
Citrus tristeza virus Promotes the Acquisition and Transmission of ‘Candidatus Liberibacter Asiaticus’ by Diaphorina citri
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Diaphorina citri Samples
2.3. RNA Extraction and cDNA Synthesis
2.4. Quantification of CTV and CLas by RT–qPCR
2.5. Sample Collection and Experimental Designs under Laboratory Conditions
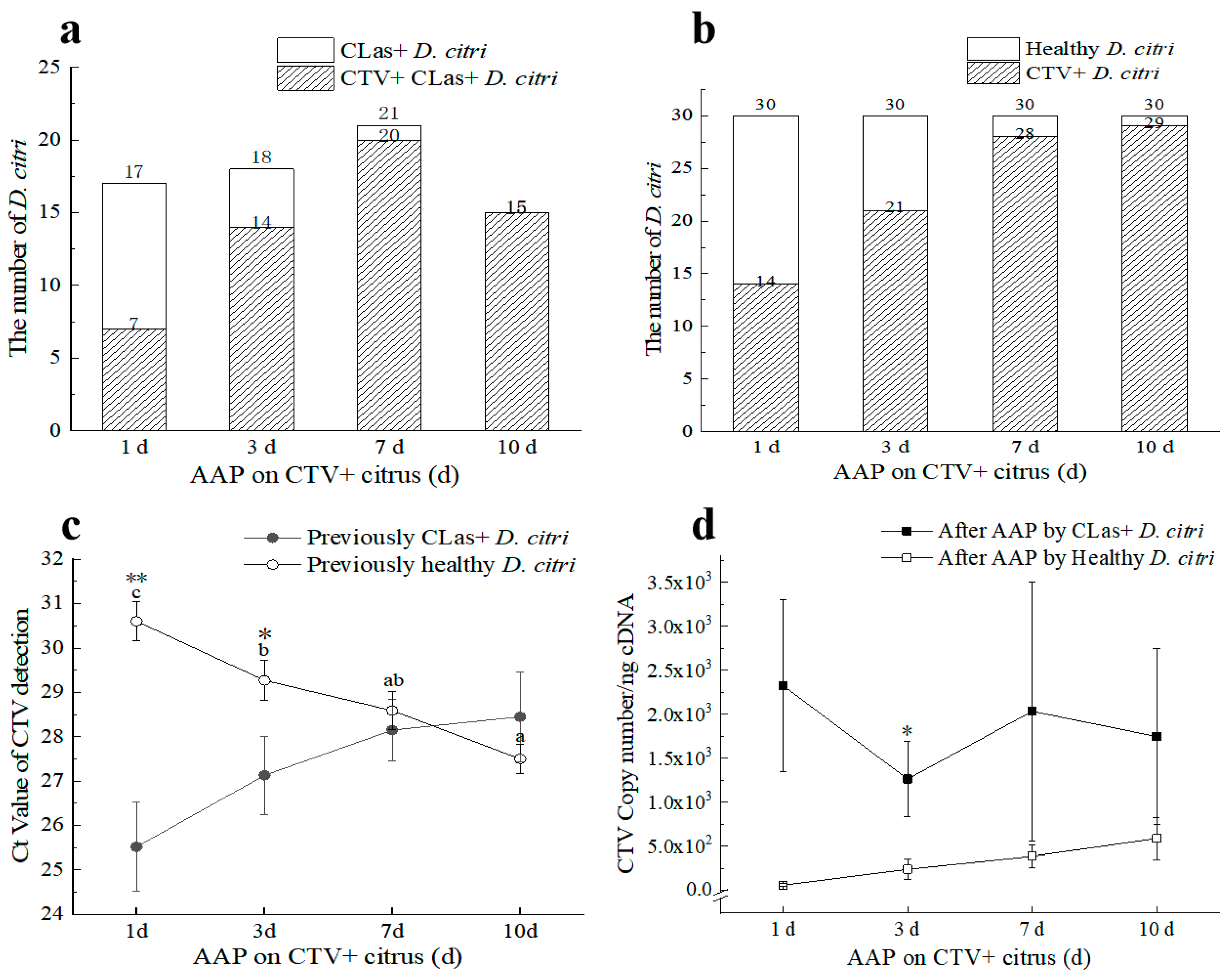
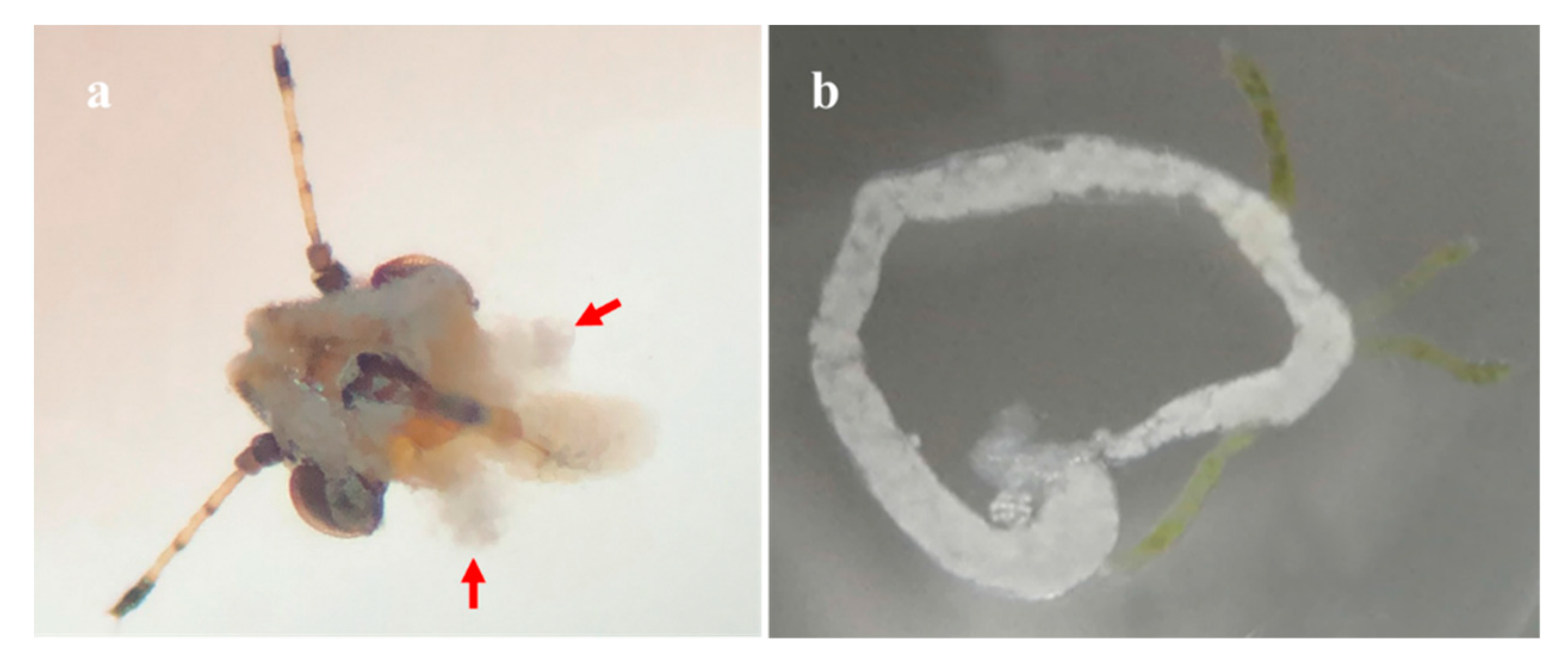
2.6. The Acquisition and Persistence Mode of CTV by D. citri
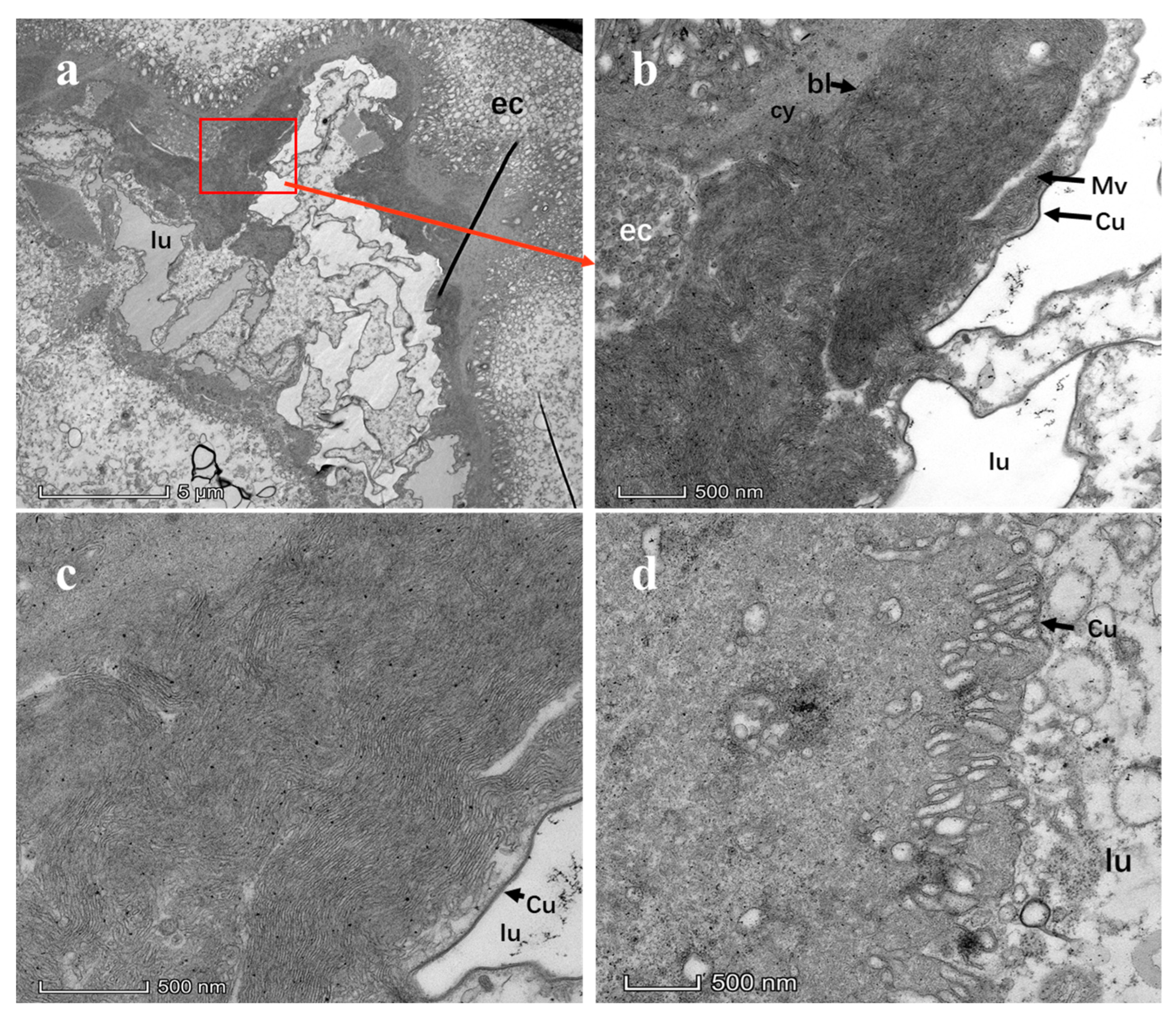
2.7. Midgut Dissection and Transmission Electron Microscopic Observation
2.8. Data Analysis
3. Results
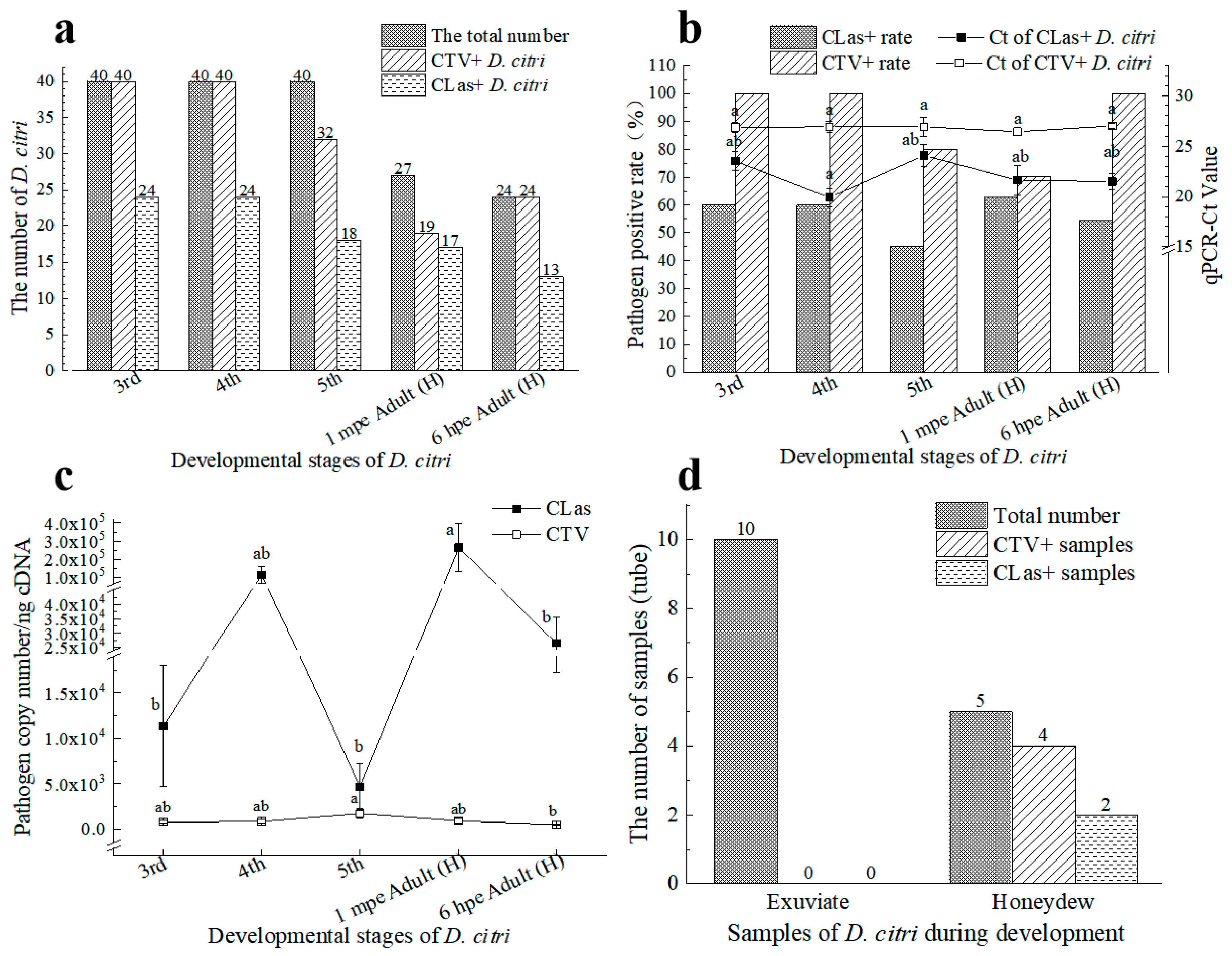
3.1. Incidence of CTV and CLas in D. citri from Citrus Orchards
3.2. CLas and CTV Acquisition by D. citri under Laboratory Conditions
3.2.1. CLas and CTV Acquisition on Citrus Trees Infected by Both Pathogens
3.2.2. CLas Facilitated the CTV Acquisition by D. citri in the Early Stage and Inhibited the CTV Acquisition Later
3.2.3. CLas Acquisition by D. citri was Promoted by CTV
3.2.4. CTV Contributed to the Transmission of CLas by D. citri
3.3. Ultrastructure of CTV in Midgut
3.4. CTV Replicates in Midgut of D. citri
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orchard Number | Sampling Time | Location | Orientation Information | Age of the Tree/Year-Old | Citrus Cultivars | Number of Trees Sampled |
|---|---|---|---|---|---|---|
| 1 | August, 2020 | Zhanjiang | 21°26′20.23″ N, 110°28′02.09″ E | 2 | Citrus sinensis Osbeck. ‘Hongjiangcheng’ | 10 |
| 2 | November, 2020 | Deqing | 23°13′21.56″ N, 111°53′57.89″ E | 10 | Citrus reticulata Blanco var. gonggan | 14 |
| 3 | November 2020 | Deqing | 23°19′21.91″ N, 112°12′16.52″ E | 4 | Citrus medica L. var. sarcodactylis Swingle | 16 |
| Orchard Number | Developmental Stages of D. citri (Quantity/Individual) | ||||||
|---|---|---|---|---|---|---|---|
| Egg | 1st | 2nd | 3rd | 4th | 5th | Adult | |
| 1 | 45 | 30 | 20 | 8 | 24 | 78 | 37 |
| 2 | 0 | 30 | 108 | 104 | 135 | 88 | 37 |
