
Altered Ex Vivo NLRP3 Inflammasome Activation Is Associated with 28-Day Mortality in Septic Patients

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Reagents
2.3. NLRP3 Activation Monitoring in Whole Blood Myeloid Cells
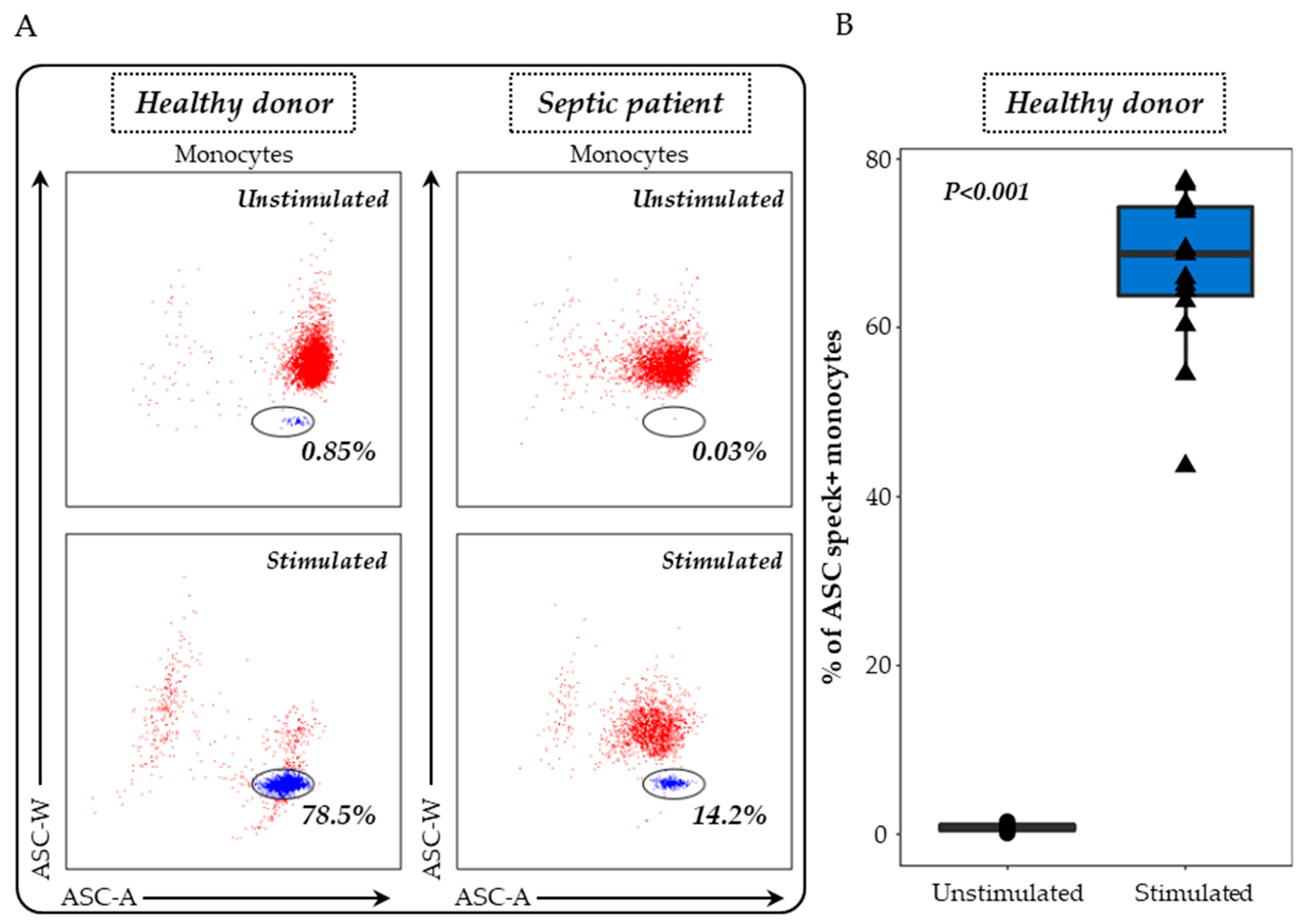
2.3.1. Monocytes ASC Staining in Whole Blood
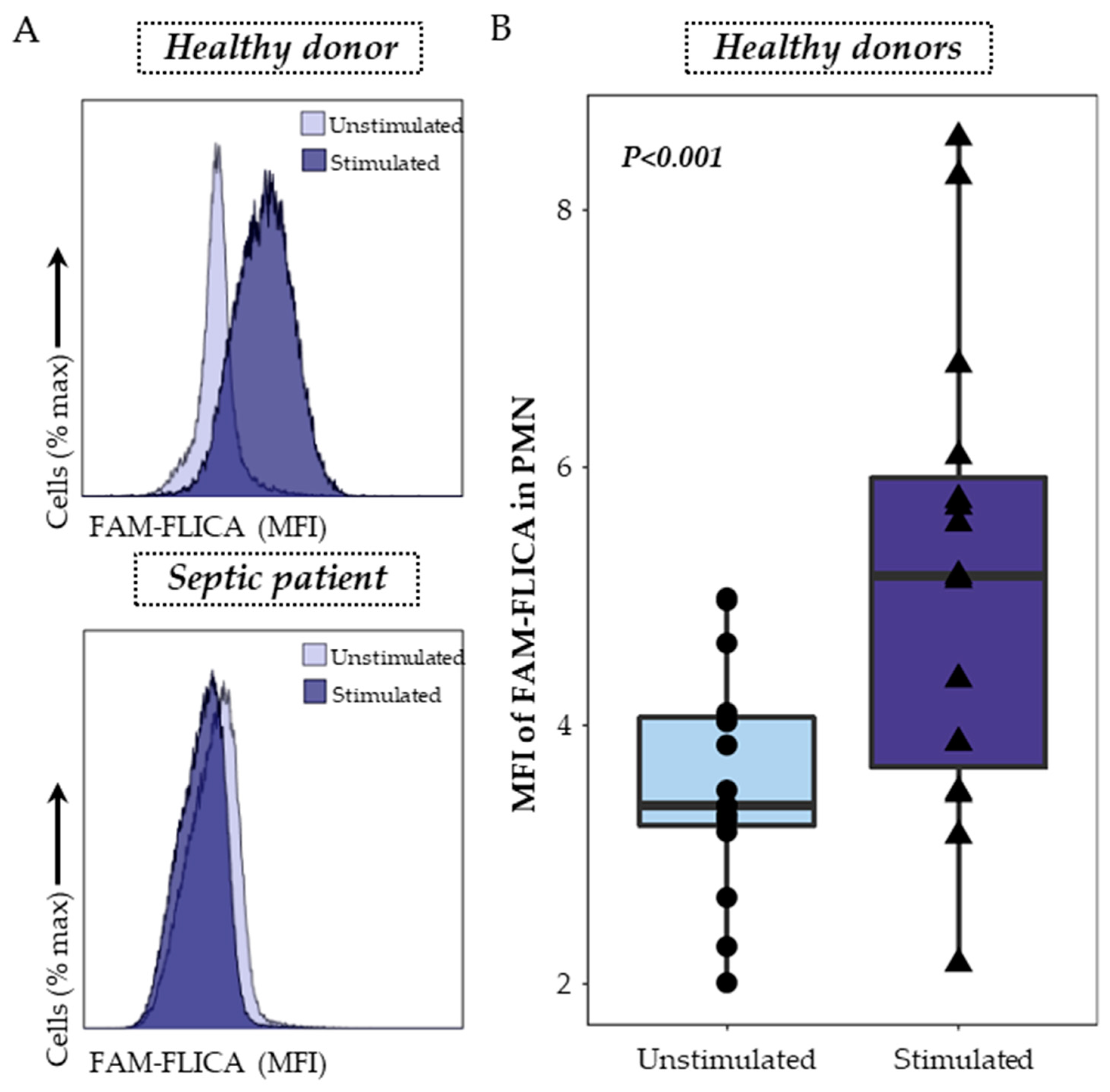
2.3.2. Caspase-1 Staining in PMN
2.4. Statistical Analysis
3. Results
3.1. Patients Characteristics
3.2. Detection of ASC Specks in Monocytes and Activated Caspase-1 in PMN as Indicators of NLRP3 Inflammasome Activation
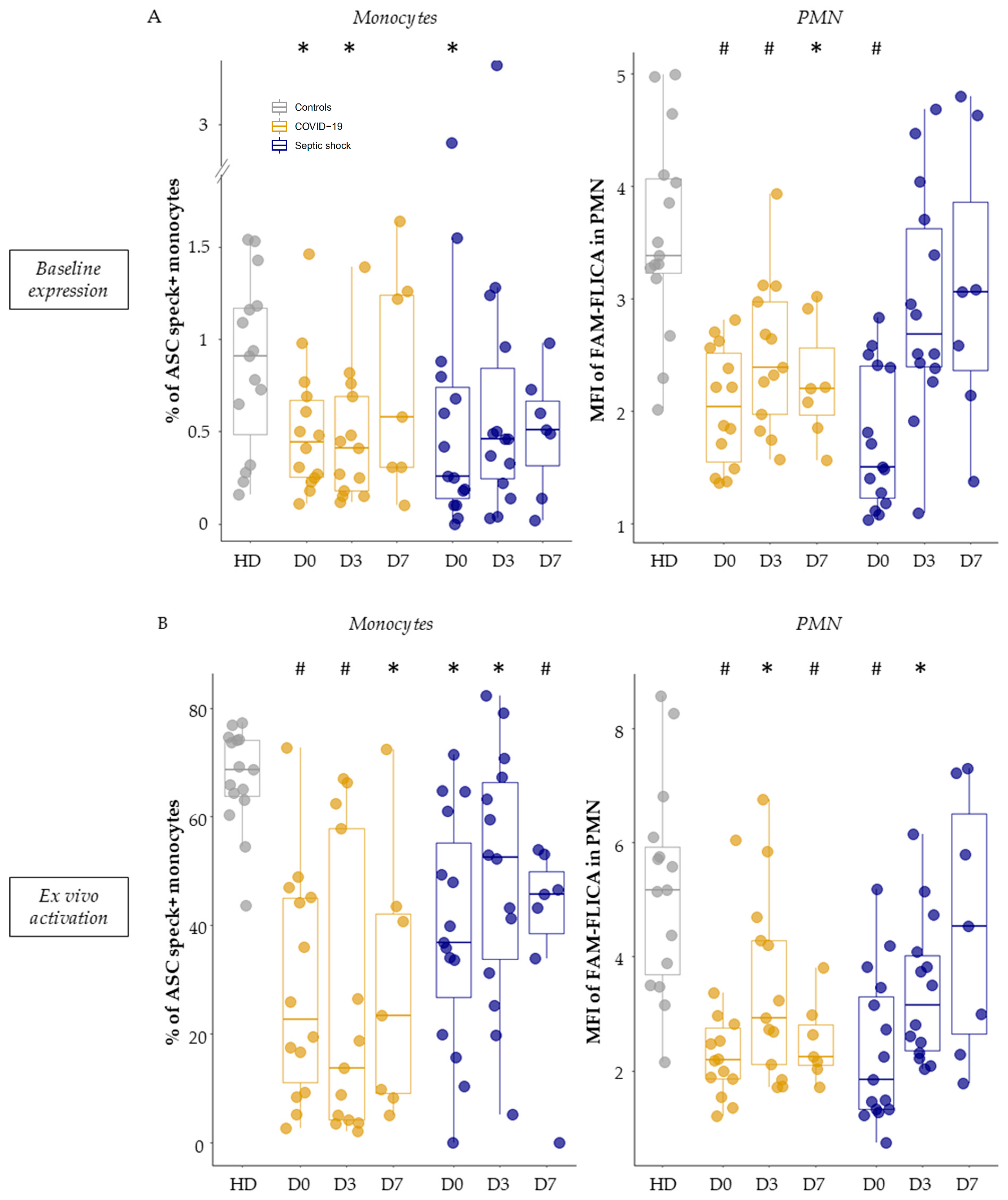
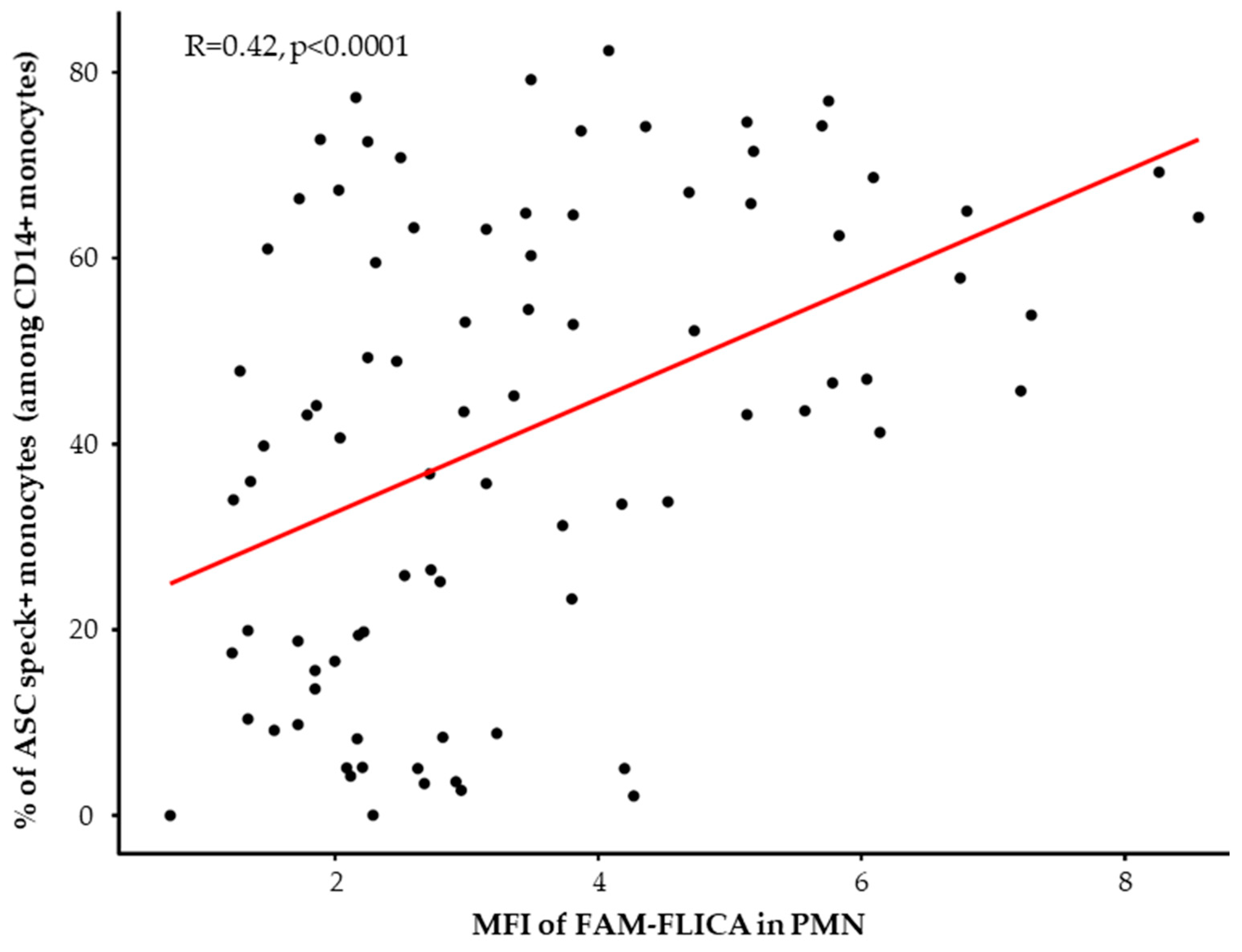
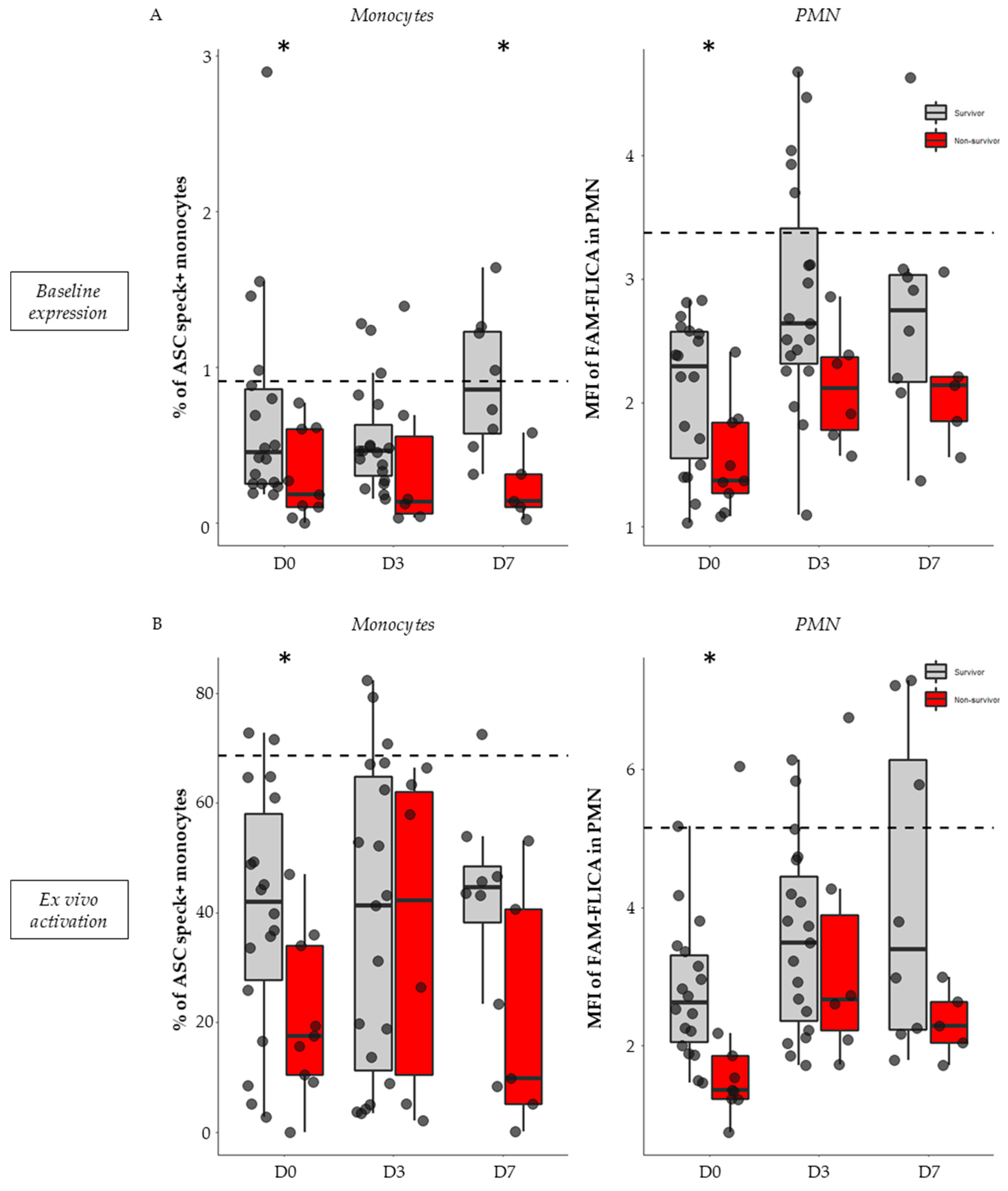
3.3. Basal Expression and Ex Vivo Reactivation of the NLRP3 Inflammasome in Septic Patients
3.4. Association between NLRP3 Inflammasome Activation and 28-Day Mortality in Septic Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, K.; Daniels, R.; Kissoon, N.; Machado, F.R.; Schachter, R.D.; Finfer, S. Recognizing Sepsis as a Global Health Priority—A WHO Resolution. N. Engl. J. Med. 2017, 377, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Sepsis-induced immunosuppression: From cellular dysfunctions to immunotherapy. Nat. Rev. Immunol. 