mosquito Viromes within the Eastern United States and Puerto Rico: Discovery of Novel Viruses. Evol. Bioinform. Online 2016, 12, 1–12. [Google Scholar] [CrossRef]
- Öhlund, P.; Hayer, J.; Lundén, H.; Hesson, J.C.; Blomström, A.-L. Viromics Reveal a Number of Novel RNA Viruses in Swedish Mosquitoes. Viruses 2019, 11, 1027. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, J.H.O.; Shi, M.; Eden, J.S.; Holmes, E.C.; Hesson, J.C. Meta-Transcriptomic Comparison of the RNA Viromes of the Mosquito Vectors Culex pipiens and Culex torrentium in Northern Europe. Viruses 2019, 11, 1033. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Altan, E.; Deng, X.; Barker, C.M.; Fang, Y.; Coffey, L.L.; Delwart, E. Virome of >12 thousand Culex mosquitoes from throughout California. Virology 2018, 523, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Neville, P.; Nicholson, J.; Eden, J.-S.; Imrie, A.; Holmes, E.C. High-Resolution Metatranscriptomics Reveals the Ecological Dynamics of Mosquito-Associated RNA Viruses in Western Australia. J. Virol. 2017, 91, e00680-17. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Wang, Y.; Shi, C.; Atoni, E.; Zhao, L.; Yuan, Z. Comparative Metagenomic Profiling of Viromes Associated with Four Common Mosquito Species in China. Virol. Sin. 2018, 33, 59–66. [Google Scholar] [CrossRef]
- Xiao, P.; Li, C.; Zhang, Y.; Han, J.; Guo, X.; Xie, L.; Tian, M.; Li, Y.; Wang, M.; Liu, H.; et al. Metagenomic Sequencing from Mosquitoes in China Reveals a Variety of Insect and Human Viruses. Front. Cell. Infect. Microbiol. 2018, 8, 364. [Google Scholar] [CrossRef]
- Charles, J.; Tangudu, C.S.; Hurt, S.L.; Tumescheit, C.; Firth, A.E.; Garcia-Rejon, J.E.; Machain-Williams, C.; Blitvich, B.J. Discovery of a novel Tymoviridae-like virus in mosquitoes from Mexico. Arch. Virol. 2019, 164, 649–652. [Google Scholar] [CrossRef]
- Miranda, K.K.P.; Galvão, G.J.P.; da Silva Araújo, P.A.; da Silva Ribeiro, A.C.; da Silva, S.P.; da Silva Lemos, P.; Martins, L.C.; Nunes, M.R.T.; da Costa Vasconcelos, P.F.; da Costa Ferreira, V.; et al. Discovery and genome sequencing of a new virus related to members of the family Tymoviridae, isolated from mosquitoes of the genus Mansonia in Brazil. Arch. Virol. 2022, 167, 1889–1892. [Google Scholar] [CrossRef]
- Wang, L.; Lv, X.; Zhai, Y.; Fu, S.; Wang, D.; Rayner, S.; Tang, Q.; Liang, G. Genomic Characterization of a Novel Virus of the Family Tymoviridae Isolated from Mosquitoes. PLoS ONE 2012, 7, e39845. [Google Scholar] [CrossRef]
- Conceição-Neto, N.; Yinda, K.C.; Van Ranst, M.; Matthijnssens, J. NetoVIR: Modular approach to customize sample preparation procedures for viral Metagenomics. Methods Mol. Biol. 2018, 1838, 85–95. [Google Scholar] [CrossRef]
- Ajami, N.J.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Petrosino, J.F. Maximal viral information recovery from sequence data using VirMAP. Nat. Commun. 2018, 9, 3205. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Thurner, C.; Witwer, C.; Hofacker, I.L.; Stadler, P.F. Conserved RNA secondary structures in Flaviviridae genomes. J. Gen. Virol. 2004, 85, 1113–1124. [Google Scholar] [CrossRef]
- Martinez-Salas, E.; Francisco-Velilla, R.; Fernandez-Chamorro, J.; Embarek, A.M. Insights into Structural and Mechanistic Features of Viral IRES Elements. Front. Microbiol. 2018, 8, 2629. [Google Scholar] [CrossRef]
- Fernández-Sanlés, A.; Ríos-Marco, P.; Romero-López, C.; Berzal-Herranz, A. Functional Information Stored in the Conserved Structural RNA Domains of Flavivirus Genomes. Front. Microbiol. 2017, 8, 546. [Google Scholar] [CrossRef]
- Charles, J.; Tangudu, C.S.; Hurt, S.L.; Tumescheit, C.; Firth, A.E.; Garcia-Rejon, J.E.; Machain-Williams, C.; Blitvich, B.J. Detection of novel and recognized RNA viruses in mosquitoes from the Yucatan Peninsula of Mexico using metagenomics and characterization of their in vitro host ranges. J. Gen. Virol. 2018, 99, 1729–1738. [Google Scholar] [CrossRef]
- French, R.K.; Holmes, E.C. An Ecosystems Perspective on Virus Evolution and Emergence. Trends Microbiol. 2020, 28, 165–175. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).