| 3 | 10 | 10 | 28 | 32 | 12 | 10 | 26 |
| Total | 55 | 70 | 156 | 144 | 171 | 176 | 100 |


References
- Garnier, M.; Jagoueix-Eveillard, S.; Cronje, P.R.; Roux, H.F.; Bové, J.M. Genomic characterization of a liberibacter present in an ornamental rutaceous tree, Calodendrum capense, in the western cape province of South Africa. Proposal of “Candidatus Liberibacter africanus subsp. capensis”. Int. J. Syst. Evol. Microbiol. 2000, 50, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Bové, J.M.; Barros, A.P. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Hu, B.; Rao, M.; Deng, X.; Pandey, S.S.; Hendrich, C.; Ding, F.; Wang, N.; Xu, Q. Molecular signatures between citrus and Candidatus Liberibacter asiaticus. PLoS Pathog. 2021, 17, e1010071. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.R.; Diniz, A.J.; Parra, J.R. Biology of the Huanglongbing vector Diaphorina citri (Hemiptera: Liviidae) on different host plants. J. Econ. Entomol. 2014, 107, 691–696. [Google Scholar] [CrossRef]
- Xie, P.; Su, C.; Lin, Z. Study on the biological characteristics of Diaphorina citri Kuwayama. J. Zhejiang Univ. 1989, 15, 198–202. [Google Scholar]
- EFSA Panel on Plant Health (PLH); Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.A.; Miret, J.A.J.; Justesen, A.F.; Magnusson, C.S.; Milonas, P.; et al. Pest categorisation of Diaphorina citri. EFSA J. 2021, 19, e06357. [Google Scholar]
- Inoue, H.; Ohnishi, J.; Ito, T.; Miyata, S.; Iwanami, T.; Ashihara, W. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Ann. Appl. Biol. 2009, 155, 29–36. [Google Scholar] [CrossRef]
- Pelz-Stelinski, K.S.; Brlansky, R.H.; Ebert, T.A.; Rogers, M.E. Transmission parameters for Candidatus Liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae). J. Econ. Entomol. 2010, 103, 1531–1541. [Google Scholar] [CrossRef]
- Wu, T.; Luo, X.; Qureshi, J.; Wu, F.; Xu, C.; Cen, Y. Feeding behavior of Diaphorina citri (Hemiptera: Liviidae) and its transmission of ‘Candidatus Liberibacter asiaticus’ to citrus. Entomol. Exp. Appl. 2016, 161, 104–111. [Google Scholar] [CrossRef]