2013, 13, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Groslambert, M.; Py, B.F. Spotlight on the NLRP3 inflammasome pathway. J. Inflamm. Res. 2018, 11, 359–374. [Google Scholar] [CrossRef]
- Booshehri, L.M.; Hoffman, H.M. CAPS and NLRP. J. Clin. Immunol. 2019, 39, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.R.; Kanneganti, T.-D. NLRP3 inflammasome in cancer and metabolic diseases. Nat. Immunol. 2021, 22, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, J.A.; Jarosz-Griffiths, H.H.; Caseley, E.; Lara-Reyna, S.; Poulter, J.A.; Williams-Gray, C.H.; Peckham, D.; McDermott, M.F. Neurodegenerative Disease and the NLRP3 Inflammasome. Front. Pharmacol. 2021, 12, 643254. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Oehrl, S.; Ahmad, F.; Brenner, T.; Uhle, F.; Nusshag, C.; Rupp, C.; Funck, F.; Meisel, S.; Weigand, M.A.; et al. Detection of In Vivo Inflammasome Activation for Predicting Sepsis Mortality. Front. Immunol. 2020, 11, 613745. [Google Scholar] [CrossRef]
- Nasser, S.M.T.; Rana, A.A.; Doffinger, R.; Kafizas, A.; Khan, T.A.; Nasser, S. Elevated free interleukin-18 associated with severity and mortality in prospective cohort study of 206 hospitalised COVID-19 patients. Intensive Care Med. Exp. 2023, 11, 9. [Google Scholar] [CrossRef]
