
Isolation of Flaviviruses and Alphaviruses with Encephalitogenic Potential Diagnosed by Evandro Chagas Institute (Pará, Brazil) in the Period of 1954–2022: Six Decades of Discoveries

 ,
,  , , , ,
, , , ,  , , ,
, , ,  ,
,  , and add
Show full author list
, and add
Show full author list
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Alphavirus Genus
1.1.1. Mayaro Virus (MAYV)
1.1.2. Eastern Equine Encephalitis Virus (EEEV)
1.1.3. Western Equine Encephalitis Virus (WEEV)
1.1.4. Chikungunya Virus (CHIKV)
1.1.5. Pixuna Virus (PIXV), Mucambo Virus (MUCV), Una Virus (UNAV), Aura Virus (AURAV) and Triniti Virus (TNTV)
1.2. Flavivirus Genus
1.2.1. Dengue Virus (DENV)
1.2.2. Saint Louis Encephalitis Virus (SLEV)
1.2.3. West Nile Virus (WNV)
1.2.4. Cacipacoré Virus (CACV), Ilhéus Virus (ILHV) and Bussuquara Virus (BUSV)
2. Methods
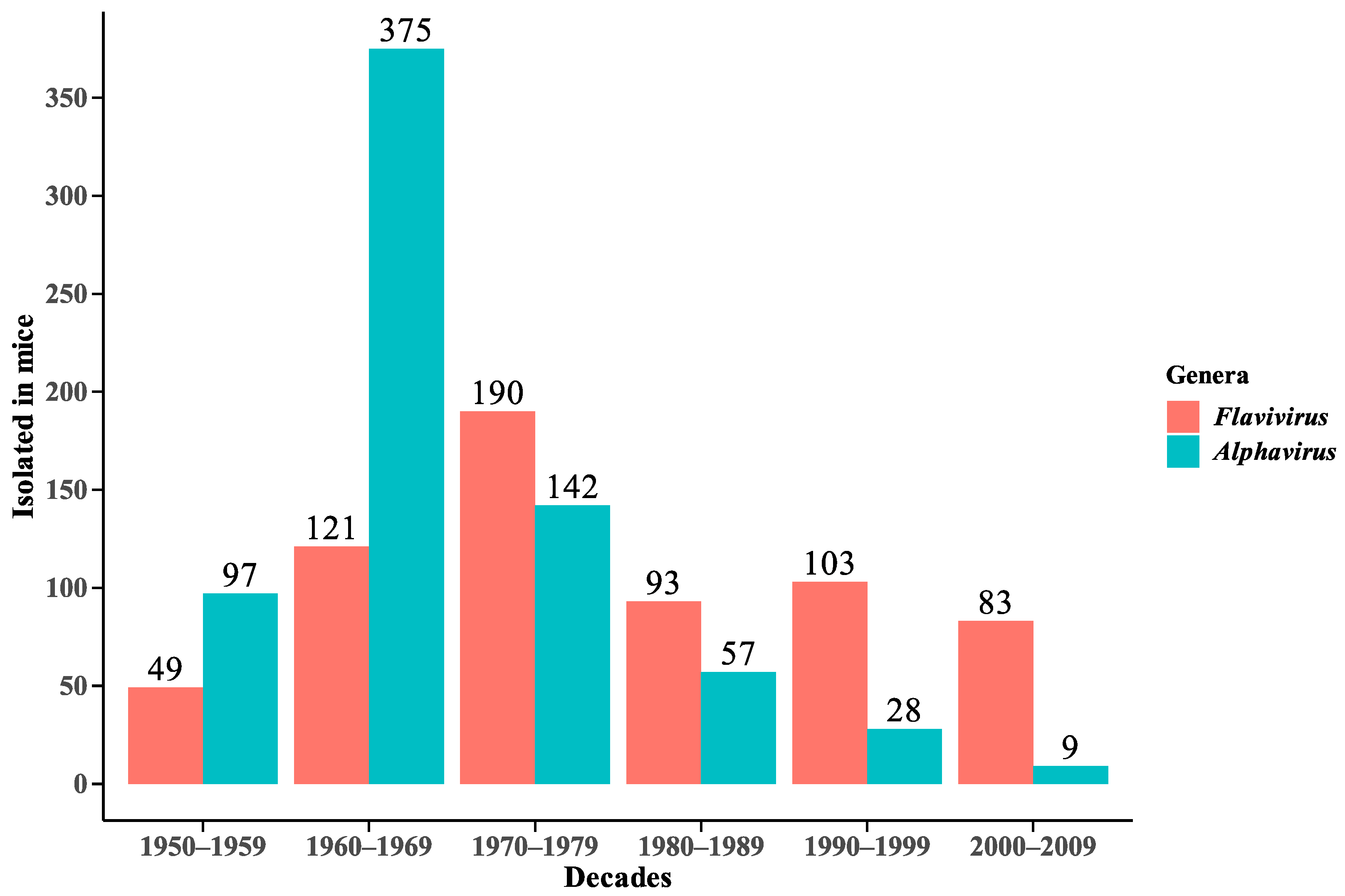
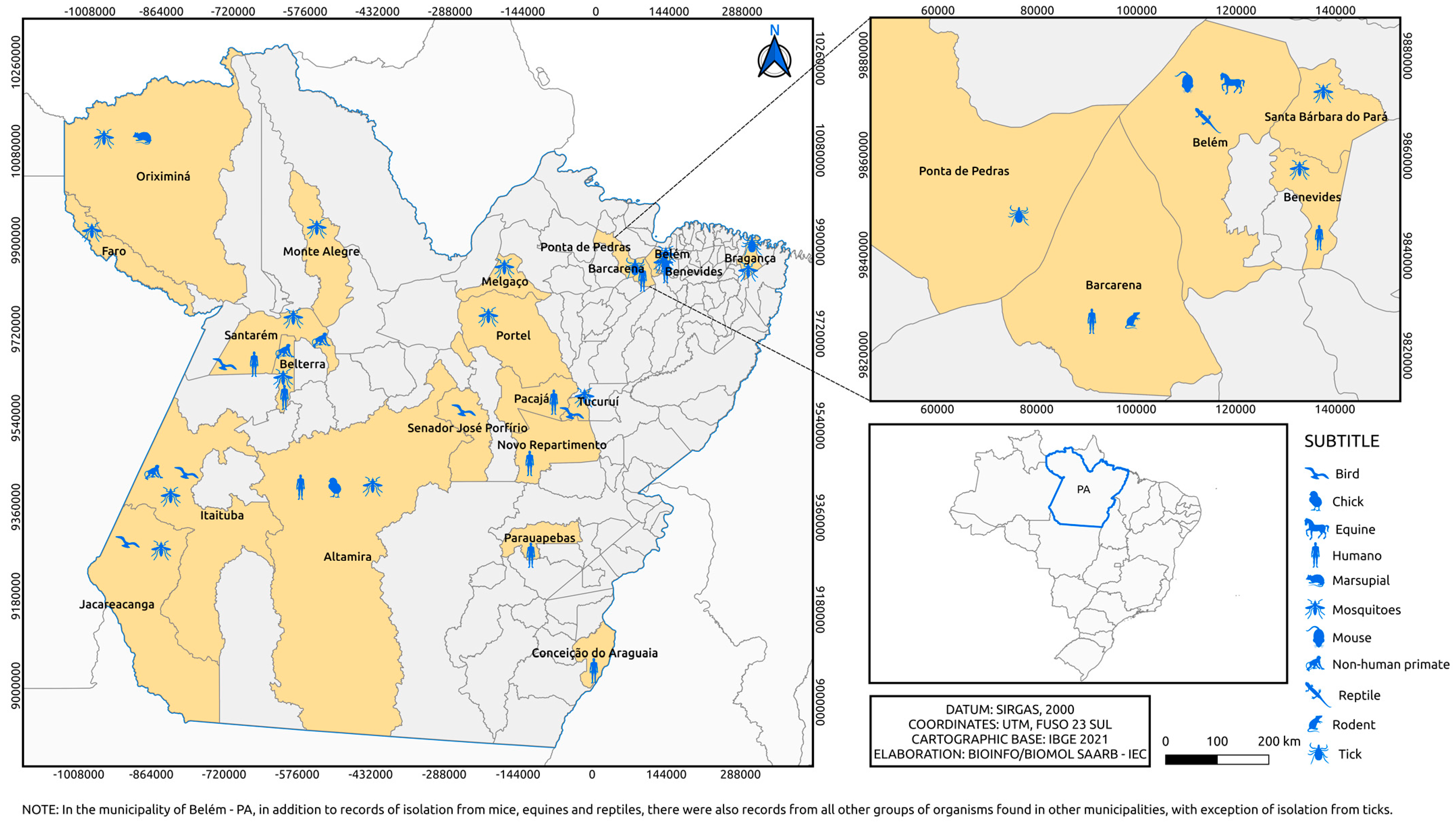
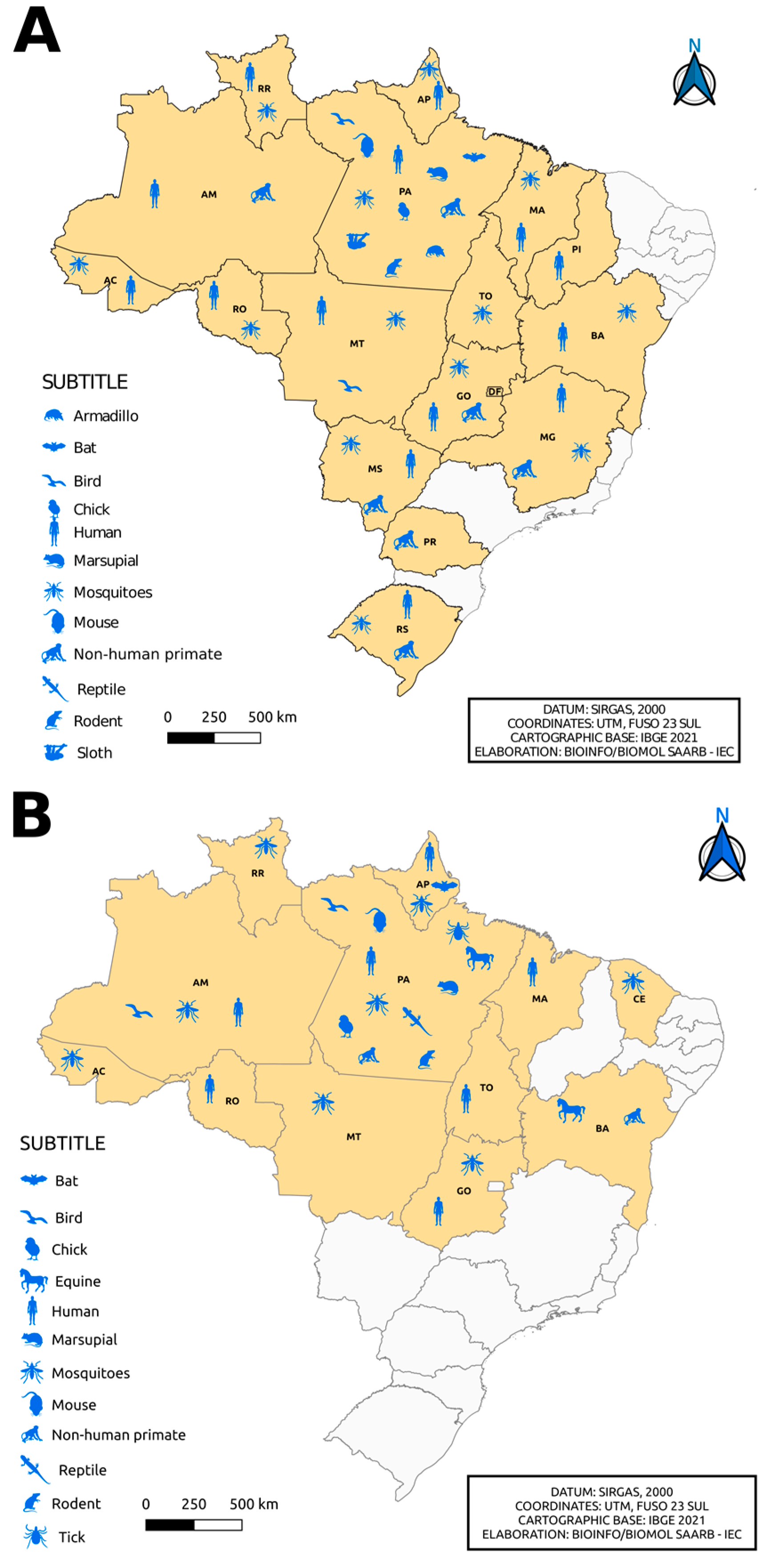
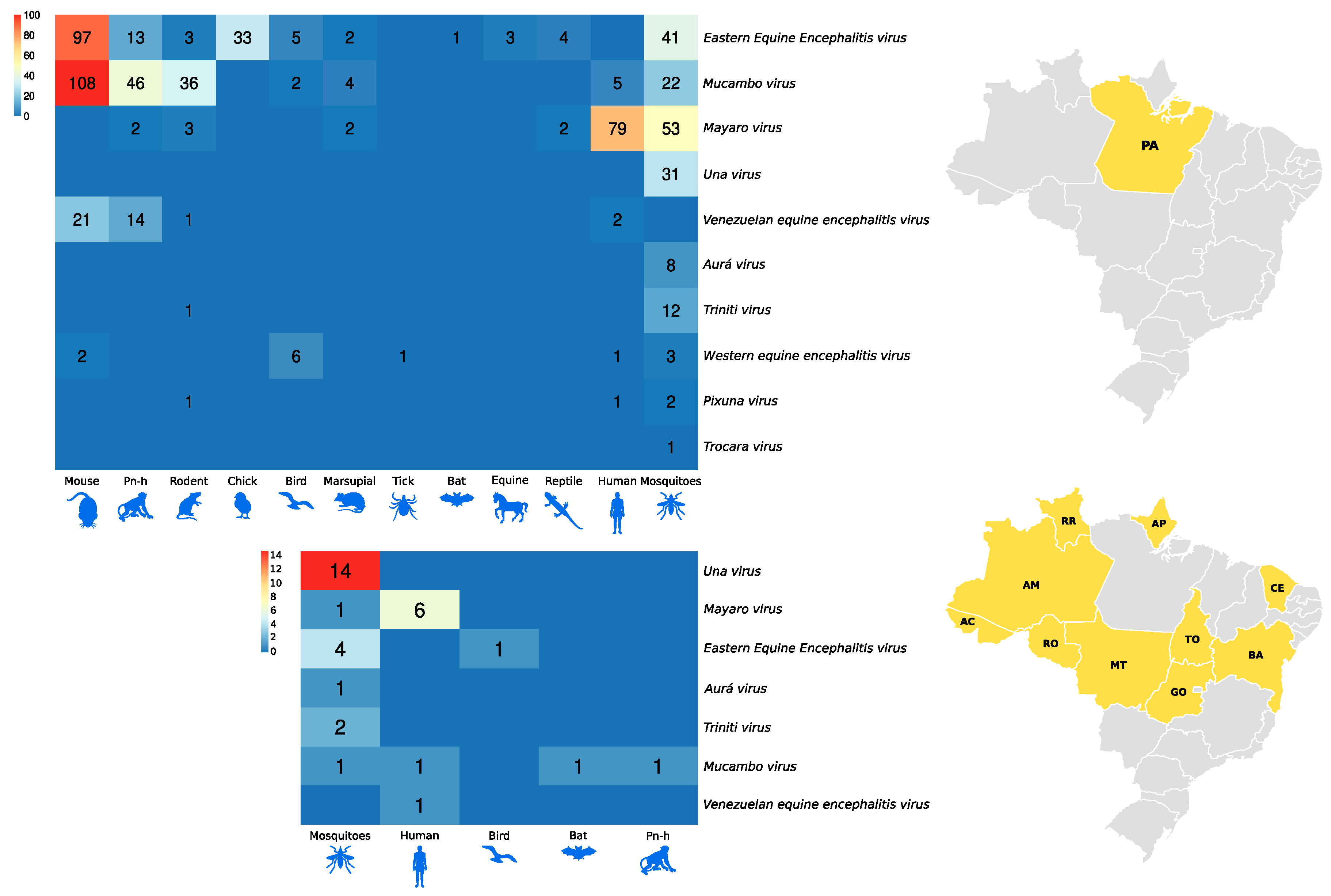
3. Result and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Causey, O.R.; Causey, C.E.; Maroja, O.M.; Macedo, D.G. The isolation of arthropod-borne viruses, including members of two hitherto undescribed serological groups, in the Amazon region of Brazil. Am. J. Trop. Med. Hyg. 1961, 10, 227–249. [Google Scholar] [CrossRef]
- Travassos da Rosa, A.P.A.; Vasconcelos, P.F.C.; Pinheiro, F.P.; Travassos da Rosa, J.F.S.; Rodrigues, S.G. Dengue. In Doenças Infecciosas e Parasitárias: Enfoque Amazônico; Leão, R.N.Q., Ed.; CEJUP: Belém, Brazil, 1997; pp. 227–241. [Google Scholar]
- WHO (World Health Organization). Arthropod-borne and rodent-borne viral diseases. In Technical Report Series, n. 719; World Health Organization: Geneva, Switzerland, 1985. [Google Scholar]
- Brazil Ministry of Health. Manual de Vigilância Sentinela de Doenças Neuroinvasivas por Arbovírus; Ministry of Health: Brasília, Brazil, 2017. [Google Scholar]
- CDC (Centers for Disease Control and Prevention). Arboviral Diseases, Neuroinvasive and Non-Neuroinvasive. Available online: https://ndc.services.cdc.gov/case-definitions/arboviral-diseases-neuroinvasive-and-non-neuroinvasive-2015/ (accessed on 17 March 2022).