- Buitendag, C.H.; Von Broembsen, L.A. Living with citrus greening in South Africa. Citrus 1993, 3, 29–32. [Google Scholar] [CrossRef]
- Mann, R.S.; Pelz-Stelinski, K.; Hermann, S.L.; Tiwari, S.; Stelinski, L.L. Sexual transmission of a plant pathogenic bacterium, Candidatus Liberibacter asiaticus, between conspecific insect vectors during mating. PLoS ONE 2011, 6, 29197. [Google Scholar] [CrossRef]
- Roistacher, C.N. Should we introduce protective isolates of Citrus tristeza virus. Citrograph 1992, 77, 5–9. [Google Scholar]
- Zhou, Y.; Zhou, C.; Wang, X.; Liu, K.; Liu, J.; Tang, K. Preliminary study on molecular characterization of 11 isolates of Citrus tristeza virus. Acta Phytopathol. Sin. 2005, 35, 147–150. [Google Scholar]
- Bar, J.M.; Marcus, R.; Lee, R.F. The continuous challenge of Citrus tristeza virus control. Annu. Rev. Phytopathol. 1989, 27, 291–316. [Google Scholar]
- Ruiz-Ruiz, S.; Moreno, P.; Guerri, J.; Ambrós, S. Discrimination between mild and severe Citrus tristeza virus isolates with a rapid and highly specific real-time reverse transcription-polymerase chain reaction method using TaqMan LNA probes. Phytopathology 2009, 99, 307–315. [Google Scholar] [CrossRef]
- Mehta, P.; Brlansky, R.H.; Gowda, S.; Yokomi, R.K. Reverse-transcription polymerase chain reaction detection of Citrus tristeza virus in aphids. Plant Dis. 1997, 81, 1066–1069. [Google Scholar] [CrossRef]
- Michaud, J.P. A review of the literature on Toxoptera citricida (Homoptera: Aphididae). Fla. Entomol. 1998, 81, 37–61. [Google Scholar] [CrossRef]
- Rocha, P.M. Citrus tristeza virus and its aphid vector Toxoptera citmicida: Threats to citrus production in the Caribbean and central and north America. Plant Dis. 1995, 79, 437–445. [Google Scholar] [CrossRef]
- Yokomi, R.K.; Damsteegt, V.D. Comparison of Citrus tristeza virus transmission efficiency between Toxoptera citricidus and Aphis gossypii. Misc. Publ. -Agric. Exp. Stn. Okla. State Univ. 1991, 132, 319. [Google Scholar]
