
COVID-19 and Pulmonary Angiogenesis: The Possible Role of Hypoxia and Hyperinflammation in the Overexpression of Proteins Involved in Alveolar Vascular Dysfunction
 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approvals
2.2. Samples
2.3. Histopathological and Immunohistochemistry Analysis
2.4. Statistical Analysis
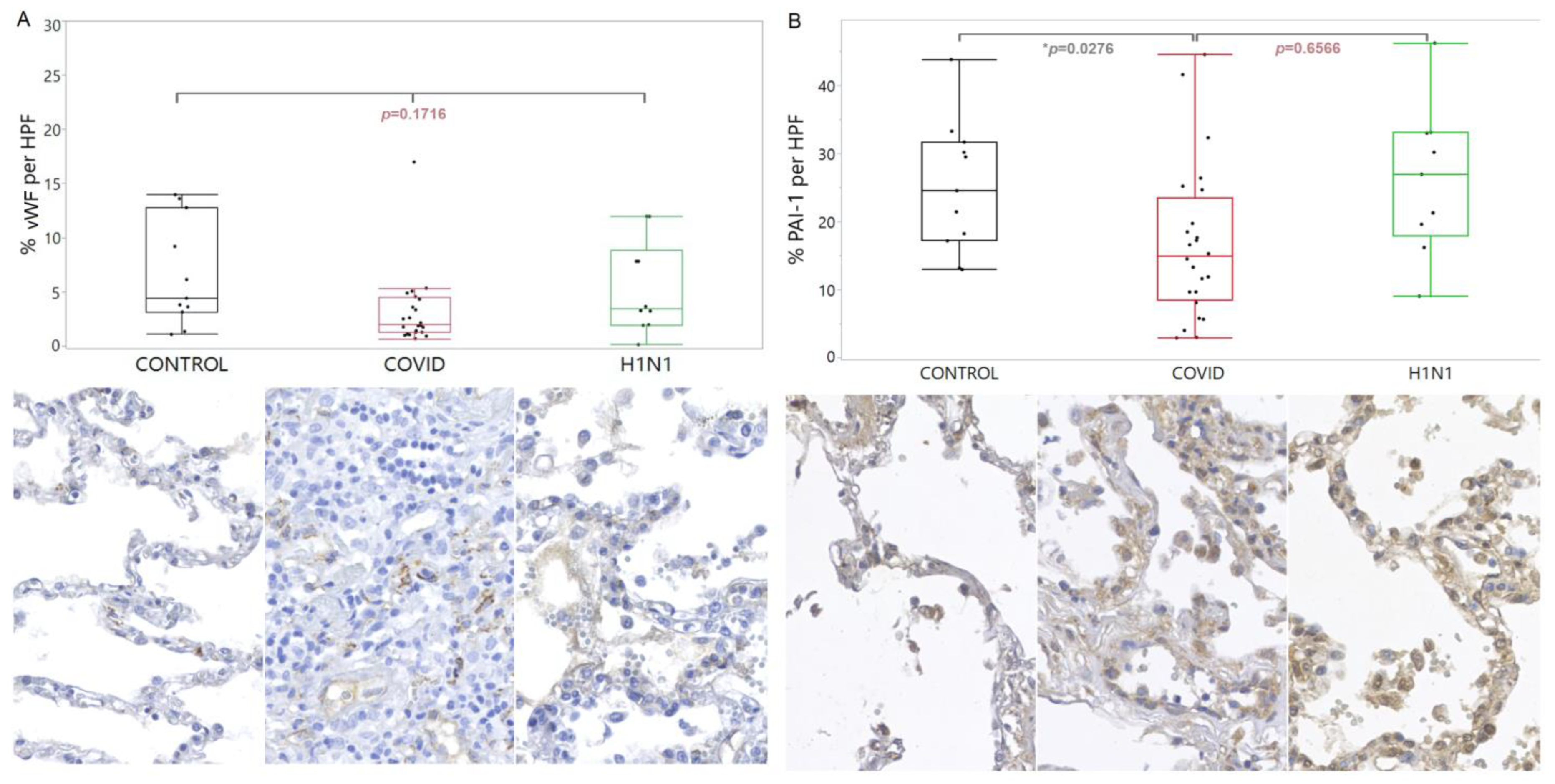
3. Results
4. Discussion
4.1. Anatomopathological and Immunohistochemical Findings
4.2. Clinical Findings
4.3. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beyerstedt, S.; Casaro, E.B.; Rangel, É.B. COVID-19: Angiotensin-converting enzyme 2 (ACE2) expression and tissue susceptibility to SARS-CoV-2 infection. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Yang, X.; Yang, D.; Bao, J.; Li, R.; Xiao, Y.; Hou, C.; Wang, H.; Liu, J.; Yang, D.; et al. Role of angiotensin-converting enzyme 2 (ACE2) in COVID-19. Crit. Care 2020, 24, 422. [Google Scholar] [CrossRef] [PubMed]
- Norooznezhad, A.H.; Mansouri, K. Endothelial cell dysfunction, coagulation, and angiogenesis in coronavirus disease 2019 (COVID-19). Microvasc. Res. 2021, 137, 104188. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, A.; Vecchié, A.; Dagna, L.; Martinod, K.; Dixon, D.L.; Van Tassell, B.W.; Dentali, F.; Montecucco, F.; Massberg, S.; Levi, M.; et al. Endothelial dysfunction and immunothrombosis as key pathogenic mechanisms in COVID-19. Nat. Rev. Immunol. 2021, 21, 319–329. [Google Scholar] [CrossRef]
- Nagashima, S.; Mendes, M.C.; Camargo Martins, A.P.; Borges, N.H.; Godoy, T.M.; Dos Santos Miggiolaro, A.F.R.; Dos Santos Dezidério, F.; Machado-Souza, C.; De Noronha, L. Endothelial Dysfunction and Thrombosis in Patients With COVID-19—Brief Report. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2404–2407. [Google Scholar] [CrossRef]
- Ali, M.A.M.; Spinler, S.A. COVID-19 and thrombosis: From bench to bedside. Trends Cardiovasc. Med. 2020, 31, 143–160. [Google Scholar] [CrossRef]
- Vlahopoulos, S.; Wang, K.; Xue, Y.; Zheng, X.; Boldogh, I.; Pan, L. Endothelial Dysfunction through Oxidatively Generated Epigenetic Mark in Respiratory Viral Infections. Cells 2021, 10, 3067. [Google Scholar] [CrossRef]
- Marchenko, V.A.; Barashkova, S.V.; Zelinskaya, I.A.; Toropova, Y.G.; Ramsay, E.S.; Zhilinskaya, I.N. Modulation of endothelial factors activity in human endothelial cells in influenza A(H1N1)pdm09 virus infection. Probl. Virol. 2021, 66, 198–210. [Google Scholar] [CrossRef]
- Ambrosino, P.; Calcaterra, I.L.; Mosella, M.; Formisano, R.; D’Anna, S.E.; Bachetti, T.; Marcuccio, G.; Galloway, B.; Mancini, F.P.; Papa, A.; et al. Endothelial Dysfunction in COVID-19: A Unifying Mechanism and a Potential Therapeutic Target. Biomedicines 2022, 10, 812. [Google Scholar] [CrossRef]