- Wilder-Smith, A.; Gubler, D.J.; Weaver, S.C.; Monath, T.P.; Heymann, D.L.; Scott, T.W. Epidemic arboviral diseases: Priorities for research and public health. Lancet Infect. Dis. 2017, 17, e101–e106. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organization). Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Eng. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Lopes, N.; Nozawa, C.; Linhares, R.E.C. Características gerais e epidemiologia dos arbovírus emergentes no Brasil. Rev. Pan-Amaz. Saude 2014, 5, 55–64. [Google Scholar] [CrossRef]
- Donalisio, M.R.; Freitas, R.R.; Von Zuben, A.P.B. Arboviruses emerging in Brazil: Challenges for clinic and implications for public health. Rev. Saude Publica 2017, 51, 30. [Google Scholar] [CrossRef]
- Pimentel, V.; Afonso, R.; Nunes, M.; Vieira, M.L.; Bravo-Barriga, D.; Frontera, E.; Martinez, M.; Pereira, A.; Maia, C.; Paiva-Cardoso, M.N.; et al. Geographic dispersal and genetic diversity of tick-borne phleboviruses (Phenuiviridae, Phlebovirus) as revealed by the analysis of L segment sequences. Ticks Tick Borne Dis. 2019, 10, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Tesh, R.B.; Popov, V.L.; Widen, S.G.; Wood, T.G.; Forrester, N.L.; Gonzalez, J.P.; Saluzzo, J.F.; Alkhovsky, S.; Lam, S.K.; et al. Exploiting the Legacy of the Arbovirus Hunters. Viruses 2019, 11, 471. [Google Scholar] [CrossRef]
- Travassos da Rosa, J.F.S.; Travassos da Rosa, A.P.A.; Vasconcelos, P.F.C.; Pinheiro, F.P.; Rodrigues, S.G.; Travassos da Rosa, E.S.; Dias, L.B.; Cruz, A.C.R. Arboviruses isolated in the Evandro Chagas Institute, including some described for the first time in the Brazilian Amazon region, their know hosts, and their pathology for man. In An Overview of Arbovirology in Brazil and Neighbouring Countries; Travassos da Rosa, J.F.S., Travassos da Rosa, A.P.A., Vasconcelos, P.F.C., Eds.; Instituto Evandro Chagas: Belém, Brazil, 1998; pp. 19–31. [Google Scholar]
- Chen, R.; Mukhopadhyay, S.; Merits, A.; Bolling, B.; Nasar, F.; Coffey, L.L.; Powers, A.; Weaver, S.C.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Togaviridae. J. Gen. Virol. 2018, 99, 761–762. [Google Scholar] [CrossRef]
- Lwande, O.W.; Lutomiah, J.; Obanda, V.; Gakuya, F.; Mutisya, J.; Mulwa, F.; Michuki, G.; Chepkorir, E.; Fischer, A.; Venter, M.; et al. Isolation of tick and mosquito-borne arboviruses from ticks sampled from livestock and wild animal hosts in Ijara District, Kenya. Vector Borne Zoonotic Dis. 2013, 13, 637–642. [Google Scholar] [CrossRef]
- Hayes, R.O.; Francy, D.B.; Lazuick, J.S.; Smith, G.C.; Gibbs, E.P.J. Role of the Cliff Swallow Bug (Oeciacus vicarius) in the Natural Cycle of a Western Equine Encephalitis-Related Alphavirus. J. Med. Entomol. 1977, 14, 257–262. [Google Scholar] [CrossRef]
- Kuhn, R.J. Togaviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 629–650. [Google Scholar]
- Simmond, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef] [PubMed]
- ICTV (International Committee on Taxonomy of Viruses). Currently ICTV Taxonomy Release. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 4 April 2022).
- Pinheiro, F.P.; Freitas, R.B.; Travassos da Rosa, J.F.; Gabbay, Y.B.; Mello, W.A.; LeDuc, J.W. An outbreak of Mayaro virus disease in Belterra, Brazil. I. Clinical and virological findings. Am. J. Trop. Med. Hyg. 1981, 30, 674–681. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Travassos da Rosa, A.P.A.; Rodrigues, S.G.; Rosa, E.S.T.; Dégallier, N.; Travassos da Rosa, J.F. Inadequate management of natural ecosystem in the Brazilian Amazon region results in the emergence and reemergence of arboviruses. Cad. Saude Publica 2001, 17, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, T.L.M.; Santos, C.L.S.; Suzuki, A.; Petrella, S.M.C.; Bisordi, I.; Nagamori, A.H.; Marti, A.T.; Santos, R.N.; Fialho, D.M.; Lavigne, S.; et al. Mayaro virus: Imported cases of human infection in São Paulo State, Brazil. Rev. Int. Med. Trop. 2007, 4, 221–224. [Google Scholar] [CrossRef]
- Azevedo, R.S.; Silva, E.V.; Carvalho, V.L.; Rodrigues, S.G.; Nunes-Neto, J.P.; Monteiro, H.; Peixoto, V.S.; Chiang, J.O.; Nunes, M.R.; Vasconcelos, P.F. Mayaro Fever Virus, Brazilian Amazon. Emerg. Infect. Dis. 2009, 15, 1830–1832. [Google Scholar] [CrossRef]
- Zuchi, N.; Heinen, L.B.; Santos, M.A.; Pereira, F.C.; Slhessarenko, R.D. Molecular detection of Mayaro virus during a dengue outbreak in the state of Mato Grosso, Central-West Brazil. Mem. Inst. Oswaldo Cruz 2014, 109, 820–823. [Google Scholar] [CrossRef]
- Ribeiro, A.P.S.; Chiang, J.O.; Ferreira, M.S.; Henriques, D.F.; Oliveira, C.F.; Carvalho, V.L.; Silva, E.V.P.; Azevedo, R.S.S.; Martins, L.C. Research on the circulation of arboviruses in human populations living in the Municipalities of Parauapebas and Canaã de Carajás, located in the southeastern mesoregion of the state of Pará. Res. Soc. Dev. 2022, 11, e6211326043. [Google Scholar] [CrossRef]
- Pereira, N.T.; Rocha, M.N.; Sucupira, P.H.F.; Carvalho, F.D.; Moreira, L.A. Wolbachia significantly impacts the vector competence of Aedes aegypti for Mayaro virus. Sci. Rep. 2018, 10, 6889. [Google Scholar] [CrossRef] [PubMed]