- Rasmussen, G.; Idosa, B.A.; Bäckman, A.; Monecke, S.; Strålin, K.; Särndahl, E.; Söderquist, B. Caspase-1 inflammasome activity in patients with Staphylococcus aureus bacteremia. Microbiol. Immunol. 2019, 63, 487–499. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; de Sá, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Gonçalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218, e20201707. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, A.; Penco, F.; Mollica, H.; Bocca, P.; Prigione, I.; Corcione, A.; Cangelosi, D.; Schena, F.; Del Zotto, G.; Amaro, A.; et al. Spontaneous NLRP3 inflammasome-driven IL-1-β secretion is induced in severe COVID-19 patients and responds to anakinra treatment. J. Allergy Clin. Immunol. 2022, 150, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Giamarellos-Bourboulis, E.J.; van de Veerdonk, F.L.; Mouktaroudi, M.; Raftogiannis, M.; Antonopoulou, A.; Joosten, L.A.B.; Pickkers, P.; Savva, A.; Georgitsi, M.; van der Meer, J.W.; et al. Inhibition of caspase-1 activation in Gram-negative sepsis and experimental endotoxemia. Crit. Care 2011, 15, R27. [Google Scholar] [CrossRef]
- Courjon, J.; Dufies, O.; Robert, A.; Bailly, L.; Torre, C.; Chirio, D.; Contenti, J.; Vitale, S.; Loubatier, C.; Doye, A.; et al. Heterogeneous NLRP3 inflammasome signature in circulating myeloid cells as a biomarker of COVID-19 severity. Blood Adv. 2021, 5, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Cour, M.; Rimmelé, T.; Viel, S.; Yonis, H.; Coudereau, R.; Amaz, C.; Abraham, P.; Monard, C.; Casalegno, J.S.; et al. Longitudinal assessment of IFN-I activity and immune profile in critically ill COVID-19 patients with acute respiratory distress syndrome. Crit. Care 2021, 25, 140. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Textoris, J.; Blein, S.; Rol, M.-L.; Bodinier, M.; Canard, B.; Cortez, P.; Meunier, B.; Tan, L.K.; Tipple, C.; et al. Immune Profiling Demonstrates a Common Immune Signature of Delayed Acquired Immunodeficiency in Patients With Various Etiologies of Severe Injury. Crit. Care Med. 2022, 50, 565–575. [Google Scholar] [CrossRef]
- Gossez, M.; Malcus, C.; Demaret, J.; Frater, J.; Poitevin-Later, F.; Monneret, G. Evaluation of a novel automated volumetric flow cytometer for absolute CD4+ T lymphocyte quantitation. Cytom. Part B Clin. Cytom. 2017, 92, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Demaret, J.; Walencik, A.; Jacob, M.