
Characterization of Virulent T4-Like Acinetobacter baumannii Bacteriophages DLP1 and DLP2


 ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria and Bacteriophages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organisms | Phase and Description | Source |
|---|---|---|
| A. baumannii AB5075cm | Capsule mutant AB5075; host for DLP1 and DLP2 | This study |
| A. baumannii AB5075O | AB5075; DLP1R and DLP2R | [19] |
| A. baumannii AB5075O Δwzc | Clean deletion of wzc in AB5075O; DLP1S and DLP2S | [19] |
| A. baumannii AB5075O Δwzc/carO::MarTc | Clean deletion of wzc and Tc1/mariner transposon insertion of carO in AB5075O; DLP1S and DLP2R | This study |
| A. baumannii AB5075O Δwzc/carO::T26 | Clean deletion of wzc and T26 transposon insertion of carO in AB5075O; DLP1S and DLP2R | This study |
| pQF1266Blue | Broad-host-range plasmid | [20] |
| pQF1266Blue + carO | Broad-host-range plasmid encoding carO gene with native RBS | This study |
| A. baumannii 17978O | South Korean isolate ATCC 17978 | |
| A. baumannii AB0057O | Walter Reed Army Medical Center | [21] |
| A. baumannii AYEO | French epidemic strain with high mortality | [22] |
| A. baumannii 19606O | ATCC 19606-type strain | [23] |
| A. baumannii AB030cm | Hypervirulent XDR Canadian isolate | [24] |
| A. baumannii LAC-4O | Hypervirulent XDR American isolate | [25] |
| A. baumannii MRSN 7460 | Human wound isolate | [26] |
| A. baumannii MRSN 11669 | Human urine isolate | [26] |
| A. baumannii MRSN 15088 | Human wound isolate | [26] |
| A. baumannii MRSN 15093 | Human wound isolate | [26] |
| A. baumannii MRSN 17493 | Human respiratory isolate | [26] |
| A. baumannii MRSN 21681 | Human catheter isolate | [26] |
| A. baumannii MRSN 22112 | Human blood isolate | [26] |
| A. baumannii MRSN 24603 | Human blood isolate | [26] |
| A. baumannii MRSN 30885 | Human respiratory isolate | [26] |
| A. baumannii MRSN 31159 | Human tissue sample | [26] |
| A. baumannii MRSN 31461 | Unknown | [26] |
| A. baumannii MRSN 31915 | Unknown | [26] |
| A. baumannii MRSN 31937 | Human wound isolate | [26] |
| A. baumannii MRSN 32304 | Unknown | [26] |
| A. baumannii MRSN 32875 | Unknown | [26] |
| A. baumannii MRSN 32915 | Human wound isolate | [26] |
| A. baumannii MRSN 351524 | Human blood isolate | [26] |
| A. baumannii MRSN 423159 | Human respiratory isolate | [26] |
| A. baumannii MRSN 480561 | Human respiratory isolate | [26] |
| A. baumannii MRSN 480622 | Human urine isolate | [26] |
| A. baumannii MRSN 489669 | Human respiratory isolate | [26] |
| vB_AbaM_DLP1 | Lytic myovirus DLP1 | This study |
| vB_AbaM_DLP2 | Lytic myovirus DLP2 | This study |
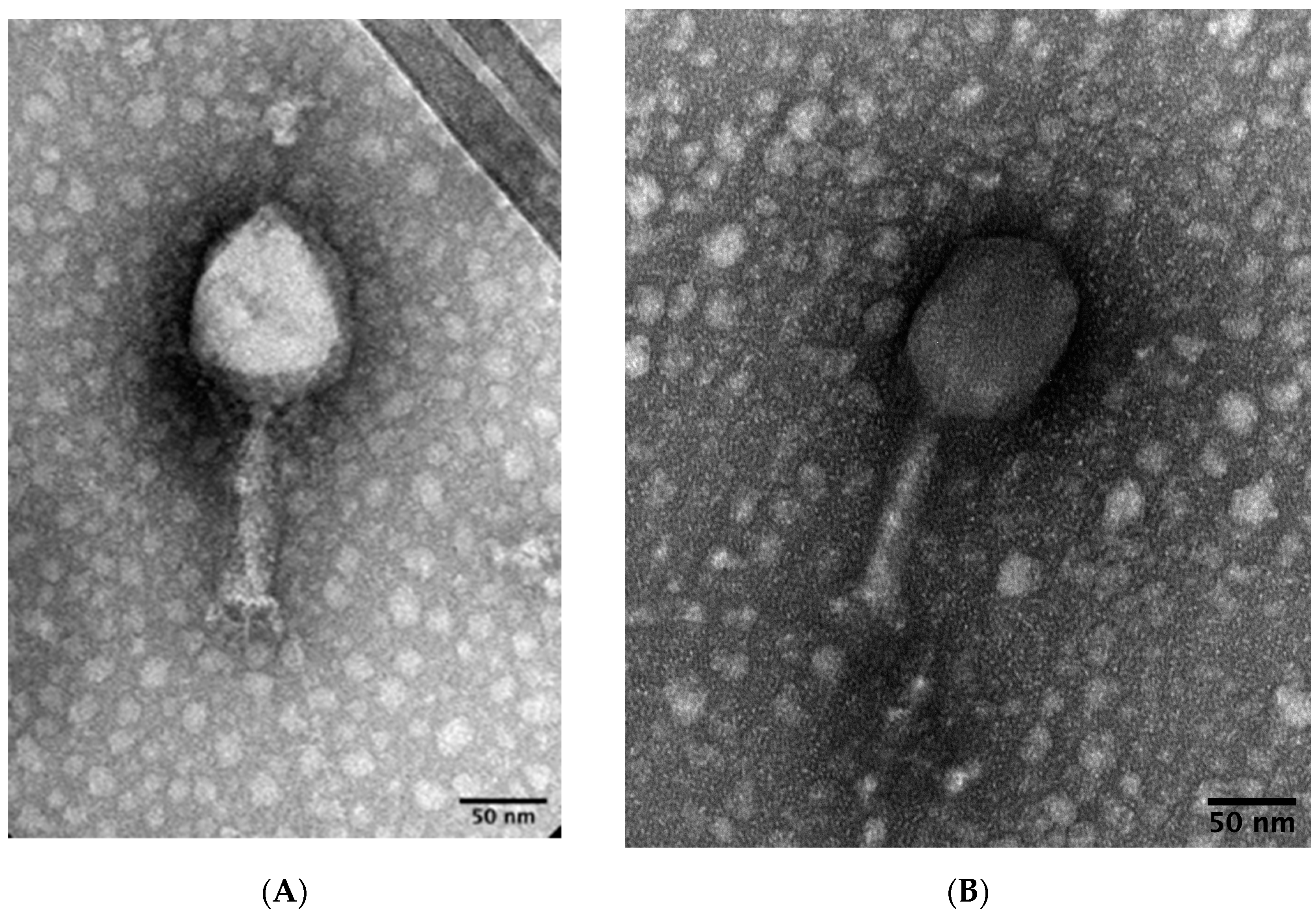
2.2. Transmission Electron Microscopy (TEM)
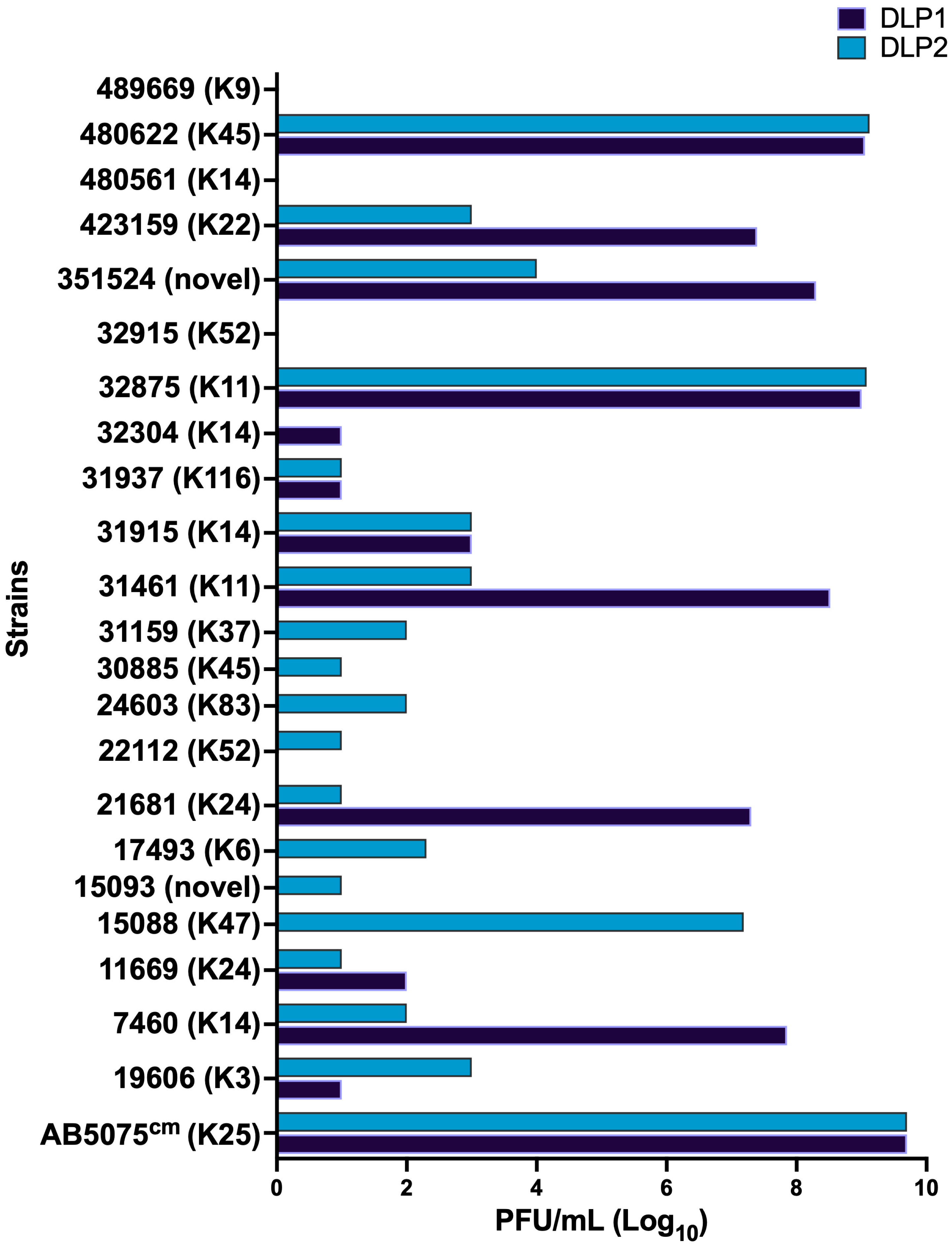
2.3. Host Range and Efficiency of Plating (EOP)
2.4. One-Step Growth Curves
2.5. Virulence Assay of DLP1 and DLP2 against AB5075cm
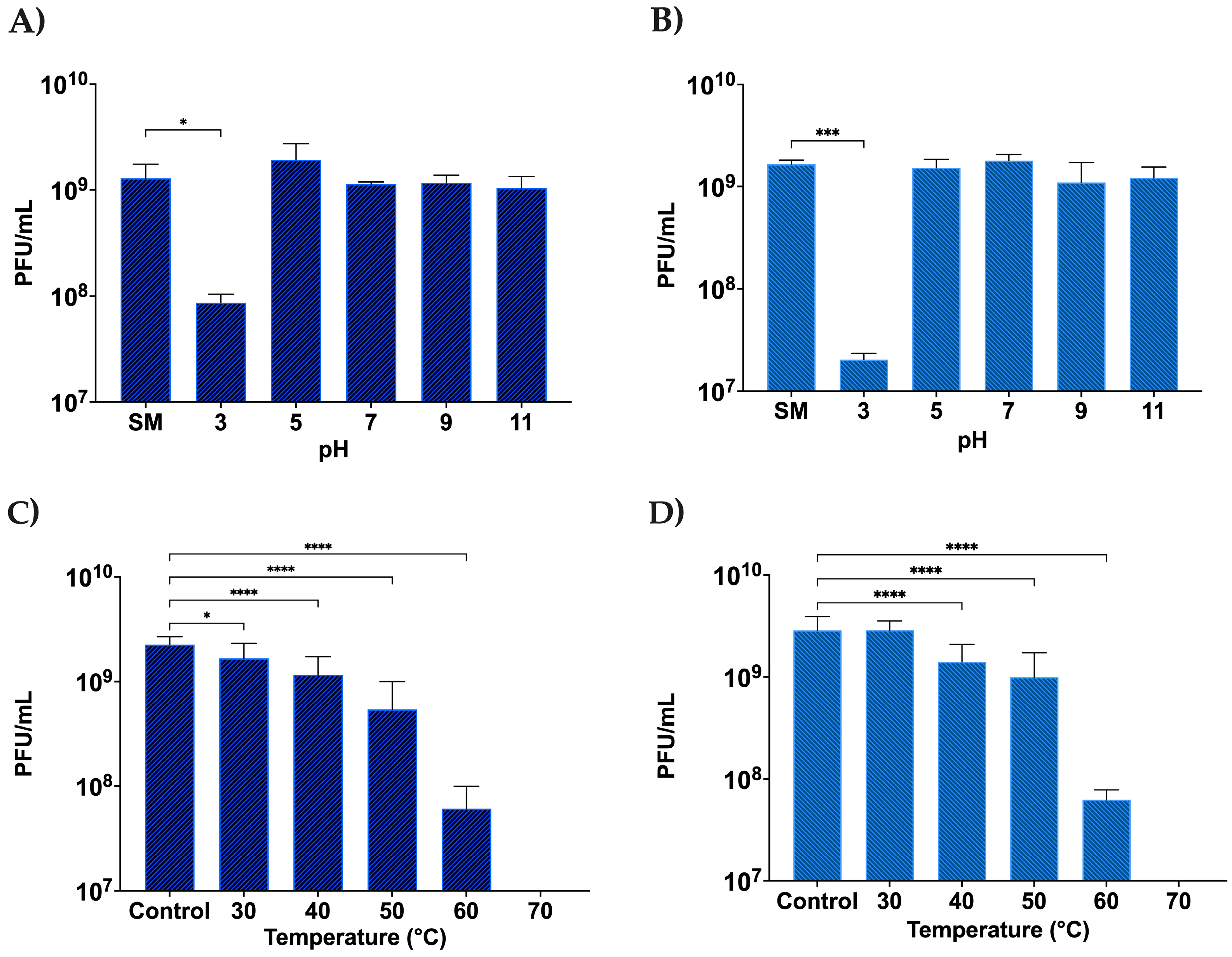
2.6. Phage pH Stability Assay
2.7. Phage Temperature Stability Assay
2.8. Biofilm-Inhibition Assessment of DLP1 and DLP2
2.9. Identification of DLP2 Receptor
2.10. Complementation of the carO Mutant
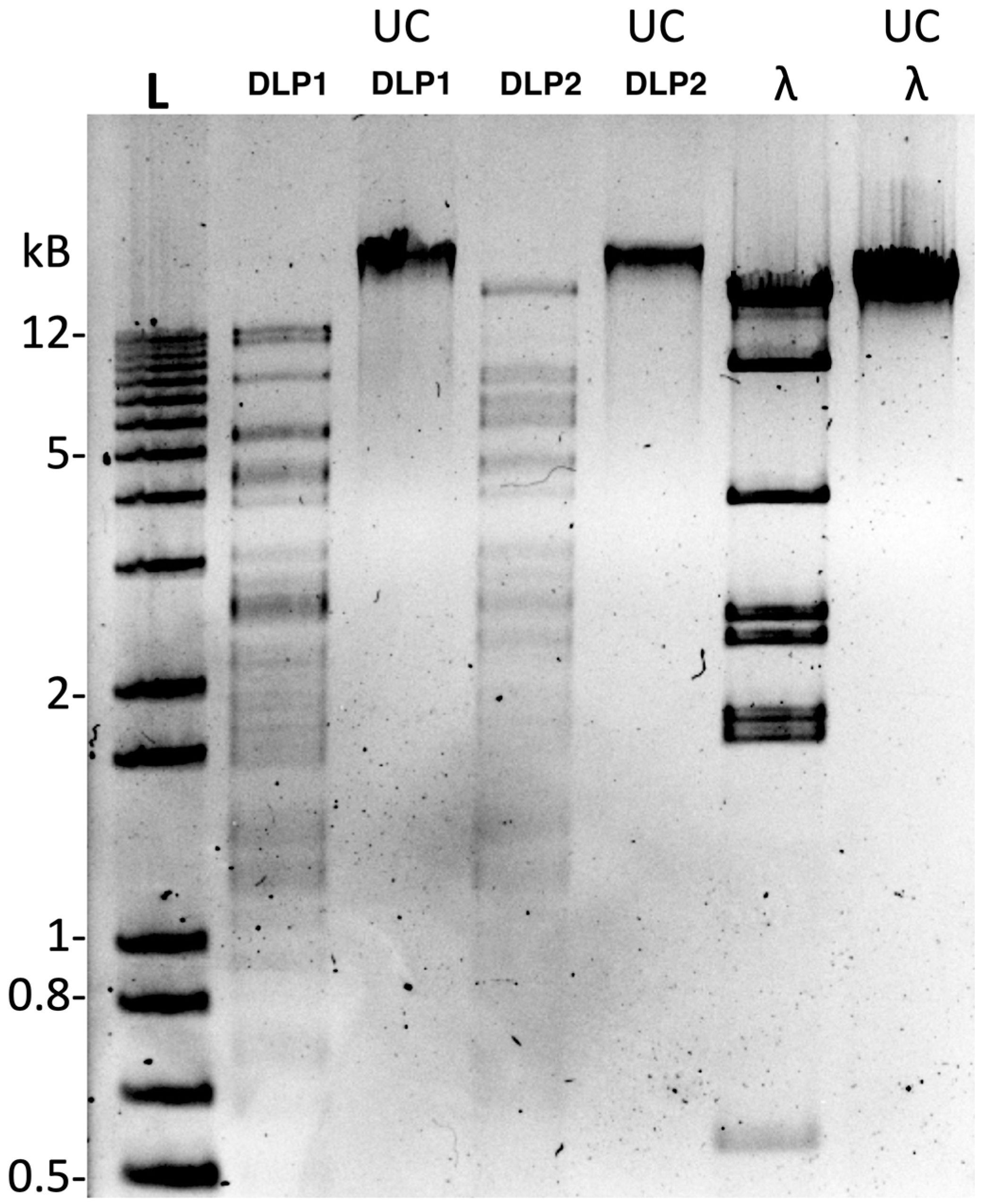
2.11. Phage Concentration, DNA Isolation, Sequencing, and RFLP Analysis
2.12. Bioinformatics
2.13. Statistical Analysis
3. Results