- Chioh, F.W.J.; Fong, S.W.; Young, B.E.; Wu, K.X.; Siau, A.; Krishnan, S.; Chan, Y.H.; Carissimo, G.; Teo, L.L.Y.; Gao, F.; et al. Convalescent COVID-19 patients are susceptible to endothelial dysfunction due to persistent immune activation. eLife 2021, 10, e64909. [Google Scholar] [CrossRef]
- Ambrosino, P.; Calcaterra, I.; Molino, A.; Moretta, P.; Lupoli, R.; Spedicato, G.; Papa, A.; Motta, A.; Maniscalco, M.; Di Minno, M. Persistent Endothelial Dysfunction in Post-Acute COVID-19 Syndrome: A Case-Control Study. Biomedicines 2021, 9, 957. [Google Scholar] [CrossRef] [PubMed]
- Amraei, R.; Rahimi, N. COVID-19, Renin-Angiotensin System and Endothelial Dysfunction. Cells 2020, 9, 1652. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, L.; Chew, K.Y.; Stocks, C.J.; Yordanov, T.E.; Essebier, P.; Kulasinghe, A.; Monkman, J.; Dos Santos Miggiolaro, A.F.R.; Cooper, C.; de Noronha, L.; et al. Endothelial cells are not productively infected by SARS-CoV-2. Clin. Transl. Immunol. 2021, 10, e1350. [Google Scholar] [CrossRef] [PubMed]
- Daher, J. Endothelial dysfunction and COVID-19 (Review). Biomed. Rep. 2021, 15, 6102. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Narazaki, M.; Metwally, H.; Kishimoto, T. Historical overview of the interleukin-6 family cytokine. J. Exp. Med. 2020, 217, e20190347. [Google Scholar] [CrossRef]
- Melms, J.C.; Biermann, J.; Huang, H.; Wang, Y.; Nair, A.; Tagore, S.; Katsyv, I.; Rendeiro, A.F.; Amin, A.D.; Schapiro, D.; et al. A molecular single-cell lung atlas of lethal COVID-19. Nature 2021, 595, 114–119. [Google Scholar] [CrossRef]
- Mahat, R.K.; Panda, S.; Rathore, V.; Swain, S.; Yadav, L.; Sah, S.P. The dynamics of inflammatory markers in coronavirus disease-2019 (COVID-19) patients: A systematic review and meta-analysis. Clin. Epidemiol. Glob. Health 2021, 11, 100727. [Google Scholar] [CrossRef]
- The WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group; Shankar-Hari, M.; Vale, C.L.; Godolphin, P.J.; Fisher, D.; Higgins, J.P.T.; Spiga, F.; Savović, J.; Tierney, J.; Baron, G.; et al. Association Between Administration of IL-6 Antagonists and Mortality Among Patients Hospitalized for COVID-19: A Meta-analysis. JAMA 2021, 326, 499–518. [Google Scholar] [CrossRef]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef]
- Kaivola, J.; Nyman, T.A.; Matikainen, S. Inflammasomes and SARS-CoV-2 Infection. Viruses 2021, 13, 2513. [Google Scholar] [CrossRef]
- Birnhuber, A.; Fließer, E.; Gorkiewicz, G.; Zacharias, M.; Seeliger, B.; David, S.; Welte, T.; Schmidt, J.; Olschewski, H.; Wygrecka, M.; et al. Between inflammation and thrombosis: Endothelial cells in COVID-19. Eur. Respir. J. 2021, 58, 2100377. [Google Scholar] [CrossRef]
