Abstract
Bunyamwera virus is the prototype of the Bunyamwera serogroup, which belongs to the order Bunyavirales of the Orthobunyavirus genus in the Peribunyaviridae family. Bunyamwera is a negative-sense RNA virus composed of three segments S, M, and L. Genetic recombination is possible between members of this order as it is already documented. Additionally, it can lead to pathogenic or host range improvement, if it occurs with viruses of public health and agricultural importance such as Rift Valley fever virus and Crimea–Congo hemorrhagic fever virus. Here, we characterize five African Orthobunyavirus viruses from different geographical regions. Our results suggest that the five newly characterized strains are identified as Bunyamwera virus strains. Furthermore, two of the five strains sequenced in this study are recombinant strains, as fragments of their segments are carried by Ngari and Bunyamwera strains. Further investigations are needed to understand the functional impact of these recombinations.
1. Introduction
The Bunyavirales are one of the largest viral orders, containing more than 350 viruses including several viruses of public health and agricultural importance such as Rift Valley fever virus (RVF) and Crimean–Congo hemorrhagic fever virus [1]. As Bunyavirales members, Orthobunyaviruses include a heterogeneous viral group, which Bunyamwera serogroup is one of the largest. The Bunyamwera serogroup regroups arthropod-borne virus members, Bunyamwera virus (BUNV), Ngari virus (NRIV), and Batai virus (BATV), are some of the most isolated members of this serogroup in Africa [2].
BUNV, BATV, and NRIV have a tri-segmented RNA genome: small (S), medium (M), and large (L). The S segment encodes the nucleocapsid protein (N) and a non-structural protein (NSs), which are translated from overlapping open reading frames (ORF). The M encodes glycoproteins (Gn and Gc) and a non-structural protein (NSm), and the L mRNA encodes the RNA-dependent RNA polymerase (RDRP) [3].
Bunyamwera virus (BUNV) was first isolated in Uganda in 1943. Since that, BUNV was isolated from many arthropods such as Aedes mosquitoes and Amblyomma and Hyalomma ticks [4,5]. BUNV is widespread in several African countries, North America, and South America. To note, the virus was reported in Uganda, Tanzania, Mozambique, Nigeria, Guinea, South Africa, the Democratic Republic of Congo, Senegal, Guinea, the Ivory Coast, Nigeria, Cameroon, the Central African Republic, Kenya, Uganda, South Africa, Madagascar, Argentina, and the United States of America [4,6,7]. BUNV causes disease in both humans and domestic animals. The disease associated with BUNV has been reported to cause mild symptoms, such as fever, joint pain, and rash, in many mammals, including humans [4]. In 2015, BUNV infection was associated with encephalitis/neurological disease and abortion in three horses in Argentina [6].
Firstly, isolated from Anopheles maculipennis in Czechoslovakia in 1950, the Batai virus (BATV) is nowadays the Bunyamwera serogroup member with the largest geographical distribution. BATV was isolated in Asia (India and Malaysia), Europe (Ukraine, Croatia, Norway, Montenegro, Slovakia, Belarus, the Czech Republic, Serbia, Bosnia, Italy, Hungary, Romania, Austria, Portugal, Germany, Finland, and Sweden), and Africa (Uganda) [2]. In livestock animals, BATV can cause severe symptoms including inborn deficiency and abortion [8], and in humans, flu-like symptoms are frequently associated with BATV infection [9].
Genetic reassortment and genetic recombination are the driving forces in the evolution of viruses from the Bunyamwera serogroup [10,11]. Recombination events had already been demonstrated between the snowshoe hare virus and La Crosse virus in co-infected cell culture and also in co-infected arthropod hosts [11]. Additionally, natural reassortment had been documented between four orthobunyavirus members: between Bunyamwera virus and Batai virus, and between Potosi virus and Main Drain virus [12,13,14,15,16]. For instance, Ngari (NRIV) possesses the M segment of the Batai virus (BATV) combined with the S and L segments from BUNV [15]. The current demarcation criteria for interspecific differentiation established for bunyaviruses, BUNV, and NRIV belong to Bunyamwera orthobunyavirus spp. [16]. They are two different genetic variants of the same species—Bunyamwera orthobunyavirus sp. [16]. As the Batai virus is known to cause more severe symptoms than the Bunyamwera virus, the reassortment event likely affected the emergence of NRIV, making it more virulent and capable of inducing severe and fatal hemorrhagic fever with a similarity of its clinical manifestations with RVF in humans and small ruminants [2,17,18]. For the record, NRIV is frequently isolated during Rift Valley fever virus (RVF) outbreaks, which could be due to the similarity of their symptoms or signs [14,19].