3.1. Phage Isolation, Morphology, and Host Range
3.1.1. Phage Isolation and Morphology
3.1.2. Host Range and Efficiency of Plating (EOP)
3.2. Burst Size, Virulence and Stability of DLP1 and DLP2
3.2.1. Burst Size
3.2.2. AB5075cm Suppression and Virulence Curves of DLP1 and DLP2
3.2.3. Biofilm Inhibition of DLP1 and DLP2
3.2.4. Stability of DLP1 and DLP2 across a pH and Temperature Range
3.2.5. Identification of DLP2 Receptor
3.3. Genomic Analysis
3.3.1. Restriction Fragment Length Polymorphism Analysis (RFLP)
3.3.2. Genome Features of DLP1 and DLP2
3.3.3. Comparison of DLP1 and DLP2 Genomes and Tail Fibers
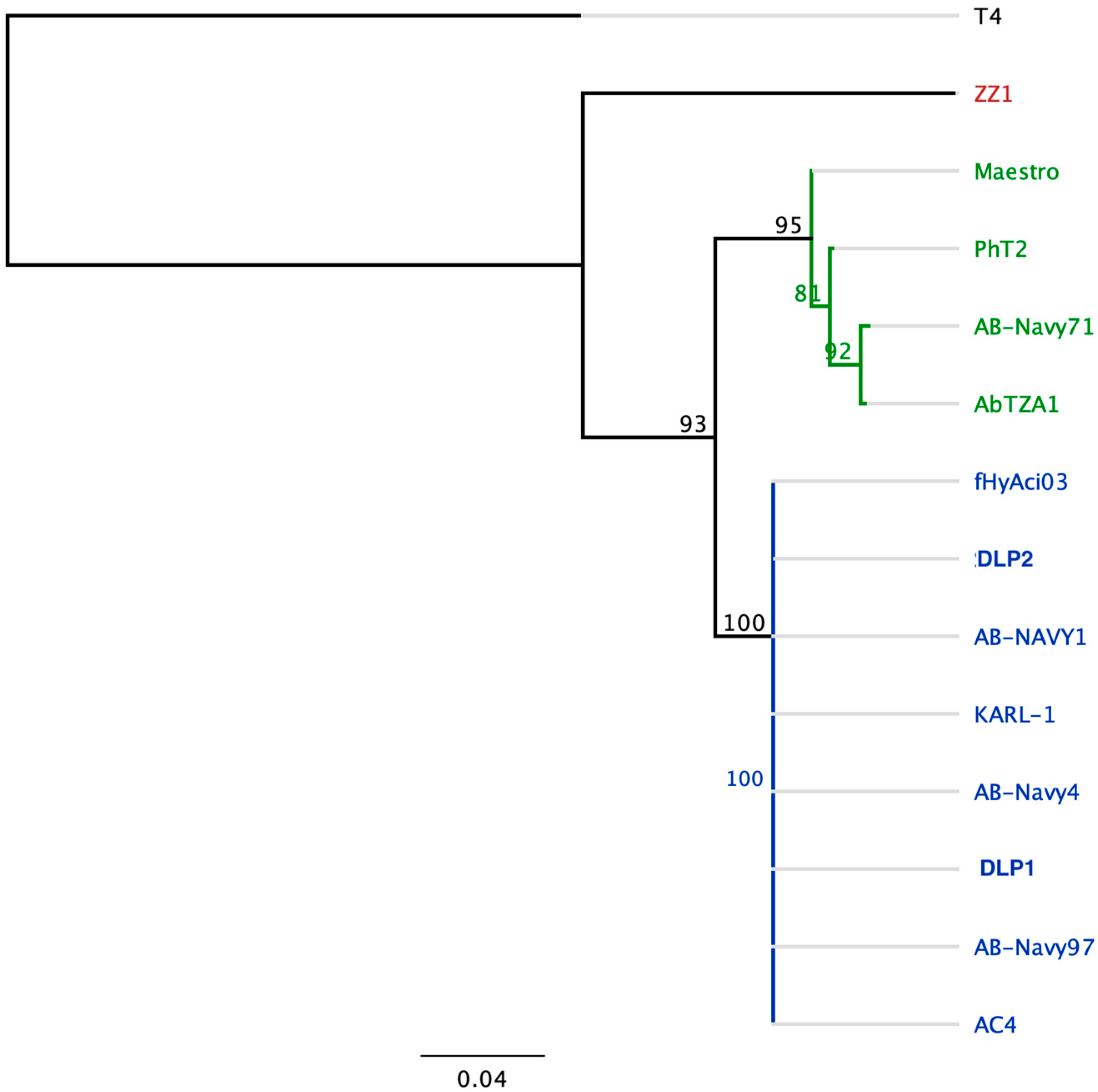
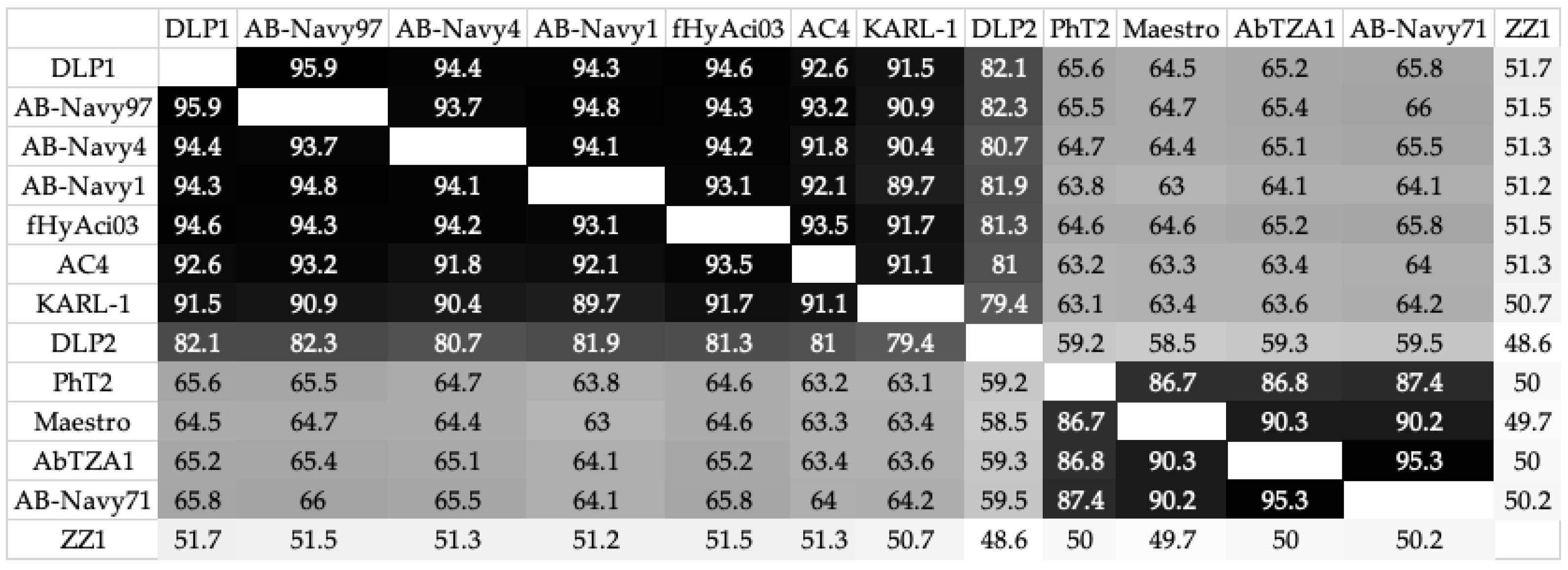
3.3.4. Genomic Analysis of DLP1 and DLP2 against Published Acinetobacter Phages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- CDC. COVID-19: U.S. Impact on Antimicrobial Resistance, Special Report 2022; National Center for Emerging and Zoonotic Infectious Diseases: Atlanta, GA, USA, 2022. [Google Scholar] [CrossRef]
- World Health Organization. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report: 2021; World Health Organization: Geneva, Switzerland, 2021; ISBN 978-92-4-002733-6. [Google Scholar]
- Lee, C.-R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.-J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Ayoub Moubareck, C.; Hammoudi Halat, D. Insights into Acinetobacter baumannii: A Review of Microbiological, Virulence, and Resistance Traits in a Threatening Nosocomial Pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Kakasis, A.; Panitsa, G. Bacteriophage Therapy as an Alternative Treatment for Human Infections. A Comprehensive Review. Int. J. Antimicrob. Agents 2019, 53, 16–21. [Google Scholar] [CrossRef]
- Ross, A.; Ward, S.; Hyman, P. More Is Better: Selecting for Broad Host Range Bacteriophages. Front. Microbiol. 2016, 7, 1352. [Google Scholar] [CrossRef]
- Vandersteegen, K.; Kropinski, A.M.; Nash, J.H.E.; Noben, J.-P.; Hermans, K.; Lavigne, R. Romulus and Remus, Two Phage Isolates Representing a Distinct Clade within the Twortlikevirus Genus, Display Suitable Properties for Phage Therapy Applications. J. Virol. 2013, 87, 3237–3247. [Google Scholar] [CrossRef]
- Yehl, K.; Lemire, S.; Yang, A.C.; Ando, H.; Mimee, M.; Torres, M.D.T.; de la Fuente-Nunez, C.; Lu, T.K. Engineering Phage Host-Range and Suppressing Bacterial Resistance through Phage Tail Fiber Mutagenesis. Cell 2019, 179, 459–469. [Google Scholar] [CrossRef]
- Dams, D.; Brøndsted, L.; Drulis-Kawa, Z.; Briers, Y. Engineering of Receptor-Binding Proteins in Bacteriophages and Phage Tail-like Bacteriocins. Biochem. Soc. Trans. 2019, 47, 449–460. [Google Scholar] [CrossRef]
- Burrowes, B.; Molineux, I.; Fralick, J. Directed in Vitro Evolution of Therapeutic Bacteriophages: The Appelmans Protocol. Viruses 2019, 11, 241. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter baumannii Infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef]
- LaVergne, S.; Hamilton, T.; Biswas, B.; Kumaraswamy, M.; Schooley, R.T.; Wooten, D. Phage Therapy for a Multidrug-Resistant Acinetobacter baumannii Craniectomy Site Infection. Open Forum Infect. Dis. 2018, 5, ofy064. [Google Scholar] [CrossRef] [PubMed]