- Smadja, D.M.; Guerin, C.L.; Chocron, R.; Yatim, N.; Boussier, J.; Gendron, N.; Khider, L.; Hadjadj, J.; Goudot, G.; Debuc, B.; et al. Angiopoietin-2 as a marker of endothelial activation is a good predictor factor for intensive care unit admission of COVID-19 patients. Angiogenesis 2020, 23, 611–620. [Google Scholar] [CrossRef]
- Alay, H.; Laloglu, E. The role of angiopoietin-2 and surfactant protein-D levels in SARS-CoV-2-related lung injury: A prospective, observational, cohort study. J. Med. Virol. 2021, 93, 6008–6015. [Google Scholar] [CrossRef]
- Sibila, O.; Perea, L.; Albacar, N.; Moisés, J.; Cruz, T.; Mendoza, N.; Solarat, B.; Lledó, G.; Espinosa, G.; Barberà, J.A.; et al. Elevated plasma levels of epithelial and endothelial cell markers in COVID-19 survivors with reduced lung diffusing capacity six months after hospital discharge. Respir. Res. 2022, 23, 37. [Google Scholar] [CrossRef] [PubMed]
- Price, D.R.; Benedetti, E.; Hoffman, K.L.; Gomez-Escobar, L.; Alvarez-Mulett, S.; Capili, A.; Sarwath, H.; Parkhurst, C.N.; Lafond, E.; Weidman, K.; et al. Angiopoietin 2 Is Associated with Vascular Necroptosis Induction in Coronavirus Disease 2019 Acute Respiratory Distress Syndrome. Am. J. Pathol. 2022, 192, 1001–1015. [Google Scholar] [CrossRef] [PubMed]
- Schmaier, A.A.; Pajares, G.M.; Manickas-Hill, Z.J.; Sack, K.D.; Chen, S.M.; Bhambhani, V.; Quadir, J.; Nath, A.K.; Collier, A.Y.; Ngo, D.; et al. Tie2 activation protects against prothrombotic endothelial dysfunction in COVID-19. J. Clin. Investig. 2021, 6, e151527. [Google Scholar] [CrossRef] [PubMed]
- Smadja, D.M.; Mentzer, S.J.; Fontenay, M.; Laffan, M.A.; Ackermann, M.; Helms, J.; Jonigk, D.; Chocron, R.; Pier, G.B.; Gendron, N.; et al. COVID-19 is a systemic vascular hemopathy: Insight for mechanistic and clinical aspects. Angiogenesis 2021, 24, 755–788. [Google Scholar] [CrossRef]
- Wichmann, D.; Sperhake, J.P.; Lütgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schröder, A.S.; et al. Autopsy Findings and Venous Thromboembolism in Patients with COVID-19. Ann. Intern. Med. 2020, 173, 268–277. [Google Scholar] [CrossRef]
- Menter, T.; Haslbauer, J.D.; Nienhold, R.; Savic, S.; Hopfer, H.; Deigendesch, N.; Frank, S.; Turek, D.; Willi, N.; Pargger, H.; et al. Post-mortem examination of COVID19 patients reveals diffuse alveolar damage with severe capillary congestion and variegated findings of lungs and other organs suggesting vascular dysfunction. Histopathology 2020, 77, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Loo, J.; A Spittle, D.; Newnham, M. COVID-19, immunothrombosis and venous thromboembolism: Biological mechanisms. Thorax 2021, 76, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, Z.M.; Ware, J. von Willebrand factor. FASEB J. 1993, 7, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.A.B.; Maeda, N.Y.; Bydlowski, S.P. Fator von Willebrand e disfunção endotelial pulmonar. Implicações prognósticas. Arq. Bras. De Cardiol. 1998, 70, 141–145. [Google Scholar] [CrossRef]