-C.; Timsit, J.-F.; Venet, F.; Lepape, A.; Monneret, G. Inter-laboratory assessment of flow cytometric monocyte HLA-DR expression in clinical samples. Cytom. B Clin. Cytom. 2013, 84, 59–62. [Google Scholar] [CrossRef]
- Coudereau, R.; Gossez, M.; Py, B.F.; Henry, T.; Lukaszewicz, A.-C.; Monneret, G.; Venet, F. Monitoring NLRP3 Inflammasome Activation and Exhaustion in Clinical Samples: A Refined Flow Cytometry Protocol for ASC Speck Formation Measurement Directly in Whole Blood after Ex Vivo Stimulation. Cells 2022, 11, 3306. [Google Scholar]
- Rogers, A.J.; Guan, J.; Trtchounian, A.; Hunninghake, G.M.; Kaimal, R.; Desai, M.; Kozikowski, L.-A.; DeSouza, L.; Mogan, S.; Liu, K.D.; et al. Association of Elevated Plasma Interleukin-18 Level With Increased Mortality in a Clinical Trial of Statin Treatment for Acute Respiratory Distress Syndrome. Crit. Care Med. 2019, 47, 1089–1096. [Google Scholar] [CrossRef]
- Weighardt, H.; Heidecke, C.D.; Emmanuilidis, K.; Maier, S.; Bartels, H.; Siewert, J.R.; Holzmann, B. Sepsis after major visceral surgery is associated with sustained and interferon-gamma-resistant defects of monocyte cytokine production. Surgery 2000, 127, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, J.J.; Martínez-Banaclocha, H.; Angosto-Bazarra, D.; de Torre-Minguela, C.; Baroja-Mazo, A.; Alarcón-Vila, C.; Martínez-Alarcón, L.; Amores-Iniesta, J.; Martín-Sánchez, F.; Ercole, G.A.; et al. P2X7 receptor induces mitochondrial failure in monocytes and compromises NLRP3 inflammasome activation during sepsis. Nat. Commun. 2019, 10, 2711. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.M.; Adib-Conquy, M.; Cloëz-Tayarani, I.; Fitting, C. Immunodepression in sepsis and SIRS assessed by ex vivo cytokine production is not a generalized phenomenon: A review. J. Endotoxin Res. 2001, 7, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.-M.; Annane, D. Compartmentalization of the inflammatory response in sepsis and SIRS. J. Endotoxin Res. 2006, 12, 151–170. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, W.-C.; Chen, X.-Y.; Wang, X.; Li, J.-L.; Zhang, X. Gasdermin D-mediated pyroptosis: Mechanisms, diseases, and inhibitors. Front. Immunol. 2023, 14, 1178662. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Liu, Q.; Zheng, Q.; Dong, X.; Liu, X.; Gao, W.; Bai, X.; Li, Z. Caspase-1-Dependent Pyroptosis of Peripheral Blood Mononuclear Cells Is Associated with the Severity and Mortality of Septic Patients. BioMed Res. Int. 2020, 2020, 9152140. [Google Scholar] [CrossRef]