- Nir-Paz, R.; Gelman, D.; Khouri, A.; Sisson, B.M.; Fackler, J.; Alkalay-Oren, S.; Khalifa, L.; Rimon, A.; Yerushalmy, O.; Bader, R.; et al. Successful Treatment of Antibiotic-Resistant, Poly-Microbial Bone Infection With Bacteriophages and Antibiotics Combination. Clin. Infect. Dis. 2019, 69, 2015–2018. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Dai, J.; Guo, M.; Li, J.; Zhou, X.; Li, F.; Gao, Y.; Qu, H.; Lu, H.; Jin, J.; et al. Pre-Optimized Phage Therapy on Secondary Acinetobacter baumannii Infection in Four Critical COVID-19 Patients. Emerg. Microbes Infect. 2021, 10, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Chen, H.; Zhang, M.; Zhao, Y.; Jiang, Y.; Liu, X.; Huang, W.; Ma, Y. Clinical Experience of Personalized Phage Therapy Against Carbapenem-Resistant Acinetobacter baumannii Lung Infection in a Patient With Chronic Obstructive Pulmonary Disease. Front. Cell. Infect. Microbiol. 2021, 11, 631585. [Google Scholar] [CrossRef]
- Liu, M.; Hernandez-Morales, A.; Clark, J.; Le, T.; Biswas, B.; Bishop-Lilly, K.A.; Henry, M.; Quinones, J.; Voegtly, L.J.; Cer, R.Z.; et al. Comparative Genomics of Acinetobacter baumannii and Therapeutic Bacteriophages from a Patient Undergoing Phage Therapy. Nat. Commun. 2022, 13, 3776. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.L.; Harris, G.; Davis, C.M.; Dennis, J.J.; Chen, W. Bacteriophage Isolation, Purification, and Characterization Techniques Against Ubiquitous Opportunistic Pathogens. Curr. Protoc. 2022, 2, e594. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.Y.; Tipton, K.A.; Farokhyfar, M.; Burd, E.M.; Weiss, D.S.; Rather, P.N. A High-Frequency Phenotypic Switch Links Bacterial Virulence and Environmental Survival in Acinetobacter baumannii. Nat. Microbiol. 2018, 3, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.E.; Chin, C.Y.; Weiss, D.S.; Rather, P.N. Copy Number of an Integron-Encoded Antibiotic Resistance Locus Regulates a Virulence and Opacity Switch in Acinetobacter baumannii AB5075. mBio 2020, 11, e02338-20. [Google Scholar] [CrossRef]
- Adams, M.D.; Goglin, K.; Molyneaux, N.; Hujer, K.M.; Lavender, H.; Jamison, J.J.; MacDonald, I.J.; Martin, K.M.; Russo, T.; Campagnari, A.A.; et al. Comparative Genome Sequence Analysis of Multidrug-Resistant Acinetobacter baumannii. J. Bacteriol. 2008, 190, 8053–8064. [Google Scholar] [CrossRef]
- Fournier, P.-E.; Vallenet, D.; Barbe, V.; Audic, S.; Ogata, H.; Poirel, L.; Richet, H.; Robert, C.; Mangenot, S.; Abergel, C.; et al. Comparative Genomics of Multidrug Resistance in Acinetobacter baumannii. PLoS Genet. 2006, 2, e7. [Google Scholar] [CrossRef]
- Tsubouchi, T.; Suzuki, M.; Niki, M.; Oinuma, K.-I.; Niki, M.; Kakeya, H.; Kaneko, Y. Complete Genome Sequence of Acinetobacter baumannii ATCC 19606T, a Model Strain of Pathogenic Bacteria Causing Nosocomial Infection. Microbiol. Resour. Announc. 2020, 9, e00289-20. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; De Silva, P.M.; Al-Saadi, Y.; Switala, J.; Loewen, P.C.; Hausner, G.; Chen, W.; Hernandez, I.; Castillo-Ramirez, S.; Kumar, A. Characterization of Extremely Drug-Resistant and Hypervirulent Acinetobacter baumannii AB030. Antibiotics 2020, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.-Y.; Kuang, S.N.; He, X.; Molgora, B.M.; Ewing, P.J.; Deng, Z.; Osby, M.; Chen, W.; Xu, H.H. Complete Genome Sequence of Hypervirulent and Outbreak-Associated Acinetobacter baumannii Strain LAC-4: Epidemiology, Resistance Genetic Determinants and Potential Virulence Factors. Sci. Rep. 2015, 5, 8643. [Google Scholar] [CrossRef] [PubMed]