- Goshua, G.; Pine, A.B.; Meizlish, M.L.; Chang, C.-H.; Zhang, H.; Bahel, P.; Baluha, A.; Bar, N.; Bona, R.D.; Burns, A.J.; et al. Endotheliopathy in COVID-19-associated coagulopathy: Evidence from a single-centre, cross-sectional study. Lancet Haematol. 2020, 7, e575–e582. [Google Scholar] [CrossRef]
- Ladikou, E.E.; Sivaloganathan, H.; Milne, K.M.; Arter, W.E.; Ramasamy, R.; Saad, R.; Stoneham, S.M.; Philips, B.; Eziefula, A.C.; Chevassut, T. Von Willebrand factor (vWF): Marker of endothelial damage and thrombotic risk in COVID-19? Clin. Med. 2020, 20, e178–e182. [Google Scholar] [CrossRef]
- Cesari, M.; Pahor, M.; Incalzi, R.A. REVIEW: Plasminogen Activator Inhibitor-1 (PAI-1): A Key Factor Linking Fibrinolysis and Age-Related Subclinical and Clinical Conditions. Cardiovasc. Ther. 2010, 28, e72–e91. [Google Scholar] [CrossRef]
- Matsuyama, T.; Kubli, S.P.; Yoshinaga, S.K.; Pfeffer, K.; Mak, T.W. An aberrant STAT pathway is central to COVID-19. Cell Death Differ. 2020, 27, 3209–3225. [Google Scholar] [CrossRef]
- D’Alonzo, D.; De Fenza, M.; Pavone, V. COVID-19 and pneumonia: A role for the uPA/uPAR system. Drug Discov. Today 2020, 25, 1528–1534. [Google Scholar] [CrossRef]
- Bahadori, M.; Dabiri, S.; Javadi, A.; Shamsi-Meymandi, S.; Movahedinia, S.; Shamsi-Meymandi, M.; Khorasani, P.; Farrokhnia, M.; Yousefi, M.; Sarrafzadeh, F.; et al. Pathogenesis of COVID-19; Acute Auto-inflammatory Disease (Endotheliopathica & Leukocytoclastica COVIDicus). Arch. Iran. Med. 2021, 24, 419–426. [Google Scholar] [CrossRef]
- Renna, N.F.; de las Heras, N.; Miatello, R.M. Pathophysiology of Vascular Remodeling in Hypertension. Int. J. Hypertens. 2013, 2013, 808353. [Google Scholar] [CrossRef] [PubMed]
- de Cevins, C.; Luka, M.; Smith, N.; Meynier, S.; Magérus, A.; Carbone, F.; García-Paredes, V.; Barnabei, L.; Batignes, M.; Boullé, A.; et al. A monocyte/dendritic cell molecular signature of SARS-CoV-2-related multisystem inflammatory syndrome in children with severe myocarditis. Med 2021, 2, 1072–1092.e7. [Google Scholar] [CrossRef] [PubMed]
- Jahani, M.; Dokaneheifard, S.; Mansouri, K. Hypoxia: A key feature of COVID-19 launching activation of HIF-1 and cytokine storm. J. Inflamm. 2020, 17, 33. [Google Scholar] [CrossRef] [PubMed]
- van Uden, P.; Kenneth, N.S.; Rocha, S. Regulation of hypoxia-inducible factor-1α by NF-κB. Biochem. J. 2008, 412, 477–484. [Google Scholar] [CrossRef]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of CC-Chemokines in the Regulation of Angiogenesis. Int. J. Mol. Sci. 2016, 17, 1856. [Google Scholar] [CrossRef]