- Liao, W.; Lin, W.; Yang, X.; Zeng, B.; Chen, P.; Kong, Y. Advances in detection and control methods of Citrus tristeza disease. J. Anhui Agric. Sci. 2012, 397, 17559–17561. [Google Scholar]
- Zhang, H.; Chen, J.; Lei, J.; Adams, M.J. Sequence analysis shows that a dwarfing disease on rice, wheat and maize in China is caused by rice black-streaked dwarf virus. Eur. J. Plant Pathol. 2001, 107, 563–567. [Google Scholar] [CrossRef]
- Achon, M.A.; Subira, J.; Sin, E. Seasonal occurrence of Laodelphax striatellus in Spain: Effect on the incidence of Maize rough dwarf virus. Crop Prot. 2013, 47, 1–5. [Google Scholar] [CrossRef]
- Deng, J.; Li, S.; Hong, J.; Ji, Y.; Zhou, Y. Investigation on subcellular localization of Rice stripe virus in its vector small brown planthopper by electron microscopy. Virol. J. 2013, 10, 310. [Google Scholar] [CrossRef] [PubMed]
- Di, D.; Zhang, Y.; Yan, C.; Yan, T.; Zhang, A.; Yang, F.; Cao, X.; Li, D.; Lu, Y.; Wang, X.; et al. First report of Barley yellow striate mosaic virus on wheat in China. Plant Dis. 2014, 98, 1450–1451. [Google Scholar] [CrossRef] [PubMed]
- Tanno, F.; Nakatsu, A.; Toriyama, S.; Kojima, M. Complete nucleotide sequence of Northern cereal mosaic virus and its genome organization. Arch. Virol. 2000, 145, 1373–1384. [Google Scholar] [CrossRef]
- Toriyama, S.; Guy, P.L.; Fuji, S.; Takahashi, M. Characterization of a new picorna-like virus, himetobi P virus, in planthoppers. J. Gen. Virol. 1992, 73, 1021–1023. [Google Scholar] [CrossRef]
- Syller, J. Biological and molecular events associated with simultaneous transmission of plant viruses by invertebrate and fungal vectors. Mol. Plant Pathol. 2014, 15, 417–426. [Google Scholar] [CrossRef]
- Moreno, A.B.; López-Moya, J.J. When viruses play team sports: Mixed infections in plants. Phytopathology 2020, 110, 29–48. [Google Scholar] [CrossRef]
- Wang, H.; Xu, D.; Pu, L.; Zhou, G. Southern rice black-streaked dwarf virus alters insect vectors’ host orientation preferences to enhance spread and increase rice ragged stunt virus co-infection. Phytopathology 2014, 104, 196–201. [Google Scholar] [CrossRef]