- Galac, M.R.; Snesrud, E.; Lebreton, F.; Stam, J.; Julius, M.; Ong, A.C.; Maybank, R.; Jones, A.R.; Kwak, Y.I.; Hinkle, K.; et al. A Diverse Panel of Clinical Acinetobacter baumannii for Research and Development. Antimicrob. Agents Chemother. 2020, 64, e00840-20. [Google Scholar] [CrossRef] [PubMed]
- Barth, O.M. The Use of Polylysine during Negative Staining of Viral Suspensions. J. Virol. Methods 1985, 11, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.C. Use of Polylysine for Adsorption of Nuclei Acids and Enzymes to Electron Microscope Specimen Films. Proc. Natl. Acad. Sci. USA 1977, 74, 2311–2315. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.J.; Kropinski, A.M. (Eds.) Bacteriophages; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2009; Volume 501, ISBN 978-1-58829-682-5. [Google Scholar]
- Davis, C.M.; Ruest, M.K.; Cole, J.H.; Dennis, J.J. The Isolation and Characterization of a Broad Host Range Bcep22-like Podovirus JC1. Viruses 2022, 14, 938. [Google Scholar] [CrossRef]
- Tipton, K.A.; Chin, C.-Y.; Farokhyfar, M.; Weiss, D.S.; Rather, P.N. Role of Capsule in Resistance to Disinfectants, Host Antimicrobials, and Desiccation in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2018, 62, e01188-18. [Google Scholar] [CrossRef]
- Liberati, N.T.; Urbach, J.M.; Miyata, S.; Lee, D.G.; Drenkard, E.; Wu, G.; Villanueva, J.; Wei, T.; Ausubel, F.M. An Ordered, Nonredundant Library of Pseudomonas aeruginosa Strain PA14 Transposon Insertion Mutants. Proc. Natl. Acad. Sci. USA 2006, 103, 2833–2838. [Google Scholar] [CrossRef]
- Gallagher, L.A.; Ramage, E.; Weiss, E.J.; Radey, M.; Hayden, H.S.; Held, K.G.; Huse, H.K.; Zurawski, D.V.; Brittnacher, M.J.; Manoil, C. Resources for Genetic and Genomic Analysis of Emerging Pathogen Acinetobacter baumannii. J. Bacteriol. 2015, 197, 2027–2035. [Google Scholar] [CrossRef]
- Asif, M.; Alvi, I.A.; Tabassum, R.; Rehman, S.U. TAC1, an Unclassified Bacteriophage of the Family Myoviridae Infecting Acinetobacter baumannii with a Large Burst Size and a Short Latent Period. Arch. Virol. 2020, 165, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Seed, K.D.; Dennis, J.J. Isolation and Characterization of Bacteriophages of the Burkholderia cepacia Complex. FEMS Microbiol. Lett. 2005, 251, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.H.; Stothard, P.; Dennis, J.J. Comparative Analysis of Two Phenotypically-Similar but Genomically-Distinct Burkholderia Cenocepacia-Specific Bacteriophages. BMC Genomics 2012, 13, 223. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Rukayadi, Y.; Hasan, H.; Abdul-Mutalib, N.-A.; Jambari, N.N.; Hara, H.; Thung, T.Y.; Lee, E.; Radu, S. Isolation and Characterization of Six Vibrio Parahaemolyticus Lytic Bacteriophages From Seafood Samples. Front. Microbiol. 2021, 12, 616548. [Google Scholar] [CrossRef]
- Lee, S.H.; Onuki, M.; Satoh, H.; Mino, T. Isolation, Characterization of Bacteriophages Specific to Microlunatus Phosphovorus and Their Application for Rapid Host Detection. Lett. Appl. Microbiol. 2006, 42, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.L.; Lynch, K.H.; Stothard, P.; Dennis, J.J. The Isolation and Characterization of Two Stenotrophomonas Maltophilia Bacteriophages Capable of Cross-Taxonomic Order Infectivity. BMC Genom. 2015, 16, 664. [Google Scholar] [CrossRef]