- Theilacker, C.; Held, J.; Allering, L.; Emmerich, P.; Schmidt-Chanasit, J.; Kern, W.V.; Panning, M. Prolonged polyarthralgia in a German traveller with Mayaro virus infection without inflammatory correlates. BMC Infect. Dis. 2013, 13, 369. [Google Scholar] [CrossRef]
- Neufeld, P.M. About Mayaro Fever: An emergin arbovirus. Rev. Bras Anal. Clin. 2017, 49, 118–119. [Google Scholar] [CrossRef]
- Diagne, C.T.; Bengue, M.; Choumet, V.; Hamel, R.; Pompon, J.; Missé, D. Mayaro Virus Pathogenesis and Transmission Mechanisms. Pathogens 2020, 9, 738. [Google Scholar] [CrossRef]
- Barba, M.; Fairbanks, E.L.; Daly, J.M. Equine viral encephalitis: Prevalence, impact, and management strategies. Vet. Med. 2019, 10, 99–110. [Google Scholar] [CrossRef]
- Beer, J. Doenças Infecciosas em Animais Domésticos; Roca: São Paulo, Brazil, 1999; ISBN 978-8572412483. [Google Scholar]
- Gil, L.H.V.G.; Magalhaes, T.; Santos, B.S.A.S.; Oliveira, L.V.; Oliveira-Filho, E.F.; Cunha, J.L.R.; Fraiha, A.L.S.; Rocha, B.M.M.; Longo, B.C.; Ecco, R.; et al. Active Circulation of Madariaga Virus, a Member of the Eastern Equine Encephalitis Virus Complex, in Northeast Brazil. Pathogens 2021, 10, 983. [Google Scholar] [CrossRef]
- Alice, F.J. Encefalomielite equina na Bahia, estudo de três amostras isoladas. Rev. Bras. Biol. 1951, 11, 125–144. [Google Scholar]
- Cunha, R. Estudos sobre uma amostra de vírus da encefalomielite equina isolada de material proveniente de Recife. Bol. Soc. Bras. Med. Vet. 1954, 14, 201–215. [Google Scholar]
- Causey, O.R.; Shope, R.E.; Laemmert, H.W. Report of an epizootic of encephalomyelitis virus in Pará, Brazil. Rev. Serv. Esp. Saude Publica 1962, 12, 47–50. [Google Scholar]
- Correa, W.M.; Correa, C.N.M. Encefalomielite equina. In Enfermidades Infecciosas dos Mamíferos Domésticos, 2nd ed.; MEDSI: Rio de Janeiro, Brazil, 1992; pp. 635–642. [Google Scholar]
- Iversson, L.B.; Silva, R.A.; Rosa, A.; Barros, V.L.R. Circulation of Eastern Equine Encephalitis, Western equine encephalitis, Ilhéus, Maguari and Tacaiuma viruses in equines of the Brazilian Pantanal, South America. Rev. Inst. Med. Trop. 1993, 35, 355–359. [Google Scholar] [CrossRef]
- Nilsson, M.R.; Sugay, W. Ocorrência da encefalomielite equina em Itaporanga, Estado de São Paulo. Arq. Inst. Biol. 1962, 29, 63–68. [Google Scholar]
- Campos, K.F.; de Oliveira, C.H.S.; Reis, A.B.; Vamasaki, E.M.; Brito, M.F.; Andrade, S.T.J.; Duarte, M.D.; Barbosa, J.D. Surto de encefalomielite equina Leste na Ilha de Marajó, Pará. Pesq. Vet Bras. 2013, 33, 443–448. [Google Scholar] [CrossRef]
- Casseb, A.R.; Casseb, L.M.N.; Silva, S.P.; Vasconcelos, P.F.C. Abovírus: Importante zoonose na Amazônia Brasileira. Vet. Zootec. 2013, 20, 391–403. [Google Scholar]
- Fernández, Z.; Richartz, R.; Travassos da Rosa, A.P.A.; Soccol, V.T. Identification of the encephalitis equine virus, Brazil. Rev. Saude Pública 2000, 34, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Wigg, M.D. Isolamento de uma Amostra de Vírus WEE em Haemagogous Janthinomys. Master’s Dissertation, Rio de Janeiro Federal University, Rio de Janeiro, Brazil, 1977. [Google Scholar]
- Heinemann, M.B.; Souza, M.C.C.; Cortez, A.; Ferreira, F.; Homem, V.S.F.; Ferreira-Neto, J.S.; Soares, R.M.; Cunha, E.M.S.; Richtzenhain, L.J. Soroprevalência da encefalomielite equina do leste e do oeste no Município de Uruará, PA, Brasil. Braz. J. Vet. Res. Anim. Sci. 2006, 43, 137–139. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Travassos da Rosa, J.F.S.; Travassos da Rosa, A.P.A.; Dégallier, N.; Pinheiro, F.D.P. Epidemiologia das encefalites por arbovírus na Amazônia brasileira. Rev. Inst. Med. Trop. S Paulo 1991, 33, 465–476. [Google Scholar] [CrossRef]
- Dexter, E.P.; Dexter, D.D.; Lindsay, C.W.; Ross, R.R.; Lutwick, L. Case of fatal eastern equine encephalitis. ID Cases 2021, 26, e01288. [Google Scholar] [CrossRef]
- Hervé, J.P.; Dégallier, N.; Travassos da Rosa, A.P.A.; Pinheiro, F.P.; Sá Filho, G.C. Arboviroses: Aspectos ecológicos. In Instituto Evandro Chagas: 50 Anos de Contribuição às Ciências Biológicas e à Medicina Tropical; Fundação SESP: Belém, Brazil, 1986; pp. 409–437. [Google Scholar]
- Rust, R.S. Human arboviral encephalitis. Semin Pediatr. Neurol. 2012, 19, 130–151. [Google Scholar] [CrossRef]
- Acha, P.N.; Szyfres, B. Zoonosis y enfermedades transmisibles comunes al hombre y a los animales. In Volumen 1: Bacteriosis y Micosis, 3rd ed.; Pan American Health Organization: Washington, DC, USA, 2003. [Google Scholar]
- Bruno-Lobo, G.; Bruno-Lobo, M.; Travassos, J.; Pinheiro, F.; Pazin, I.P. Estudos sobre arbovírus. III. Isolamento de vírus sorologicamente relacionado ao sub-grupo Western—Sindbis de um caso de encefalomielite equina no Rio de Janeiro. An Microbiol. 1961, 9, 183–195. [Google Scholar]
- Jonkers, A.H.; Downs, W.G.; Spence, L.; Aitken, T.H.G. Arthropod borne encephalitis viruses in Northern South America. II. A serological survey of northeastern Venezuela. Am. J. Trop. Med. Hyg. 1965, 14, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Kenney, J.L.; Deardoff, E.; Wang, E.; Weaver, S.C. Western Equine Encephalitis submergence: Lack of evidence for a decline in virus virulence. Virology 2008, 380, 170–172. [Google Scholar] [CrossRef]
- BVS (Biblioteca Virtual em Saúde). Febre Chikungunya. Available online: https://bvsms.saude.gov.br/febre-de-chikungunya/ (accessed on 17 May 2022).