- Kliot, A.; Cilia, M.; Czosnek, H.; Ghanim, M. Implication of the bacterial endosymbiont Rickettsia spp. in interactions of the whitefly Bemisia tabaci with Tomato Yellow Leaf Curl Virus. J. Virol. 2014, 88, 5652–5660. [Google Scholar] [CrossRef]
- Dumón, A.D.; Argüello-Caro, E.B.; Mattio, M.F.; Alemandri, V.; Vas, M.D.; Truol, G. Co-infection with a wheat rhabdovirus causes a reduction in Mal de Río Cuarto virus titer in its planthopper vector. Bull. Entomol. Res. 2018, 108, 232–240. [Google Scholar] [CrossRef]
- Britt, K.; Gebben, S.; Levy, A.; Rwahnih, M.A. The detection and surveillance of Asian Citrus Psyllid (Diaphorina citri)-associated viruses in Florida citrus groves. Front. Plant Sci. 2020, 10, 1687. [Google Scholar] [CrossRef]
- Wu, F.; Huang, M.; Fox, E.G.P.; Huang, J.; Cen, Y.; Deng, X.; Xu, M. Preliminary Report on the acquisition, persistence, and potential transmission of Citrus tristeza virus by Diaphorina citri. Insects 2021, 12, 735. [Google Scholar] [CrossRef]
- Cui, Y.; Peng, A.; Song, X.; Cheng, B.; Lin, J.; Chen, X. Investigation on the occurrence of Huanglongbing and virus disease and the diversity of phage of pathogen of Huanglongbing in Meizhou, Guangdong province. Sci. Agric. Sin. 2020, 53, 78–88. [Google Scholar]
- Liu, H.; Wang, Z.; Cao, Y.; Xia, Y.; Yin, Y. Detection of Citrus tristeza virus using conventional and fluorescence quantitative RT-PCR assays. Acta Phytopathol. Sin. 2008, 1, 12. [Google Scholar]
- Bao, M.; Zheng, Z.; Sun, X.; Chen, J.; Deng, X. Enhancing PCR capacity to detect ‘Candidatus Liberibacter asiaticus’ utilizing whole genome sequence information. Plant Dis. 2020, 104, 527–532. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Ammar, E.D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef]
- Ng, J.C.; Zhou, J.S. Insect vector-plant virus interactions associated with non-circulative, semi-persistent transmission: Current perspectives and future challenges. Curr. Opin. Virol. 2015, 15, 48–55. [Google Scholar] [CrossRef]