A recent study shows that this segment reassortment between Batai and Bunyamwera probably occurred in co-infected mammalian cells [20] and that NRIV is only reported in Africa (Senegal, Burkina Faso, Mauritania, the Central African Republic, Madagascar, Kenya, and Somalia) [14,15,16,17,18,19,20,21]. These data combined with the fact that many of the known orthobunyavirus members use the same hosts and are located in the same area showed enormous potential for the emergence of reassortment and recombinant bunyaviruses with pathogenic potential improvement or host range [18].
In Senegal, few are known about Orthobunyavirus, so a genetic characterization of orthobunyavirus isolates from the Institut Pasteur de Dakar (IPD) biobank using Next Generation Sequencing (NGS) was aimed in this study.
2. Materials and Methods
2.1. Viral Stock Preparation
The viral strains used in this study were provided by the WHO collaborating center for reference and research for arboviruses and hemorrhagic fever viruses (WHOCC) in the Virology Department of the Institut Pasteur de Dakar (IPD). Those strains are described in Table 1. The viral stock was prepared by inoculating 200 µL of each viral strain into Vero cells monolayer continuous cell lines (ATCC, USA) in Leibovitz 15 (L-15) growth medium (GibcoBRL, Grand Island, NY, USA) supplemented with 5% fetal bovine serum (FBS) (GibcoBRL, Grand Island, NY, USA), 1% penicillin-streptomycin, and 0.05% fungizone (Sigma, Germany). After the observation of the cytopathic effect, supernatants were harvested and stored at −80 °C until further use.

Table 1.
Strains used in this study.
2.2. Conventional RT-PCR
RNA was extracted from supernatants using the QIAamp RNA Viral Kit (Qiagen GmbH, Heiden, Germany) following the manufacturer’s recommendations. Using the RNA extracts, we tested the presence of the Orthobunyavirus S segment by RT-PCR using TITAN one-tube RT-PCR kit (Boehringer Mannheim Biochemicals) reagents and the primers previously described [22]. For the reaction mixture, we added 4 μL of RNA with the mixture reaction composed of 10.5 μL H2O, 5 μL of buffer, 2.5 μL of each primer, and 0.5 μL of the enzyme. The PCR was performed following the manufacturer’s recommendations. PCR products were visualized on 2% ethidium bromide-stained agarose gel.
2.3. Next-Generation Sequencing
RNA extract was used for Illumina sequencing. Briefly, the host’s background RNA was depleted as previously described using mammal ribosomal RNA-specific primers and RNAseH enzyme (NEB). Purified viral RNA was reverse transcribed into first-stranded cDNA using the Invitrogen SuperScript IV Reverse Transcriptase kit (Thermo Fisher, Waltham, MA, USA). Then, amplicons were tagged using the “index tags” incorporated into the Nextera kit XT DNA Library Preparation kit (Illumina, San Diego, CA, USA). The obtained libraries were quantified, normalized, pooled, and loaded onto an Illumina MiSeq platform with 151 bp paired-end reads using the Miseq reagents kit v3, according to the manufacturer’s protocol (Illumina, San Diego, CA, USA). Demultiplexing, removal of sequencing adapters, and genome mapping were performed by a homemade package incorporated into the IPD server. Genetic identity was determined by a nucleotide Basic Local Alignment Search Tool (nBLAST).