- Allantaz-Frager, F.; Turrel-Davin, F.; Venet, F.; Monnin, C.; De Saint Jean, A.; Barbalat, V.; Cerrato, E.; Pachot, A.; Lepape, A.; Monneret, G. Identification of biomarkers of response to IFNg during endotoxin tolerance: Application to septic shock. PLoS ONE 2013, 8, e68218. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Septic Patients (n = 32) |
|---|---|
| Age at admission (years) | 64 (61–69) |
| Gender–Male n (%) | 25 (78) |
| Main admission category n (%) Medical Surgical | 22 (69) 10 (31) |
| SAPSII score at admission | 38 (33–46) |
| SOFA score at admission | 6 (3–10) |
| Charlson score | 1 (0–3) |
| Infection diagnosis n (%) Microbiology Surgery Other | 26 (81) 3 (9) 3 (9) |
| Microbiologically documented n (%) Bacilli gram − Cocci gram + SARS-CoV-2 | 7 (22) 4 (12) 15 (46) |
| Respiratory dysfunction Mechanical ventilation n (%) PaO2/FiO2 | 18 (56) 177 (86–279) |
| Site of primary infection n (%) Pulmonary Abdominal Other | 16 (50) 10 (31) 4 (12) |
| Follow-up n (%) 28-day non survivors Secondary nosocomial infections | 9 (28) 3 (9) |