- Bhagabati, K.N.; Nariana, T. Interaction of greening and tristeza pathogens in Kagzi lime (Citrus aurantifolia (Christm.) Swing.) and their effect on growth and development of disease symptoms. Indian Phytopathol. 1980, 33, 292–295. [Google Scholar]
- Kim, J.S.; Sagaram, U.S.; Burns, J.K.; Li, J.; Wang, N. Response of sweet orange (Citrus sinensis) to “Candidatus Liberibacter asiaticus” infection: Microscopy and microarray analyses. Phytopathology 2009, 99, 50–57. [Google Scholar] [CrossRef]
- Tao, Z.; Yi, L.; Lu, Z.; Li, F.; Lai, X.; Zhong, B. Investigation on the incidence of Citrus tristeza disease in navel orange main producing areas in Southern Jiangxi. Chin. Agric. Sci. Bull. 2011, 27, 297–300. [Google Scholar]
- Cui, Y.; Peng, A.; Li, Z.; Lin, J.; Song, X.; Guo, Z. Preliminary report on the occurrence and harm of Huanglongbing and virus disease in the main citrus producing areas of Hainan Province. Plant Prot. 2019, 45, 236–242. [Google Scholar]
- Wisler, G.C.; Li, R.; Liu, H.; Duffus, J.E. Tomato chlorosis virus: A new whitefly-transmitted, phloemlimited, bipartite closterovirus of tomato. Phytopathology 1998, 88, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Dai, F.; Zhou, X. First report of Tomato yellow leaf curl virus in China. Plant Dis. 2006, 90, 1359. [Google Scholar] [CrossRef]
- Dumón, A.D.; Argüello-Caro, E.B.; Alemandri, V.; Bainotti, C.; Mattio, F.; Rodriauez, S.M. Identificación y caracterización biológica del Barley yellow striate mosaic virus (BYSMV): Nueva enfermedad del trigo en Argentina. Trop. Plant Pathol. 2011, 36, 374–382. [Google Scholar] [CrossRef]
- Dumón, A.D.; Mattio, F.; Argüello-Caro, E.B.; Alemandri, V.; Puyane, N.; Vas, M.D. Occurrence of a closely-related isolate to Maize yellow striate virus in wheat plants. Agriscientia 2015, 32, 107–112. [Google Scholar] [CrossRef]