2.4. Sequence Analysis
The MUSCLE alignment algorithm was used for multiple nucleotides sequences alignments in VIPR software [23] using nucleotide sequences classified as Bunyamwera, Batai, Kairi, and Ngari [16] (Table A1). Recombination was tested with Recombination Detection Program (RDP) version 4 (RDP4), a software package suitable for statistical identification and characterization of recombination events in nucleotide sequences [24]. For this purpose, the 3 segments were linearized and aligned. Then, the seven primary exploratory recombination signal detection methods provided by the package (the original RDP method, GENECONV, BOOTSCAN/RECSCAN, MAXCHI, Chimaera, 3SEQ, and SISCAN) were used for the evaluation of recombination events using defaults parameters. Analysis by pairwise comparison of L-segment’s full-length amino acid was performed to check if our strains belonged to the Bunyamwera group according to the current ICTV demarcation criteria [25]. Furthermore, the genetic distance between population was calculated to measure the genetic divergence between groups of sequences, and fixation index (FST) values were estimated to estimate the genetic differentiation of the tested viruses using MEGA [25] and DNAsp [26] software, respectively. Genetic distance and FST values vary between 0 and 1. That means that populations with a value of 0 are genetically similar whereas a value of 1 indicates that these populations are genetically divergent. Maximum likelihood (ML) tree constructions were performed using IQ-TREE software [27]. The appropriate model for each segment was also calculated and the robustness of the tree topology was tested during 1000 non-parametric bootstrap analyses. Kairi virus was used as an outgroup. For the record, genetic recombination is an evolutionary process that results in an exchange of genetic information between segments. Recombination takes place between viruses of the same group or subgroup, whereas, reassortment takes place between viruses of different groups. Reassortment is a non-classical mechanism of recombination that does not involve breaking the nucleic acid of the viral genome.
3. Results
To fulfill the gap of the lack of data about the genetic characterization of the Bunyamwera viruses isolated in Senegal, we aim to genetically characterize Orthobunyavirus strains that are available in the WHO collaborating center for arboviruses and viral hemorrhagic fever (WHOCC) located in IPD.
After sequencing, five complete genomes were obtained and nucleotide BLAST showed that the segments of the ArB218, ArN31, ArMsam263, ArY380, and ArY52 strains were identified as Bunyamwera viruses with high nucleotide similarities (>95%). Except for the strain ArB218 which showed 98.12% of nucleotide identity for the segment S of NRIV strain 9800535 (JX857328.1) for the S segment. An analysis of the genetic distances shows that our isolates are closer to Bunyamwera than Ngari and Batai (Table 2).
Table 2.
Estimates of net genetic distance between groups.
The fixation indices (FST) measured between populations also show that these newly sequenced strains are closer to the Bunyamwera virus (Table 3).
Table 3.
Estimates of FST between groups.
Analyses of the amino acid similarities of the L segments between our strains and the sampled NRIV, BATV, and BUNV sequences reveal 98.82% of similarities with BUNV (Table 4).
Table 4.
Estimates of L segments amino acid similarities between groups.
The recombination test identified two recombinants among our five strains and the breakpoint was detected by some of the seven primary exploratory recombination signal detection methods provided by the package. Recombination analysis shows that the S and L of the Ngari virus strains (15Guidimaka_ME, 428Trarza_ME, 9800521, D28542_4e, HKV66, HKV141, and Adrar) result from recombination with the S and L segments of the newly characterized strain ArN31, with a recombination signal detected by all the seven detections methods (p-value < 0.001) (Figure 1). Moreover, six methods (RDP, BOOTSCAN/RECSCAN, MAXCHI, Chimaera, 3SEQ, and SISCAN) indicate that the segments S and L from the Bunyamwera virus strain ATCC_R_VR_87 and Bunyamwera virus (reference strain) carried fragments from S, M, and L of the ArB218 strains (p-value < 0.001) (Figure 1).

Figure 1.
Representation of recombination events between our strains and the sampled sequences. *: Bunyamwera virus (reference strain). **: Ngari virus strains (15Guidimaka_ME, 428Trarza_ME, 9800521, D28542_4e, HKV66, HKV141, and Adrar).
Phylogenetic analysis shows that all the strains characterized in this study had clustered with Bunyamwera virus strains for all segments, except for the S and L segments of ArB218, which had clustered in the base of the node of the S and L segments of the NRIV strains group (Figure 2A–C) with the two recombinants Bunyamwera virus strains, KW_S1_25397 and 84 Brakna ME, respectively, for the S and L segments. More particularly, the phylogenetic analyses of the four other strains of this study show that they are grouped by two by country (Ar Ms am 263_Kenya with Ar Na 31_Kenya and Ar YM 52_Cameroon with Ar Y 380/69_Cameroon) for the S and M segments. For the L segment, Ar Ms am 263 and Ar Na 31 remain grouped, whereas Ar YM 52 and Ar Y 380/69 were no longer grouped by two for the L segments. These strains from Cameroon seem to be close to some Bunyamwera strains from Kenya.