- Hallewell, J.; Niu, Y.D.; Munns, K.; McAllister, T.A.; Johnson, R.P.; Ackermann, H.-W.; Thomas, J.E.; Stanford, K. Differing Populations of Endemic Bacteriophages in Cattle Shedding High and Low Numbers of Escherichia Coli O157:H7 Bacteria in Feces. Appl. Environ. Microbiol. 2014, 80, 3819–3825. [Google Scholar] [CrossRef] [PubMed]
- Merabishvili, M.; Verhelst, R.; Glonti, T.; Chanishvili, N.; Krylov, V.; Cuvelier, C.; Tediashvili, M.; Vaneechoutte, M. Digitized Fluorescent RFLP Analysis (FRFLP) as a Universal Method for Comparing Genomes of Culturable DsDNA Viruses: Application to Bacteriophages. Res. Microbiol. 2007, 158, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Maszewska, A.; Wójcik, E.; Ciurzyńska, A.; Wojtasik, A.; Piątkowska, I.; Dastych, J.; Różalski, A. Differentiation of Polyvalent Bacteriophages Specific to Uropathogenic Proteus Mirabilis Strains Based on the Host Range Pattern and RFLP. Acta Biochim. Pol. 2016, 63, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.R.; Depardieu, F.; Fortier, L.-C.; Bikard, D.; Monot, M. PhageTerm: A Tool for Fast and Accurate Determination of Phage Termini and Packaging Mechanism Using next-Generation Sequencing Data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- McNair, K.; Zhou, C.; Dinsdale, E.A.; Souza, B.; Edwards, R.A. PHANOTATE: A Novel Approach to Gene Identification in Phage Genomes. Bioinformatics 2019, 35, 4537–4542. [Google Scholar] [CrossRef] [PubMed]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying Bacterial Genes and Endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.P.; Lowe, T.M. TRNAscan-SE: Searching for TRNA Genes in Genomic Sequences. In Gene Prediction; Kollmar, M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1962, pp. 1–14. ISBN 978-1-4939-9172-3. [Google Scholar]
- Sampaio, M.; Rocha, M.; Oliveira, H.; Dias, O. Predicting Promoters in Phage Genomes Using PhagePromoter. Bioinformatics 2019, 35, 5301–5302. [Google Scholar] [CrossRef]
- Kingsford, C.L.; Ayanbule, K.; Salzberg, S.L. Rapid, Accurate, Computational Discovery of Rho-Independent Transcription Terminators Illuminates Their Relationship to DNA Uptake. Genome Biol. 2007, 8, R22. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Konstantinidis, K.T. Bypassing Cultivation To Identify Bacterial Species: Culture-Independent Genomic Approaches Identify Credibly Distinct Clusters, Avoid Cultivation Bias, and Provide True Insights into Microbial Species. Microbe Mag. 2014, 9, 111–118. [Google Scholar] [CrossRef]
- Schwartz, S.; Kent, W.J.; Smit, A.; Zhang, Z.; Baertsch, R.; Hardison, R.C.; Haussler, D.; Miller, W. Human–Mouse Alignments with BLASTZ. Genome Res. 2003, 13, 103–107. [Google Scholar] [CrossRef]
- Harris, R. Improved Pairwise Alignment of Genomic DNA. Computer Science and Engineering. Ph.D. Thesis, Pennsylvania State University, State College, PA, USA, 2007. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal Omega for Making Accurate Alignments of Many Protein Sequences: Clustal Omega for Many Protein Sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.-H. Clinker & Clustermap.Js: Automatic Generation of Gene Cluster Comparison Figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef]