- Ding, T.; Li, J.; Chen, E.; Niu, J.; Dong, C. Transcriptome profiling of the whitefly Bemisia tabaci MED in response to single infection of Tomato yellow leaf curl virus, Tomato chlorosis virus, and their co-infection. Front. Physiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- Canale, M.C.; Tomaseto, A.F.; Haddad, M.D.; Spotti, L.J.R.; Fernando, T.A. Latency and persistence of ‘Candidatus Liberibacter asiaticus’ in its psyllid vector, Diaphorina citri (Hemiptera: Liviidae). Phytopathology 2017, 107, 264–272. [Google Scholar] [CrossRef]
- Ren, S.; Ou, D.; Zhang, L.; Sang, W.; Ji, Q.; Qiu, B. Effects of different host plants on the development and propagation of Diaphorina citri Kuwayama. Chin. J. Appl. Entomol. 2018, 55, 6. [Google Scholar]
- Wintermantel, W.M.; Cortez, A.A.; Anchieta, A.G.; Gulati-Sakhuja, A.; Hladky, L.L. Co-infection by two criniviruses alters accumulation of each virus in a host-specific manner and influences efficiency of virus transmission. Phytopathology 2008, 98, 1340–1345. [Google Scholar] [CrossRef]
- McElhany, P.; Real, L.A.; Power, A.G. Vector preference and disease dynamics: A study of barley yellow dwarf virus. Ecology 1995, 76, 444–457. [Google Scholar] [CrossRef]
- Mauck, K.E.; De-Moraes, C.M.; Mescher, M.C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. USA 2010, 107, 3600–3605. [Google Scholar] [CrossRef]
- Zhao, P.; Yao, X.; Cai, C.; Li, R.; Du, J.; Sun, Y. Viruses mobilize plant immunity to deter nonvector insect herbivores. Adv. Sci. 2019, 5, eaav9801. [Google Scholar] [CrossRef]
- Cen, Y.; Yang, C.; Holford, P.; Beattie, G.A.C.; Spooner-Hart, R.N.; Liang, G. Feeding behaviour of the Asiatic Citrus Psyllid, Diaphorina citri, on healthy and Huanglongbing-infected citrus. Entomol. Exp. Appl. 2012, 143, 13–22. [Google Scholar] [CrossRef]
- Salvaudon, L.; De-Moraes, C.M.; Mescher, M.C. Outcomes of co-infection by two potyviruses: Implications for the evolution of manipulative strategies. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122959. [Google Scholar] [CrossRef]
- Dalmon, A.; Bouyer, S.; Cailly, M.; Girard, M.; Lecoq, H.; Desbiez, C. First report of Tomato chlorosis virus and Tomato infectious chlorosis virus in tomato crops in France. Plant Dis. 2005, 89, 1243. [Google Scholar] [CrossRef]