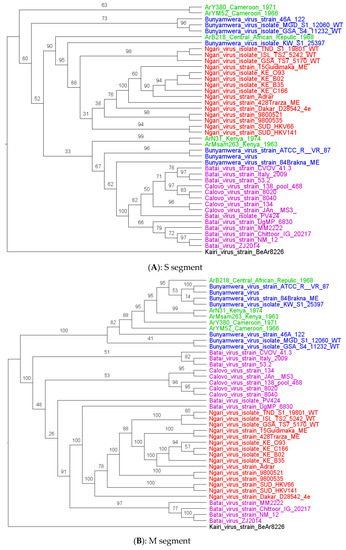
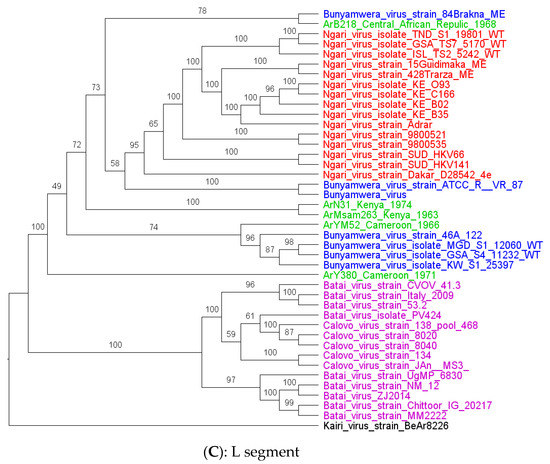
Figure 2.
Maximum likelihood trees were obtained from the S (A), M (B), and L (C) segments analysis. Legends: study strain (green), Batai virus (pink), Bunyamwera (red), and Ngari (blue). The strain characterized in this study is colored purple.
4. Discussion
In this study, we have genetically characterized five Bunyamwera strains provided by the WHOCC in IPD. The results of BLAST show that our five orthobunyavirus strains were identified as belonging to the Bunyamwera serogroup as all their three segments are identified as a Bunyamwera virus (BUNV) except for the S Segment of ArB 218, which was identified as an Ngari virus (NRIV). This result is strengthened by an analysis of the genetic distance, amino acid similarities of the L segments, and the differentiation index (FST), which indicate that these newly sequenced strains are the Bunyamwera virus. Otherwise, two recombination events were detected during our analysis. Ngari strains (15Guidimaka_ME, 428Trarza_ME, Ngari virus strain 9800535, 9800521, D28542_4e, HKV66, HKV141, and Adrar) result from recombination with S and L segments of our strain ArN31. Additionally, Bunyamwera virus strain ATCC_R_VR_87 results from recombination with S, M, and L segments of the ArB218 strain. Natural genetic segments exchange between Bunyamwera serogroup members has already occurred. Genomic data showed that NRIV carried the M segment of BATV and the L and S segments of BUNV [15,18]. According to the current knowledge, BUNV and NRIV share the same geographic areas and host (arthropod and vertebrate) [2]. Then, this segment recombination between our strains ArN31 and ArB218 with Ngari strain SUD-HKV141, Ngari virus strain 9800535, and Bunyamwera virus strain ATCC_R_VR_87 could have occurred in co-infected vectors or in the infected vertebrate hosts [12,28]. This result was strengthened by a phylogenetic analysis where the S and the L segment of ArB 218 clustered on the basal node with S and L segments of the NRIV strains group when the M segment clustered with M of BUNV groups. As the S segment encodes for the NSs, which is known to block the production of type I IFN, it blocks transcription and translation, induces apoptosis, and inhibits apoptosis. This S-segment recombination could lead to the emergence of a new virus with a greater or lesser potential to constitute a threat to humans and livestock, as already documented with NRIV [15,29].