- Mussi, M.A.; Limansky, A.S.; Viale, A.M. Acquisition of Resistance to Carbapenems in Multidrug-Resistant Clinical Strains of Acinetobacter baumannii: Natural Insertional Inactivation of a Gene Encoding a Member of a Novel Family of β-Barrel Outer Membrane Proteins. Antimicrob. Agents Chemother. 2005, 49, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Mussi, M.A.; Relling, V.M.; Limansky, A.S.; Viale, A.M. CarO, an Acinetobacter baumannii Outer Membrane Protein Involved in Carbapenem Resistance, Is Essential for l-Ornithine Uptake. FEBS Lett. 2007, 581, 5573–5578. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Li, Z.J.; Wang, S.W.; Wang, S.M.; Huang, D.H.; Li, Y.H.; Ma, Y.Y.; Wang, J.; Liu, F.; Chen, X.D.; et al. Isolation and Characterization of ZZ1, a Novel Lytic Phage That Infects Acinetobacter baumannii Clinical Isolates. BMC Microbiol. 2012, 12, 156. [Google Scholar] [CrossRef]
- Mathews, C.K. Bacteriophage T4. In eLS; John Wiley & Sons, Ltd., Ed.; Wiley: New York, NY, USA, 2015; pp. 1–11. ISBN 978-0-470-01617-6. [Google Scholar]
- Roux, S.; Hallam, S.J.; Woyke, T.; Sullivan, M.B. Viral Dark Matter and Virus–Host Interactions Resolved from Publicly Available Microbial Genomes. eLife 2015, 4, e08490. [Google Scholar] [CrossRef]
- Abdelwahed, E.K.; Hussein, N.A.; Moustafa, A.; Moneib, N.A.; Aziz, R.K. Gene Networks and Pathways Involved in Escherichia Coli Response to Multiple Stressors. Microorganisms 2022, 10, 1793. [Google Scholar] [CrossRef] [PubMed]
- Pulkkinen, E.; Wicklund, A.; Oduor, J.M.O.; Skurnik, M.; Kiljunen, S. Characterization of VB_ApiM_fHyAci03, a Novel Lytic Bacteriophage That Infects Clinical Acinetobacter Strains. Arch. Virol. 2019, 164, 2197–2199. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.; Wahida, A.; Latz, S.; Krüttgen, A.; Häfner, H.; Buhl, E.M.; Ritter, K.; Horz, H.-P. Enhanced Antibacterial Effect of the Novel T4-like Bacteriophage KARL-1 in Combination with Antibiotics against Multi-Drug Resistant Acinetobacter baumannii. Sci. Rep. 2018, 8, 14140. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Li, Z.-J.; Wang, S.-W.; Wang, S.-M.; Chen, S.-J.; Huang, D.-H.; Zhang, G.; Li, Y.-H.; Wang, X.-T.; Wang, J.; et al. Genome Organisation of the Acinetobacter Lytic Phage ZZ1 and Comparison with Other T4-like Acinetobacter Phages. BMC Genom. 2014, 15, 793. [Google Scholar] [CrossRef] [PubMed]
- Styles, K.M.; Thummeepak, R.; Leungtongkam, U.; Smith, S.E.; Christie, G.S.; Millard, A.; Moat, J.; Dowson, C.G.; Wellington, E.M.H.; Sitthisak, S.; et al. Investigating Bacteriophages Targeting the Opportunistic Pathogen Acinetobacter baumannii. Antibiotics 2020, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Borin, J.M.; Avrani, S.; Barrick, J.E.; Petrie, K.L.; Meyer, J.R. Coevolutionary Phage Training Leads to Greater Bacterial Suppression and Delays the Evolution of Phage Resistance. Proc. Natl. Acad. Sci. USA 2021, 118, e2104592118. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.A.; Dimitrova, D.; Rather, P.N. Phase-Variable Control of Multiple Phenotypes in Acinetobacter baumannii Strain AB5075. J. Bacteriol. 2015, 197, 2593–2599. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, F.G.; Forsyth, J.H.; Patwa, R.; Kostoulias, X.; Trim, M.; Subedi, D.; Archer, S.K.; Morris, F.C.; Oliveira, C.; Kielty, L.; et al. Bacteriophage-Resistant Acinetobacter baumannii Are Resensitized to Antimicrobials. Nat. Microbiol. 2021, 6, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Regeimbal, J.M.; Jacobs, A.C.; Corey, B.W.; Henry, M.S.; Thompson, M.G.; Pavlicek, R.L.; Quinones, J.; Hannah, R.M.; Ghebremedhin, M.; Crane, N.J.; et al. Personalized Therapeutic Cocktail of Wild Environmental Phages Rescues Mice from Acinetobacter baumannii Wound Infections. Antimicrob. Agents Chemother. 2016, 60, 5806–5816. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.V.; Lavysh, D.G.; Klimuk, E.I.; Edelstein, M.V.; Bogun, A.G.; Shneider, M.M.; Goncharov, A.E.; Leonov, S.V.; Severinov, K.V. Novel Fri1-like Viruses Infecting Acinetobacter baumannii—VB_AbaP_AS11 and VB_AbaP_AS12—Characterization, Comparative Genomic Analysis, and Host-recognition Strategy. Viruses 2017, 9, 188. [Google Scholar] [CrossRef]
- Wang, C.; Li, P.; Zhu, Y.; Huang, Y.; Gao, M.; Yuan, X.; Niu, W.; Liu, H.; Fan, H.; Qin, Y.; et al. Identification of a Novel Acinetobacter baumannii Phage-Derived Depolymerase and Its Therapeutic Application in Mice. Front. Microbiol. 2020, 11, 1407. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liang, L.; Lin, S.; Jia, S. Isolation and Characterization of a Virulent Bacteriophage AB1 of Acinetobacter baumannii. BMC Microbiol. 2010, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Le, S.; Peng, Y.; Zhao, Y.; Yin, S.; Zhang, L.; Yao, X.; Tan, Y.; Li, M.; Hu, F. Characterization and Genome Sequencing of Phage Abp1, a New PhiKMV-Like Virus Infecting Multidrug-Resistant Acinetobacter baumannii. Curr. Microbiol. 2013, 66, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Wozniak, D.J. Phage Cocktail Development for Bacteriophage Therapy: Toward Improving Spectrum of Activity Breadth and Depth. Pharmaceuticals 2021, 14, 1019. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.L.; McCutcheon, J.G.; Stothard, P.; Dennis, J.J. Novel Stenotrophomonas Maltophilia Temperate Phage DLP4 Is Capable of Lysogenic Conversion. BMC Genom. 2019, 20, 300. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.T.; Hanoian, P.; French, J.B.; Pringle, T.H.; Hammes-Schiffer, S.; Benkovic, S.J. Functional Significance of Evolving Protein Sequence in Dihydrofolate Reductase from Bacteria to Humans. Proc. Natl. Acad. Sci. USA 2013, 110, 10159–10164. [Google Scholar] [CrossRef] [PubMed]
- Mosher, R.A.; DiRenzo, A.B.; Mathews, C.K. Bacteriophage T4 Virion Dihydrofolate Reductase: Approaches to Quantitation and Assessment of Function. J. Virol. 1977, 23, 645–658. [Google Scholar] [CrossRef]
- Mosher, R.A.; Mathews, C.K. Bacteriophage T4-Coded Dihydrofolate Reductase: Synthesis, Turnover, and Location of the Virion Protein. J. Virol. 1979, 31, 94–103. [Google Scholar] [CrossRef]
- Bryson, A.L.; Hwang, Y.; Sherrill-Mix, S.; Wu, G.D.; Lewis, J.D.; Black, L.; Clark, T.A.; Bushman, F.D. Covalent Modification of Bacteriophage T4 DNA Inhibits CRISPR-Cas9. mBio 2015, 6, e00648-15. [Google Scholar] [CrossRef]


 ) and DLP2 (
) and DLP2 ( ) on host strain AB5075cm. The mean of biological and technical triplicates represented at each time point and error bars represent standard deviation.