- Montalbano, C.A.; Bezerra, W.S.P.; Ribeiro, K.M.; Rosa, S.B.A. Doenças Infecciosas de Relevância no Brasil; Atena Editora: Ponta Grossa, Brazil, 2021; p. 340. ISBN 978-655-983-610-9. [Google Scholar]
- Brazilian Ministry of Health. Boletim Epidemiológico v. 54, n. 01. 2023. Available online: https://www.gov.br/saude/pt-br/centrais-de-conteudo/publicacoes/boletins/epidemiologicos/edicoes/2023/boletim-epidemiologico-volume-54-no-01/ (accessed on 7 February 2022).
- Cruz, A.C.R.; Vasconcelos, P.F.C. Arbovírus no Brasil. Biológico 2008, 70, 45–46. [Google Scholar]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A structural perspective of the Flavivirus life cycle. Nat. Rev. Microbiol. 2005, 3, 13–22. [Google Scholar] [CrossRef]
- Coimbra, T.L.; Nassar, E.S.; Nagamori, A.H.; Ferreira, I.B.; Pereira, L.E.; Rocco, I.M.; Ueda-Ito, M.; Romano, N.S. Iguape: A newly recognized flavivirus from São Paulo State, Brazil. Intervirology 1993, 36, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef]
- Catão, R.C. Dengue no Brasil: Abordagem Geográfica na Escala Nacional; Cultura Acadêmica: São Paulo, Brazil, 2012; p. 17. ISBN 978-857-983-332-8. [Google Scholar]
- Bezerra, J.M.T.; Sousa, S.C.; Tauil, P.L.; Carneiro, M.; Barbosa, D.S. Entry of dengue virus serotypes and their geographic distribution in Brazilian federative units: A systematic review. Rev. Bras. Epidemiol. 2021, 24, E210020. [Google Scholar] [CrossRef] [PubMed]
- Oneda, R.M.; Basso, S.R.; Frasson, L.R.; Mottecy, N.M.; Saraiva, L.; Bassani, C. Epidemiological profile of dengue in Brazil between the years 2014 and 2019. Rev. Assoc. Med. Bras. 2021, 67, 731–735. [Google Scholar] [CrossRef]
- Roy, S.K.; Bhattacharjee, S. Dengue virus: Epidemiology, biology, and disease aetiology. Can. J. Microbiol. 2021, 67, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.L.; Amarilla, A.A.; Poloni, T.R.; Covas, D.T.; Aquino, V.H.; Figueiredo, L.T.M. Detection of dengue virus in sera of Brazilian blood donors. Transfusion 2012, 52, 1667–1671. [Google Scholar] [CrossRef]
- Slavov, S.N.; Santos, E.V.; Hespanhol, M.R.; Marta, R.; Rodrigues, E.S.; Haddad, R.; Ubiali, E.M.A.; Covas, D.T.; Kashima, S. Dengue RNA detection and seroprevalence in blood donors during an outbreak in the São Paulo State, Brazil, 2016. J. Med. Virol. 2021, 93, 3344–3349. [Google Scholar] [CrossRef]
- Kubizeski, J.R. Arboviroses Emergentes no Município de Sinop-MT: Pesquisa de Vetores. Bachelor’s Dissertation, Mato Grosso Federal University, Sinop, Brazil, 2017. [Google Scholar]
- Lopes, S.F. Identificação de Flavivirus em aves Silvestres da Amazônia Central. Master’s Dissertation, Amazonas Federal University, Manaus, Brazil, 2011. [Google Scholar]
- Rocco, I.M.; Santos, C.L.; Bisordi, I.; Petrella, S.M.; Pereira, L.E.; Souza, R.P.; Suzuki, A. St. Louis encephalitis vírus: First isolation from a human in São Paulo state, Brasil. Rev. Inst. Med. Trop. S Paulo 2005, 47, 281–285. [Google Scholar] [CrossRef]
- Moraes, M.M.; Kubiszeski, J.R.; Vieira, C.J.S.; Gusmão, A.F.; Pratis, T.S.; Colombo, T.E.; Thies, S.F.; Araujo, F.C.; Zanelli, C.F.; Milhim, B.H.G.A.; et al. Detection of Saint Louis encephalitis virus in two Brazilian states. J. Med. Virol. 2021, 94, 776–781. [Google Scholar] [CrossRef]
- Lavezzo, L.C. Estudo de Arboviruses em Dodores de Sangue na Região Amazônica e em uma Cidade do Interior de São Paulo. Master’s Dissertation, São Paulo State University, São Paulo, Brazil, 2010. [Google Scholar]
- Barbosa, B.C.M. Arbovírus Emergentes no Brasil e Seu Risco de Transmissão por Transfusão Sanguínea: Uma Revisão da Literatura. Master’s Dissertation, Minas Gerais Federal University, Belo Horizonte, Brazil, 2017. [Google Scholar]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A Neurotropic Virus Isolated from the Blood of a Native of Uganda. Am. J. Trop. Med. Hyg. 1940, 20, 471–472. [Google Scholar] [CrossRef]
- Murgue, B.; Zeller, H.; Deubel, V. The ecology and epidemiology of West Nile virus in Africa, Europe and Asia. Curr. Top Microbiol. Immunol. 2002, 267, 195–221. [Google Scholar]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile. Lancet Infect. Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef]
- Melandri, V.; Guimarães, A.E.; Komar, N.; Nogueira, M.L.; Mondini, A.; Fernandez-Sesma, A.; Alencar, J.; Bosch, I. Serological detection of West Nile virus in horses and chicken from Pantanal, Brazil. Mem. Inst. Oswaldo Cruz 2012, 107, 1073–1075. [Google Scholar] [CrossRef]