Furthermore, among the strains (Ar Ms am 263, Ar Na 31, Ar YM 52, and Ar Y 380/69) that fall in the Bunyamwera genetic cluster for all the three segments, the newly sequenced Ar Ms am 263 and Ar Na 31 from Kenya were grouped for the S, M, and L segments in a BUNV cluster, even if they were isolated 10 years apart. This could be explained by the fact that they were isolated from hosts that belong to the same genus. Besides the strains from Cameroon, Ar YM 52 and Ar Y 380/69 did not group by two except for the S segments. Additionally, the L segment is close to some of the Bunyamwera isolates from Kenya GSA/S4/11232_WT, MGD/S1/12060_WT, and 46A-122.
Finally, the genetic analyses of five newly characterized strains revealed that these strains are characterized by the Bunyamwera virus and that two of them are recombined with the Ngari and Bunyamwera strains. Further investigations are needed to elucidate the impact of these recombinations on the biology of this virus, such as growth kinetics, pathogenesis analysis, and vector competence, which are needed to understand the role of this genetic recombination.
Author Contributions
Conceptualization, O.F. (Oumar Faye) and V.G.D.; methodology, M.M., I.D., C.T.T., M.N., M.M.D; A.S.G., A.S.D., O.F. (Oumar Faye) and V.G.D.; software, M.M., I.D., C.T.T., M.N., M.M.D., A.S.G. and A.S.D.; validation, O.F. (Ousmane Faye), A.A.S., O.F. (Oumar Faye) and V.G.D.; formal analysis, M.M., I.D., C.T.T., M.N., M.M.D., A.S.G. and A.S.D.; investigation, Not applicable., data curation, O.F. (Ousmane Faye), A.A.S., O.F. (Oumar Faye) and V.G.D.; writing—original draft preparation, M.M., I.D., C.T.T., M.N., M.M.D., A.S.G., A.S.D., O.F. (Oumar Faye) and V.G.D.; writing—review and editing, M.M., I.D., C.T.T., M.N., M.M.D., A.S.G., A.S.D., O.F. (Ousmane Faye), A.A.S., B.F., O.F. (Oumar Faye) and V.G.D.; visualization, M.M., I.D., C.T.T., M.N., M.M.D., A.S.G., A.S.D.; supervision, O.F. (Ousmane Faye)., A.A.S., O.F. (Oumar Faye) and V.G.D.; project administration, O.F. (Ousmane Faye), A.A.S., O.F. (Oumar Faye) and V.G.D.; funding acquisition, O.F. (Oumar Faye) and V.G.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the RSF grant no. 20-64-46014 and IPD proper fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge the colleagues of IPD WHOCC who hosted in the Virology Department, for sharing virus strains and supportive information that was necessary for the establishment and accomplishment of this study.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Table A1.
Viruses used in this study.
Table A1.
Viruses used in this study.
| Virus Name | Virus Strains |
|---|---|
| Batai | Batai virus strain Chittoor/IG-20217 |
| Batai virus strain CVOV 41.3 | |
| Batai virus strain Italy-2009 | |
| Batai virus strain MM2222 | |
| Batai virus strain NM/12 | |
| Batai virus isolate PV424 | |
| Batai virus strain UgMP-6830 | |
| Batai virus ZJ2014 Batai_virus_strain_53.2 | |
| Calovo virus strain 134 | |
| Calovo virus strain 138-pool 468 | |
| Calovo virus strain 8020 | |
| Calovo virus strain 8040 | |
| Calovo virus strain Jan (MS3) | |
| Kairi | Kairi virus strain BeAr8226 |
| Bunyamwera | Bunyamwera virus strain 46A-122 |
| Bunyamwera virus Bunyamwera virus isolate MGD_S1_12060_WT Bunyamwera virus strain ATCC(R) VR-87 Bunyamwera virus isolate GSA_S4_11232_WT Bunyamwera virus strain 84Brakna ME Bunyamwera_virus_isolate_KW_S1_25397 | |