) and DLP2 () on host strain AB5075cm. The mean of biological and technical triplicates represented at each time point and error bars represent standard deviation.
) on host strain AB5075cm. The mean of biological and technical triplicates represented at each time point and error bars represent standard deviation.
) and DLP2 () on host strain AB5075cm. The mean of biological and technical triplicates represented at each time point and error bars represent standard deviation.

 ) and DLP2 (
) and DLP2 ( ) against A. baumannii AB5075cm. The OD600 of kill curves was measured across known MOIs (Figure 4) and virulence curves were generated as described in the methods section. A virulence index of 1 signifies theoretical maximum virulence. The data represent mean and standard deviation of technical and biological triplicate experiments.
) and DLP2 () against A. baumannii AB5075cm. The OD600 of kill curves was measured across known MOIs (Figure 4) and virulence curves were generated as described in the methods section. A virulence index of 1 signifies theoretical maximum virulence. The data represent mean and standard deviation of technical and biological triplicate experiments.
) against A. baumannii AB5075cm. The OD600 of kill curves was measured across known MOIs (Figure 4) and virulence curves were generated as described in the methods section. A virulence index of 1 signifies theoretical maximum virulence. The data represent mean and standard deviation of technical and biological triplicate experiments.
) and DLP2 () against A. baumannii AB5075cm. The OD600 of kill curves was measured across known MOIs (Figure 4) and virulence curves were generated as described in the methods section. A virulence index of 1 signifies theoretical maximum virulence. The data represent mean and standard deviation of technical and biological triplicate experiments.









| Strain | DLP1 (PFU/mL) | DLP2 (PFU/mL) |
|---|---|---|
| AB5075 Δwzc | 2.9 × 108 | 7.4 × 108 |
| AB5075 Δwzc, carO::MarTc | 1.2 × 106 | 0 |
| AB5075 Δwzc, carO::T26 | 1.0 × 106 | 0 |
| AB5075 Δwzc, carO::T26/pQF1266B | 2.4 × 106 | 0 |
| AB5075 Δwzc, carO::T26/pQF1266B + carO | 1.9 × 108 | 2.0 × 107 |
| Database | InterPro ID | InterPro Name |
|---|---|---|
| CDD | IPR001796 | DHFR_dom |
| Gene3D | IPR024072 | DHFR-like_dom_sf |
| Panther | IPR012259 | DHFR |
| Pfam | IPR001796 | DHFR_dom |
| PRINTS | PR00070 | DHFR |
| PROSITE_PROFILES | IPR001796 | DHFR_dom |
| Superfamily | IPR024072 | DHFR-like_dom_sf |
| Database | Hit Description |
|---|---|
| Gene3D | G3DSA:1.10.3730.20 |
| Panther | SMR family proton-dependent drug efflux transporter SUGE |
| Pfam | Multi_Drug_Res |
| Superfamily | Multidrug resistance efflux transporter EmrE |
| TMHMM | unknown |
| PHOBIUS | Transmembrane, cytoplasmic and non-cytoplasmic domains |
| SIGNALP_EUK | SignalP-TM |
| Phage | Genome (bp) | ORFs | tRNAs | GC (%) | Accession | Reference |
|---|---|---|---|---|---|---|
| DLP1 | 164,335 | 248 | 10 | 36.8 | OP946501 | This study |
| DLP2 | 165,122 | 248 | 8 | 36.7 | OP946502 | This study |
| fHyAci03 | 165,975 | 247 | 8 | 36.8 | MH460829 | [68] |
| KARL-1 | 166,560 | 253 | 7 | 36.8 | MH713599 | [69] |
| ZZ1 | 166,687 | 256 | 8 | 34.4 | HQ698922 | [64,70] |
| PhT2 | 166,330 | 255 | 9 | 36.4 | MN864865 | [71] |
| Maestro 1 | 169,176 | 264 | 7 | 36.3 | MT949699 | [12,17] |
| AC4 1 | 168,186 | 250 | 9 | 36.7 | OL770262 | [12,17] |
| AB-Navy1 1 | 166,113 | 247 | 8 | 36.7 | OL770258 | [12,17] |
| AB-Navy4 1 | 166,964 | 246 | 8 | 36.8 | OL770259 | [12,17] |
| AB-Navy71 1 | 166,382 | 252 | 8 | 36.4 | OL770260 | [12,17] |
| AB-Navy97 1 | 165,480 | 244 | 9 | 36.7 | OL770261 | [12,17] |
| ABTZA1 | 168,223 | 253 | 6 | 36.3 | MK278860 | [14] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peters, D.L.; Davis, C.M.; Harris, G.; Zhou, H.; Rather, P.N.; Hrapovic, S.; Lam, E.; Dennis, J.J.; Chen, W. Characterization of Virulent T4-Like Acinetobacter baumannii Bacteriophages DLP1 and DLP2. Viruses 2023, 15, 739. https://doi.org/10.3390/v15030739
Peters DL, Davis CM, Harris G, Zhou H, Rather PN, Hrapovic S, Lam E, Dennis JJ, Chen W. Characterization of Virulent T4-Like Acinetobacter baumannii Bacteriophages DLP1 and DLP2. Viruses. 2023; 15(3):739. https://doi.org/10.3390/v15030739
Chicago/Turabian StylePeters, Danielle L., Carly M. Davis, Greg Harris, Hongyan Zhou, Philip N. Rather, Sabahudin Hrapovic, Edmond Lam, Jonathan J. Dennis, and Wangxue Chen. 2023. "Characterization of Virulent T4-Like Acinetobacter baumannii Bacteriophages DLP1 and DLP2" Viruses 15, no. 3: 739. https://doi.org/10.3390/v15030739
APA StylePeters, D. L., Davis, C. M., Harris, G., Zhou, H., Rather, P. N., Hrapovic, S., Lam, E., Dennis, J. J., & Chen, W. (2023). Characterization of Virulent T4-Like Acinetobacter baumannii Bacteriophages DLP1 and DLP2. Viruses, 15(3), 739. https://doi.org/10.3390/v15030739