- Hashimoto, T.; Shibasaki, F. Hypoxia-Inducible Factor as an Angiogenic Master Switch. Front. Pediatr. 2015, 3, 33. [Google Scholar] [CrossRef]
- Alkharsah, K.R. VEGF Upregulation in Viral Infections and Its Possible Therapeutic Implications. Int. J. Mol. Sci. 2018, 19, 1642. [Google Scholar] [CrossRef]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-Induced Angiogenesis: Good and Evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef]
- Pine, A.B.; Meizlish, M.L.; Goshua, G.; Chang, C.H.; Zhang, H.; Bishai, J.; Bahel, P.; Patel, A.; Gbyli, R.; Kwan, J.M.; et al. Circulating markers of angiogenesis and endotheliopathy in COVID-19. Pulm. Circ. 2020, 10, 2045894020966547. [Google Scholar] [CrossRef]
- Freda, C.T.; Yin, W.; Ghebrehiwet, B.; Rubenstein, D.A. SARS-CoV-2 Structural Proteins Exposure Alter Thrombotic and Inflammatory Responses in Human Endothelial Cells. Cell. Mol. Bioeng. 2021, 15, 43–53. [Google Scholar] [CrossRef]
- Zhu, B.; Wu, Y.; Huang, S.; Zhang, R.; Son, Y.M.; Li, C.; Cheon, I.S.; Gao, X.; Wang, M.; Chen, Y.; et al. Uncoupling of macrophage inflammation from self-renewal modulates host recovery from respiratory viral infection. Immunity 2021, 54, 1200–1218.e9. [Google Scholar] [CrossRef] [PubMed]
- Kulasinghe, A.; Tan, C.W.; Ribeiro Dos Santos Miggiolaro, A.F.; Monkman, J.; SadeghiRad, H.; Bhuva, D.D.; Motta Junior, J.D.S.; Busatta Vaz de Paula, C.; Nagashima, S.; Baena, C.P.; et al. Profiling of lung SARS-CoV-2 and influenza virus infection dissects virus-specific host responses and gene signatures. Eur. Respir. J. 2021, 59, 2101881. [Google Scholar] [CrossRef] [PubMed]
- Landstra, C.P.; de Koning, E.J.P. COVID-19 and Diabetes: Understanding the Interrelationship and Risks for a Severe Course. Front. Endocrinol. 2021, 12, 649525. [Google Scholar] [CrossRef] [PubMed]
- Abani, O.; Abbas, A.; Abbas, F.; Abbas, M.; Abbasi, S.; Abbass, H.; Abbott, A.; Abdallah, N.; Abdelaziz, A.; Abdelfattah, M.; et al. Aspirin in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. Lancet 2022, 399, 143–151. [Google Scholar] [CrossRef]
- Formiga, F.; Rubio-Rivas, M.; Mora-Luján, J.M.; Escudero, S.C.; Martinez, R.F.M.; Mendez-Bailón, M.; Campo, P.D.-D.; Pérez, A.R.; García-Sánchez, F.-J.; Alcalá-Pedrajas, J.N.; et al. Does admission acetylsalicylic acid uptake in hospitalized COVID-19 patients have a protective role? Data from the Spanish SEMI-COVID-19 Registry. Intern. Emerg. Med. 2021, 17, 761–775. [Google Scholar] [CrossRef]
- Martha, J.W.; Pranata, R.; Lim, M.A.; Wibowo, A.; Akbar, M.R. Active prescription of low-dose aspirin during or prior to hospitalization and mortality in COVID-19: A systematic review and meta-analysis of adjusted effect estimates. Int. J. Infect. Dis. 2021, 108, 6–12. [Google Scholar] [CrossRef]