| Newly characterized strains | ArB218_Central African Repulic_1968 ArN31_Kenya_1974 ArMsam263_Kenya_1963 ArY380_Cameroon_1971 ArY52_Cameroon_1966 |
| Ngari | Ngari virus strain 9800521 Ngari virus strain 9800535 Ngari virus strain Adrar Ngari virus strain Dakar D28542/4e Ngari virus strain SUD-HKV66 Ngari virus strain SUD-HKV141 Ngari_virus_isolate_KE_B35 Ngari_virus_isolate_KE_B02 Ngari_virus_isolate_KE_O93 Ngari_virus_isolate_KE_C166 Ngari virus strain 15Guidimaka ME Ngari virus strain 428Trarza ME Ngari virus strain 428Trarza ME Ngari_virus_isolate_ISL_TS2_5242_WT Ngari_virus_isolate_GSA_TS7_5170_WT Ngari_virus_isolate_TND_S1_19801_WT |
References
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, É.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef] [PubMed]
- Dutuze, M.F.; Nzayirambaho, M.; Mores, C.N.; Christofferson, R.C. A Review of Bunyamwera, Batai, and Ngari Viruses: Understudied Orthobunyaviruses with Potential One Health Implications. Front. Vet. Sci. 2018, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.M. Orthobunyaviruses: Recent genetic and structural insights. Nat. Rev. Microbiol. 2014, 12, 673–685. [Google Scholar] [CrossRef]
- Kokernot, R.H.; Smithburn, K.C.; De Meillon, B.; Paterson, H.E. Isolation of Bunyamwera Virus from a Naturally Infected Human Being and Further Isolations from Aedes (Banksinella) Circumluteolus Theo. 1. Am. J. Trop. Med. Hyg. 1958, 7, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Lwande, O.W.; Lutomiah, J.; Obanda, V.; Gakuya, F.; Mutisya, J.; Mulwa, F.; Michuki, G.; Chepkorir, E.; Fischer, A.; Venter, M.; et al. Isolation of Tick and Mosquito-Borne Arboviruses from Ticks Sampled from Livestock and Wild Animal Hosts in Ijara District, Kenya. Vector-Borne Zoonotic Dis. 2013, 13, 637–642. [Google Scholar] [CrossRef]
- Tauro, L.B.; Rivarola, M.E.; Lucca, E.; Mariño, B.; Mazzini, R.; Cardoso, J.F.; Barrandeguy, M.E.; Nunes, M.R.T.; Contigiani, M.S. First isolation of Bunyamwera virus (Bunyaviridae family) from horses with neurological disease and an abortion in Argentina. Vet. J. 2015, 206, 111–114. [Google Scholar] [CrossRef]
- Wertheim, H.F.; Horby, P.; Woodall, J.P. (Eds.) Atlas of Human Infectious Diseases; John Wiley & Sons: Chichester, UK, 2012. [Google Scholar]
- Hoffmann, A.R.; Dorniak, P.; Filant, J.; Dunlap, K.A.; Bazer, F.W.; de la Concha-Bermejillo, A.; Welsh, C.J.; Varner, P.; Edwards, J.F. Ovine Fetal Immune Response to Cache Valley Virus Infection. J. Virol. 2013, 87, 5586–5592. [Google Scholar] [CrossRef]
- Lozach, P.-Y.; Mancini, R.; Bitto, D.; Meier, R.; Oestereich, L.; Överby, A.K.; Pettersson, R.F.; Helenius, A. Entry of Bunyaviruses into Mammalian Cells. Cell Host Microbe 2010, 7, 488–499. [Google Scholar] [CrossRef]
- Holland, J.; Domingo, E. Origin and Evolution of Viruses. Virus Genes 1998, 16, 13–21. [Google Scholar] [CrossRef]
- Gentsch, J.; Wynne, L.R.; Clewley, J.P.; Shope, R.E.; Bishop, D.H. Formation of recombinants between snowshoe hare and La Crosse bunyaviruses. J. Virol. 1977, 24, 893–902. [Google Scholar] [CrossRef]