- Glaser, R.L.; Meola, M.A. The native Wolbachia endosymbionts of Drosophila melanogaster and Culex quinquefasciatus increase host resistance to West Nile virus infection. PLoS ONE 2010, 5, e11977. [Google Scholar] [CrossRef]
- Zouache, K.; Michelland, R.J.; Failloux, A.B.; Grundmann, G.L.; Mavingui, P. Chikungunya virus impacts the diversity of symbiotic bacteria in mosquito vector. Mol. Ecol. 2012, 21, 2297–2309. [Google Scholar] [CrossRef]
- Yan, F. Plant pathogen-vector insects interactions: Advances and Prospects. Acta Entomol. Sin. 2020, 63, 8. [Google Scholar]







| Citrus Cultivars | Pathogen Carried by Scions | Ct Value of CLas | Ct Value of CTV |
|---|---|---|---|
| Citrus reticulata Blanco “Shatangju” | CLas and CTV | 24.27 ± 1.01 | 22.17 ± 0.44 |
| Citrus tangerine Hort. Ex Tanaka | CLas | 23.21 ± 0.83 | 35.49 ± 0.72 |
| Citrus grandis Osbeck | CTV | 37.41 ± 1.44 | 23.01 ± 1.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Liu, Y.; Wu, F.; Zhang, J.; Cui, X.; Wu, S.; Deng, X.; Xu, M. Citrus tristeza virus Promotes the Acquisition and Transmission of ‘Candidatus Liberibacter Asiaticus’ by Diaphorina citri. Viruses 2023, 15, 918. https://doi.org/10.3390/v15040918
Chen L, Liu Y, Wu F, Zhang J, Cui X, Wu S, Deng X, Xu M. Citrus tristeza virus Promotes the Acquisition and Transmission of ‘Candidatus Liberibacter Asiaticus’ by Diaphorina citri. Viruses. 2023; 15(4):918. https://doi.org/10.3390/v15040918
Chicago/Turabian StyleChen, Longtong, Yangyang Liu, Fengnian Wu, Jingtian Zhang, Xiaoqing Cui, Shitong Wu, Xiaoling Deng, and Meirong Xu. 2023. "Citrus tristeza virus Promotes the Acquisition and Transmission of ‘Candidatus Liberibacter Asiaticus’ by Diaphorina citri" Viruses 15, no. 4: 918. https://doi.org/10.3390/v15040918
APA StyleChen, L., Liu, Y., Wu, F., Zhang, J., Cui, X., Wu, S., Deng, X., & Xu, M. (2023). Citrus tristeza virus Promotes the Acquisition and Transmission of ‘Candidatus Liberibacter Asiaticus’ by Diaphorina citri. Viruses, 15(4), 918. https://doi.org/10.3390/v15040918