| Immunological parameters (at Day 3) mHLA-DR (AB/C) CD4+ T cell count (cells/µL) | 5914 (4748–7954) 339 (140–553) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coudereau, R.; Monneret, G.; Lukaszewicz, A.-C.; Py, B.F.; Argaud, L.; Cour, M.; Bidar, F.; Gossez, M.; Venet, F. Altered Ex Vivo NLRP3 Inflammasome Activation Is Associated with 28-Day Mortality in Septic Patients. Viruses 2023, 15, 2419. https://doi.org/10.3390/v15122419
Coudereau R, Monneret G, Lukaszewicz A-C, Py BF, Argaud L, Cour M, Bidar F, Gossez M, Venet F. Altered Ex Vivo NLRP3 Inflammasome Activation Is Associated with 28-Day Mortality in Septic Patients. Viruses. 2023; 15(12):2419. https://doi.org/10.3390/v15122419
Chicago/Turabian StyleCoudereau, Rémy, Guillaume Monneret, Anne-Claire Lukaszewicz, Bénédicte F. Py, Laurent Argaud, Martin Cour, Frank Bidar, Morgane Gossez, and Fabienne Venet. 2023. "Altered Ex Vivo NLRP3 Inflammasome Activation Is Associated with 28-Day Mortality in Septic Patients" Viruses 15, no. 12: 2419. https://doi.org/10.3390/v15122419
APA StyleCoudereau, R., Monneret, G., Lukaszewicz, A.-C., Py, B. F., Argaud, L., Cour, M., Bidar, F., Gossez, M., & Venet, F. (2023). Altered Ex Vivo NLRP3 Inflammasome Activation Is Associated with 28-Day Mortality in Septic Patients. Viruses, 15(12), 2419. https://doi.org/10.3390/v15122419