- Aslan, B.; Akyüz, A.; Işık, F.; Çap, M.; Inci, Ü.; Kaya, I.; Karahan, M.Z.; Aktan, A.; Bilge, Ö.; Özbek, M.; et al. The effect of chronic DOAC treatment on clinical outcomes of hospitalized patients with COVID-19. Int. J. Clin. Pract. 2021, 75, e14467. [Google Scholar] [CrossRef]
- Covino, M.; De Matteis, G.; Della Polla, D.; Burzo, M.L.; Pascale, M.M.; Santoro, M.; De Cristofaro, R.; Gasbarrini, A.; De Candia, E.; Franceschi, F. Does chronic oral anticoagulation reduce in-hospital mortality among COVID-19 older patients? Aging Clin. Exp. Res. 2021, 33, 2335–2343. [Google Scholar] [CrossRef]
- Harenberg, J.; Bauersachs, R.; Ageno, W. Does Chronic Treatment with Oral Anticoagulants Ameliorate the Clinical Course of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection in Coronavirus Disease 2019 (COVID-19)? Semin. Thromb. Hemost. 2020, 47, 338–340. [Google Scholar] [CrossRef]
- Sisinni, A.; Rossi, L.; Battista, A.; Poletti, E.; Battista, F.; Battista, R.A.; Malagoli, A.; Biagi, A.; Zanni, A.; Sticozzi, C.; et al. Pre-admission acetylsalicylic acid therapy and impact on in-hospital outcome in COVID-19 patients: The ASA-CARE study. Int. J. Cardiol. 2021, 344, 240–245. [Google Scholar] [CrossRef]
- Tomazini, B.M.; Maia, I.S.; Cavalcanti, A.B.; Berwanger, O.; Rosa, R.G.; Veiga, V.C.; Avezum, A.; Lopes, R.D.; Bueno, F.R.; Silva, M.V.A.O.; et al. Effect of Dexamethasone on Days Alive and Ventilator-Free in Patients With Moderate or Severe Acute Respiratory Distress Syndrome and COVID-19: The CoDEX Randomized Clinical Trial. JAMA 2020, 324, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Isaka, Y.; Hirasawa, Y.; Terada, J.; Shionoya, Y.; Takeshita, Y.; Kinouchi, T.; Koshikawa, K.; Tajima, H.; Kinoshita, T.; Tada, Y.; et al. Preliminary study regarding the predicted body weight-based dexamethasone therapy in patients with COVID-19 pneumonia. Pulm. Pharmacol. Ther. 2021, 72, 102108. [Google Scholar] [CrossRef] [PubMed]
- RECOVERY Collaborative Group; Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; et al. Dexamethasone in Hospitalized Patients with CoVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- Kim, W.-Y.; Kweon, O.J.; Cha, M.J.; Baek, M.S.; Choi, S.-H. Dexamethasone may improve severe COVID-19 via ameliorating endothelial injury and inflammation: A preliminary pilot study. PLoS ONE 2021, 16, e0254167. [Google Scholar] [CrossRef]