- Ometto, T.; Durigon, E.L.; de Araujo, J.; Aprelon, R.; de Aguiar, D.M.; Cavalcante, G.T.; Melo, R.M.; Levi, J.E.; de Azevedo Júnior, S.M.; Petry, M.V.; et al. West Nile virus surveillance, Brazil, 2008–2010. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Pauvolid-Corrêa, A.; Campos, Z.; Juliano, R.; Velez, J.; Nogueira, R.M.R.; Komar, N. Serological evidence of widespread circulation of West Nile virus and other flaviviruses in equines of the Pantanal, Brazil. PLoS Neg. Trop. Dis. 2014, 8, e2706. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.C.; Silva, E.V.P.; Casseb, L.M.N.; Silva, S.P.; Cruz, A.C.R.; Pantoja, J.A.S.; Medeiros, D.B.A.; Filho, A.J.M.; Cruz, E.R.M.; Araújo, M.T.F.; et al. First isolation of West Nile virus in Brazil. Mem. Inst. Oswaldo Cruz 2019, 114, e180332. [Google Scholar] [CrossRef]
- Silva, A.S.G.; Rehfeld, I.S.; Santos, B.S.A.S.; Franklin, L.F.S.; Teixeira, R.B.C.; Lobato, Z.I.P.; Guedes, M.I.M.C.; Costa, E.A. Febre do Nilo Ocidental no Brasil: O novo desafio aos medicos-veterinários. Rev. Educ. Cont. Med. Vet Zootec 2021, 19, e38082. [Google Scholar] [CrossRef]
- Costa, E.A.; Bayeux, J.J.M.; Silva, A.S.G.; de Queiroz, G.A.; Santos, B.S.A.S.; Rocha, M.N.; Rehfeld, I.S.; Franklin, L.F.S.; Valle, L.B. Epidemiological surveillance of West Nile virus in the world and Brazil: Relevance of equine surveillance in the context of “One Health”. Braz. J. Vet. Res. Anim. Sci. 2019, 56, e164335. [Google Scholar] [CrossRef]
- Meireles, A.R.; Fernandes, L.F.; Fernande, P.M.G.; da Cruz, G.H.S. Primeiro diagnóstico de febre do Nilo Ocidental em humano em Minas Gerais: Relato de caso. Braz. J. Infect. Dis. 2022, 26, 102294. [Google Scholar] [CrossRef]
- Vieira, M.A.C.S.; Romano, A.P.M.; Borba, A.S.; Silva, E.V.P.; Chiang, J.O.; Eulálio, K.D.; Azevedo, R.S.S.; Rodrigues, S.G.; Almeida-Neto, W.S.; Vasconcelos, P.F.C. West Nile virus encephalitis: The first human case recorded in Brazil. Am. J. Trop. Med. Hyg. 2015, 93, 377–379. [Google Scholar] [CrossRef]
- Colpitts, T.M.; Conway, M.J.; Montgomery, R.R.; Fikrig, E. West Nile Virus: Biology, transmission, and human infection. Clin. Microbiol. Rev. 2012, 25, 635–648. [Google Scholar] [CrossRef]
- Davis, L.E.; Debiasi, R.; Goade, D.E.; Haaland, K.T.; Harrington, J.A.; Harnar, J.B.; Pergam, S.A.; King, M.K.; Demasters, B.K.; Tyler, K.L. West Nile virus neuroinvasive disease. Ann. Neurol. 2006, 60, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Chapman, G.E.; Baylis, M.; Archer, D.; Daly, J.M. The challenges posed by equine arboviruses. Equine Vet. J. 2018, 50, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The Global Ecology and Epidemiology of West Nile Virus. BioMed Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, N.; Phadke, S. Transfusion-Transmitted Diseases. In Transfus Medicine, 5th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2021; pp. 422–452. [Google Scholar]
- Pealer, L.N.; Marfin, A.A.; Petersen, L.R.; Lanciotti, R.S.; Page, P.L.; Stramer, S.L.; Stobierski, M.G.; Signs, K.; Newman, B.; Kapoor, H.; et al. Transmission of West Nile virus through blood transfusion in the United States in 2002. N. Engl. J. Med. 2003, 349, 1236–1245. [Google Scholar] [CrossRef]
- Rios, M.; Sylvester, D.; Chancey, C.; Hewlett, I.K.; Stramer, S.L. West Nile Virus Adheres to Human Red Blood Cells in Whole Blood. Clin. Infect. Dis. 2007, 45, 181–186. [Google Scholar] [CrossRef]
- Harrington, T.; Kuehnert, M.J.; Kamel, H.; Lanciotti, R.S.; Hand, S.; Currier, M.; Chamberland, M.E.; Petersen, L.R.; Marfin, A.A. West Nile virus infection transmitted by blood transfusion. Transfusion 2003, 43, 1018–1022. [Google Scholar] [CrossRef]
- Figueiredo, M.L.G.; Amarilla, A.A.; Figueiredo, G.G.; Alfonso, H.L.; Lippi, V.; Maia, F.G.M.; Morais, F.A.; Costa, C.A.D.; Henriques, D.A.; Durigon, E.L.; et al. Cacipacore virus as an emergent mosquito-borne Flavivirus. Rev. Soc. Bras. Med. Trop. 2017, 50, 539–542. [Google Scholar] [CrossRef]
- Batista, W.C.; Tavares, G.S.B.; Vieira, D.S.; Honda, E.R.; Pereira, S.S.; Tada, M.S. Notification of the first isolation of Cacipacore virus in a human in the State of Rondônia, Brazil. Rev. Soc. Bras. Med. Trop. 2011, 44, 528–530. [Google Scholar] [CrossRef]
- Laemmert, H.W.; Hughes, T.P. The virus of Ilhéus encephalitis: Isolation, serological specificity and transmission. J. Immunol. 1947, 55, 61–67. [Google Scholar] [CrossRef]
- Bernal, M.K.M.; Chiang, J.O.; Mendes, F.F.; Andrade, S.L.S.; da Silva, S.K.S.M.; Pereira, W.L.A. Study of Arboviruses in Philander opossum, Didelphis marsupialis and Nectomys rattus captured from forest fragments in the municipality of Belém, Pará, Brazil. Cienc. Rural 2021, 51, e20200515. [Google Scholar] [CrossRef]
- Pereira, L.E.; Suzuki, A.; Coimbra, T.L.M.; Souza, R.P.; Chamelet, E.L.B. Arbovírus Ilheus em aves silvestres (Sporophila caerulescens e Molothrus bonariensis). Rev. Saude Publica 2001, 35, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Gomes, G.; Causey, O.R. Bussuquara, a new arthropod-borne virus. Proc. Soc. Exp. Biol. Med. 1959, 101, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Srihongse, S.; Johnson, C.M. The first isolation of Bussuquara virus from man. Trans. R. Soc. Trop. Med. Hyg. 1971, 65, 541–542. [Google Scholar] [CrossRef]