- Borucki, M.K.; Chandler, L.J.; Parker, B.M.; Blair, C.D.; Beaty, B.J. Bunyavirus superinfection and segment reassortment in transovarially infected mosquitoes. J. Gen. Virol. 1999, 80 Pt 12, 3173–3179. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Saiyasombat, R.; Dorman, K.S.; Garcia-Rejon, J.E.; Farfan-Ale, J.A.; Loroño-Pino, M.A. Sequence and phylogenetic data indicate that an orthobunyavirus recently detected in the Yucatan Peninsula of Mexico is a novel reassortant of Potosi and Cache Valley viruses. Arch. Virol. 2012, 157, 1199–1204. [Google Scholar] [CrossRef]
- Briese, T.; Kapoor, V.; Lipkin, W.I. Natural M-segment reassortment in Potosi and Main Drain viruses: Implications for the evolution of orthobunyaviruses. Arch. Virol. 2007, 152, 2237–2247. [Google Scholar] [CrossRef]
- Briese, T.; Bird, B.; Kapoor, V.; Nichol, S.T.; Lipkin, W.I. Batai and Ngari Viruses: M Segment Reassortment and Association with Severe Febrile Disease Outbreaks in East Africa. J. Virol. 2006, 80, 5627–5630. [Google Scholar] [CrossRef]
- Dolgova, A.S.; Safonova, M.V.; Faye, O.; Dedkov, V.G. Current View on Genetic Relationships within the Bunyamwera Serological Group. Viruses 2022, 14, 1135. [Google Scholar] [CrossRef]
- Omoga, D.C.A.; Tchouassi, D.P.; Venter, M.; Ogola, E.O.; Eibner, G.J.; Kopp, A.; Slothouwer, I.; Torto, B.; Junglen, S.; Sang, R. Circulation of Ngari Virus in Livestock, Kenya. Msphere 2022, 7, e0041622. [Google Scholar] [CrossRef]
- Gerrard, S.R.; Li, L.; Barrett, A.D.; Nichol, S.T. Ngari Virus Is a Bunyamwera Virus Reassortant That Can Be Associated with Large Outbreaks of Hemorrhagic Fever in Africa. J. Virol. 2004, 78, 8922–8926. [Google Scholar] [CrossRef]
- Eiden, M.; Vina-Rodriguez, A.; El Mamy, B.O.; Isselmou, K.; Ziegler, U.; Höper, D.; Jäckel, S.; Balkema-Buschmann, A.; Unger, H.; Doumbia, B.; et al. Ngari Virus in Goats during Rift Valley Fever Outbreak, Mauritania, 2010. Emerg. Infect. Dis. 2014, 20, 2174–2176. [Google Scholar] [CrossRef]
- Heitmann, A.; Gusmag, F.; Rathjens, M.G.; Maurer, M.; Frankze, K.; Schicht, S.; Jansen, S.; Schmidt-Chanasit, J.; Jung, K.; Becker, S.C. Mammals Preferred: Reassortment of Batai and Bunyamwera orthobunyavirus Occurs in Mammalian but Not Insect Cells. Viruses 2021, 13, 1702. [Google Scholar] [CrossRef] [PubMed]
- Makenov, M.T.; Toure, A.H.; Bayandin, R.B.; Gladysheva, A.V.; Shipovalov, A.V.; Boumbaly, S.; Sacko, N.; Korneev, M.G.; Yakovlev, S.A.; Zhurenkova, O.B.; et al. Ngari virus (Orthobunyavirus, Peribunyaviridae) in ixodid ticks collected from cattle in Guinea. Acta Trop. 2021, 214, 105790. [Google Scholar] [CrossRef]
- Yandoko, E.N.; Gribaldo, S.; Finance, C.; Le Faou, A.; Rihn, B.H. Molecular characterization of African orthobunyaviruses. J. Gen. Virol. 2007, 88, 1761–1766. [Google Scholar] [CrossRef] [PubMed]
- Pickett, B.E.; Sadat, E.L.; Zhang, Y.; Noronha, J.M.; Squires, R.B.; Hunt, V.; Liu, M.; Kumar, S.; Zaremba, S.; Gu, Z.; et al. ViPR: An open bioinformatics database and analysis resource for virology research. Nucleic Acids Res. 2012, 40, D593–D598. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Gentsch, J.; Bishop, D.H.L. Recombination and Complementation Between Temperature-Sensitive Mutants of a Bunyavirus, Snowshoe Hare Virus. J. Virol. 1976, 20, 351–354. [Google Scholar] [CrossRef]
- Bowen, M.D.; Trappier, S.G.; Sanchez, A.J.; Meyer, R.F.; Goldsmith, C.S.; Zaki, S.R.; Dunster, L.M.; Peters, C.; Ksiazek, T.G.; Nichol, S.T. A Reassortant Bunyavirus Isolated from Acute Hemorrhagic Fever Cases in Kenya and Somalia. Virology 2001, 291, 185–190. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).