- The ARDS Definition of Task Force*; Ranieri, V.M.; Rubenfeld, G.D.; Thompson, B.T.; Ferguson, N.D.; Caldwell, E.; Fan, E.; Camporota, L.; Slutsky, A.S. Acute Respiratory Distress Syndrome: The Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, G.A.; Calvisi, S.L.; DE Lorenzo, R.; DA Prat, V.; Borio, G.; Gallina, G.; Farolfi, F.; Cavallo, L.; Pascali, M.; Castellani, J.; et al. A novel evidence-based algorithm to predict thromboembolism in patients with COVID-19: Preliminary data from a single-centre cohort. Minerva Med. 2022, 113, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Sartini, S.; Massobrio, L.; Cutuli, O.; Campodonico, P.; Bernini, C.; Sartini, M.; Cristina, M.L.; Castellani, L.; Ceschi, L.; Spadaro, M.; et al. Role of SatO2, PaO2/FiO2 Ratio and PaO2 to Predict Adverse Outcome in COVID-19: A Retrospective, Cohort Study. Int. J. Environ. Res. Public Health 2021, 18, 11534. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Data | Variable | CONTROL (n = 11) | COVID-19 (n = 24) | H1N1 (n = 10) | |
|---|---|---|---|---|---|
| Gender 1 | Male | 72.7% (n = 8) | 62.5% (n = 15) | 80% (n = 8) | |
| Female * p-values | 27.3% (n = 3) p = 0.709 | 37.5% (n = 09) | 20% (n = 2) p = 0.437 | ||
| Age (years) 2 | ----- * p-values | 42.3/45 (18–60) p < 0.001 | 71.96/72.5 (46–93) | 43.5/44 (23–61) p < 0.001 | |
| Time from hospitalization to death (days) 2 | ----- * p-values | 7.6/4 (1–46) p < 0.003 | 15.87/13 (1–39) | 4.70/1.5 (1–19) p < 0.003 | |
| Mechanical ventilation (days) 2 | ----- * p-values | ----- ----- | 12.04/9.5 (0–36) | 4.70/1.5 (1–19) p = 0.028 | |
| Histological patterns of DAD | ----- | Normal septum | Interstitial pneumonitis with scarce septal neutrophils with hyaline membrane and micro thrombosis | Interstitial pneumonitis with high septal neutrophils infiltration and no micro thrombosis | |
| Semiquantitative Grading 1 | Micro thrombosis | absence | 91% (n = 10) | 16.67% (n = 4) | 90% (n = 9) |
| mild | 9% (n = 1) | 37.5% (n = 9) | 10% (n = 1) | ||
| moderate | 0% (n = 0) | 33.33% (n = 8) | 0% (n = 0) | ||
| severe | 0% (n = 0) | 12.5% (n = 3) | 0% (n = 0) | ||
| * p-values | p < 0.0001 | p < 0.0001 | |||
Endothelial Damage | absence | 45.45% (n = 5) | 0% (n = 0) | 0% (n = 0) | |
| mild | 54.55% (n = 6) | 4.17% (n = 1) | 60% (n = 6) | ||
| moderate | 0% (n = 0) | 20.83% (n = 5) | 40% (n = 4) | ||
| severe | 0% (n = 0) | 75% (n = 18) | 0% (n = 0) | ||
| * p-values | p < 0.0001 | p < 0.0001 | |||
Vascular Hypertrophy | absence | 100% (n = 11) | 8.34% (n = 2) | 80% (n = 8) | |
| mild | 0% (n = 0) | 33.33% (n = 8) | 10% (n = 1) | ||
| moderate | 0% (n = 0) | 33.33% (n = 8) | 10% (n = 1) | ||
| severe | 0% (n = 0) | 25% (n = 6) | 0% (n = 0) | ||
| * p-values | p < 0.0001 | p < 0.0001 | |||
| COMORBIDITIES | |
| Diabetes Mellitus (Type 2) 1 | 11 (45.8 %) 2 |
| Systemic Arterial Hypertension 1 | 21 (87.5%) 2 |
| Dyslipidemia 1 | 17 (70.8%) 2 |
| Obesity 1 | 6 (25%) 2 |
| Chronic Heart Disease 1 | 11 (45.8%) 2 |