- Gibrail, M.M. Detecção de Anticorpos Para Arbovírus em Primatas não Humanos no Município de Goiânia, Goiás. Master’s Dissertation, Goiás Federal University, Goiânia, Brazil, 2015. [Google Scholar]
- Casals, J. The arthropod-borne group of animal viruses. Trans. R. Soc. Trop. Med. Hyg. 1957, 19, 219–235. [Google Scholar]
- Gubler, D.J.; Kuno, G.; Sather, G.E.; Velez, M.; Olivier, A. Mosquito cell cultures and specific monoclonal antibodies in surveillance for dengue viruses. Am. J. Trop. Med. Hyg. 1984, 33, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Meslin, F.X.; Stohr, K.; Heymann, D. Public health implications of emerging zoonoses. Rev. Sci. Tech. 2001, 19, 310–317. [Google Scholar] [CrossRef]
- Davis, G.H.N.G. Estudos Epidemiológicos Sobre Arbovírus em Populações Rurais e Urbanas do Estado do Amazonas. Master’s Dissertation, Amazonas Federal University, Manaus, Brazil, 2009. [Google Scholar]
- IEC (Evandro Chagas Institute). O Estudo da Arbovirologia em Defesa da Amazônia. Available online: https://antigo.iec.gov.br/especial-85-anos-arbovirologia/ (accessed on 4 April 2022).
- Vasconcelos, H.B.; Nunes, M.R.; Casseb, L.M.; Carvalho, V.L.; Pinto da Silva, E.V.; Silva, M.; Casseb, S.M.; Vasconcelos, P.F. Molecular epidemiology of Oropouche virus, Brazil. Emerg. Infect. Dis. 2011, 17, 800–806. [Google Scholar] [CrossRef]
- Chastel, C. Asymptomatic infections in man: A Trojan horse for the introduction and spread of mosquito-borne arboviruses in non-endemic areas? Bull Soc. Pathol. Exot. 2011, 104, 213–219. [Google Scholar] [CrossRef]
- Faria, N.R.; Azevedo, R.S.S.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika virus in the Americas: Early epidemiological and genetic findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef]
- Marbán-Castro, E.; Goncé, A.; Fumadó, V.; Romero-Acevedo, L.; Bardají, A. Zika virus infection in pregnant women and their children: A review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 265, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Lowe, R.; Lee, S.; Martins, L.R.; Torres, C.C.; Castro, M.C.; Pascual, M. Emerging arboviruses in the urbanized Amazon rainforest. BMJ 2020, 371, m4385. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.; McCarthy, K.; Bedford, P.; Johnson, C.T.; Nichols, R.; Yoksan, S.; Marchesani, R.; Knauber, M.; Wells, K.H.; Arroyo, J.; et al. Clinical proof of principle for ChimeriVax: Recombinant live, attenuated vaccines against flavivirus infections. Vaccine 2002, 20, 1004–1018. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanzeller, A.L.M.; da Silva, F.S.; Hernández, L.H.A.; Barros, L.J.L.; Freitas, M.N.O.; Santos, M.M.; Gonçalves, E.d.J.; Pantoja, J.A.S.; Lima, C.d.S.; Lima, M.F.; et al. Isolation of Flaviviruses and Alphaviruses with Encephalitogenic Potential Diagnosed by Evandro Chagas Institute (Pará, Brazil) in the Period of 1954–2022: Six Decades of Discoveries. Viruses 2023, 15, 935. https://doi.org/10.3390/v15040935
Wanzeller ALM, da Silva FS, Hernández LHA, Barros LJL, Freitas MNO, Santos MM, Gonçalves EdJ, Pantoja JAS, Lima CdS, Lima MF, et al. Isolation of Flaviviruses and Alphaviruses with Encephalitogenic Potential Diagnosed by Evandro Chagas Institute (Pará, Brazil) in the Period of 1954–2022: Six Decades of Discoveries. Viruses. 2023; 15(4):935. https://doi.org/10.3390/v15040935
Chicago/Turabian StyleWanzeller, Ana Lucia Monteiro, Fabio Silva da Silva, Leonardo Henrique Almeida Hernández, Landerson Junior Leopoldino Barros, Maria Nazaré Oliveira Freitas, Maissa Maia Santos, Ercília de Jesus Gonçalves, Jamilla Augusta Sousa Pantoja, Creuza de Sousa Lima, Maxwell Furtado Lima, and et al. 2023. "Isolation of Flaviviruses and Alphaviruses with Encephalitogenic Potential Diagnosed by Evandro Chagas Institute (Pará, Brazil) in the Period of 1954–2022: Six Decades of Discoveries" Viruses 15, no. 4: 935. https://doi.org/10.3390/v15040935
APA StyleWanzeller, A. L. M., da Silva, F. S., Hernández, L. H. A., Barros, L. J. L., Freitas, M. N. O., Santos, M. M., Gonçalves, E. d. J., Pantoja, J. A. S., Lima, C. d. S., Lima, M. F., Costa, L. R. O., das Chagas, L. L., Silva, I. F., da Cunha, T. C. A. d. S., do Nascimento, B. L. S., Vasconcelos, H. B., da Rosa, E. S. T., Rodrigues, S. G., Azevedo, R. d. S. d. S., ... da Silva, E. V. P. (2023). Isolation of Flaviviruses and Alphaviruses with Encephalitogenic Potential Diagnosed by Evandro Chagas Institute (Pará, Brazil) in the Period of 1954–2022: Six Decades of Discoveries. Viruses, 15(4), 935. https://doi.org/10.3390/v15040935