| Chronic Lung Disease 1 | 5 (20.8%) 2 |
| Malignancy 1 | 3 (12.5%) 2 |
| Time from symptom onset to death (days) 3 | 18.6 (07–42) Median = 17 |
| CHRONIC USE MEDICATIONS | |
| Anticoagulant (use chronic) 1 | 4 (16.6%) 2 |
| Antiaggregant Platelets (use chronic) 1 | 15 (62.5%) 2 |
| MECHANICAL VENTILATION DATA | |
| Time of Mechanical Ventilation (MV) 4 | 12.4 (0–29) Median = 11.5 |
| PaO2/FiO2 Ratio (1ºday MV) 4 | 159.9 (0–313) Median = 127.5 |
| LABORATORIES DATA | |
| C-Reactive Protein (mg/dL) 5 | 172.35 (8.7–420) Median = 154.2 |
| D-dimer (µg/L) 5 | 14,784 (425–152,174) Median = 1848 |
| Blood platelets (/mm³) 5 | 197,600 (38,000–366,000) Median = 195,000 |
| MEDICATIONS ADMINISTERED DURING HOSPITAL STAY | |
| Anticoagulant 1 | 24 (100%) 2 |
| Corticosteroids 1 | 17 (70.8%) 2 |
| Oseltamivir 1 | 08 (33.3%) 2 |
| Antibiotic 1 | 24 (100%) 2 |
| Hydroxychloroquine and azithromycin 1 | 5 (20.8%) 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miggiolaro, A.F.R.S.; da Silva, F.P.G.; Wiedmer, D.B.; Godoy, T.M.; Borges, N.H.; Piper, G.W.; Oricil, A.G.G.; Klein, C.K.; Hlatchuk, E.C.; Dagostini, J.C.H.; et al. COVID-19 and Pulmonary Angiogenesis: The Possible Role of Hypoxia and Hyperinflammation in the Overexpression of Proteins Involved in Alveolar Vascular Dysfunction. Viruses 2023, 15, 706. https://doi.org/10.3390/v15030706
Miggiolaro AFRS, da Silva FPG, Wiedmer DB, Godoy TM, Borges NH, Piper GW, Oricil AGG, Klein CK, Hlatchuk EC, Dagostini JCH, et al. COVID-19 and Pulmonary Angiogenesis: The Possible Role of Hypoxia and Hyperinflammation in the Overexpression of Proteins Involved in Alveolar Vascular Dysfunction. Viruses. 2023; 15(3):706. https://doi.org/10.3390/v15030706
Chicago/Turabian StyleMiggiolaro, Anna Flavia Ribeiro Santos, Felipe Paes Gomes da Silva, David Batista Wiedmer, Thiago Mateus Godoy, Nicolas Henrique Borges, Giulia Werner Piper, Alessandro G. G. Oricil, Carolline Konzen Klein, Elisa Carolina Hlatchuk, Júlio César H. Dagostini, and et al. 2023. "COVID-19 and Pulmonary Angiogenesis: The Possible Role of Hypoxia and Hyperinflammation in the Overexpression of Proteins Involved in Alveolar Vascular Dysfunction" Viruses 15, no. 3: 706. https://doi.org/10.3390/v15030706
APA StyleMiggiolaro, A. F. R. S., da Silva, F. P. G., Wiedmer, D. B., Godoy, T. M., Borges, N. H., Piper, G. W., Oricil, A. G. G., Klein, C. K., Hlatchuk, E. C., Dagostini, J. C. H., Collete, M., Arantes, M. P., D’Amico, R. C., Dutra, A. A., de Azevedo, M. L. V., & de Noronha, L. (2023). COVID-19 and Pulmonary Angiogenesis: The Possible Role of Hypoxia and Hyperinflammation in the Overexpression of Proteins Involved in Alveolar Vascular Dysfunction. Viruses, 15(3), 706. https://doi.org/10.3390/v15030706