
Remdesivir Derivative VV116 Is a Potential Broad-Spectrum Inhibitor of Both Human and Animal Coronaviruses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines, Viruses, and Reagents
2.2. Cytotoxicity Assay
2.3. Viral Yield Reduction Assay
2.4. RNA Extraction and Real-Time PCR
2.5. Time-of-Addition Assay
2.6. Plaque Assay
2.7. Immunofluorescence Assay
2.8. rNTP Competitive Inhibition Assay
2.9. In Vivo Study
2.10. Statistical Analysis
3. Results
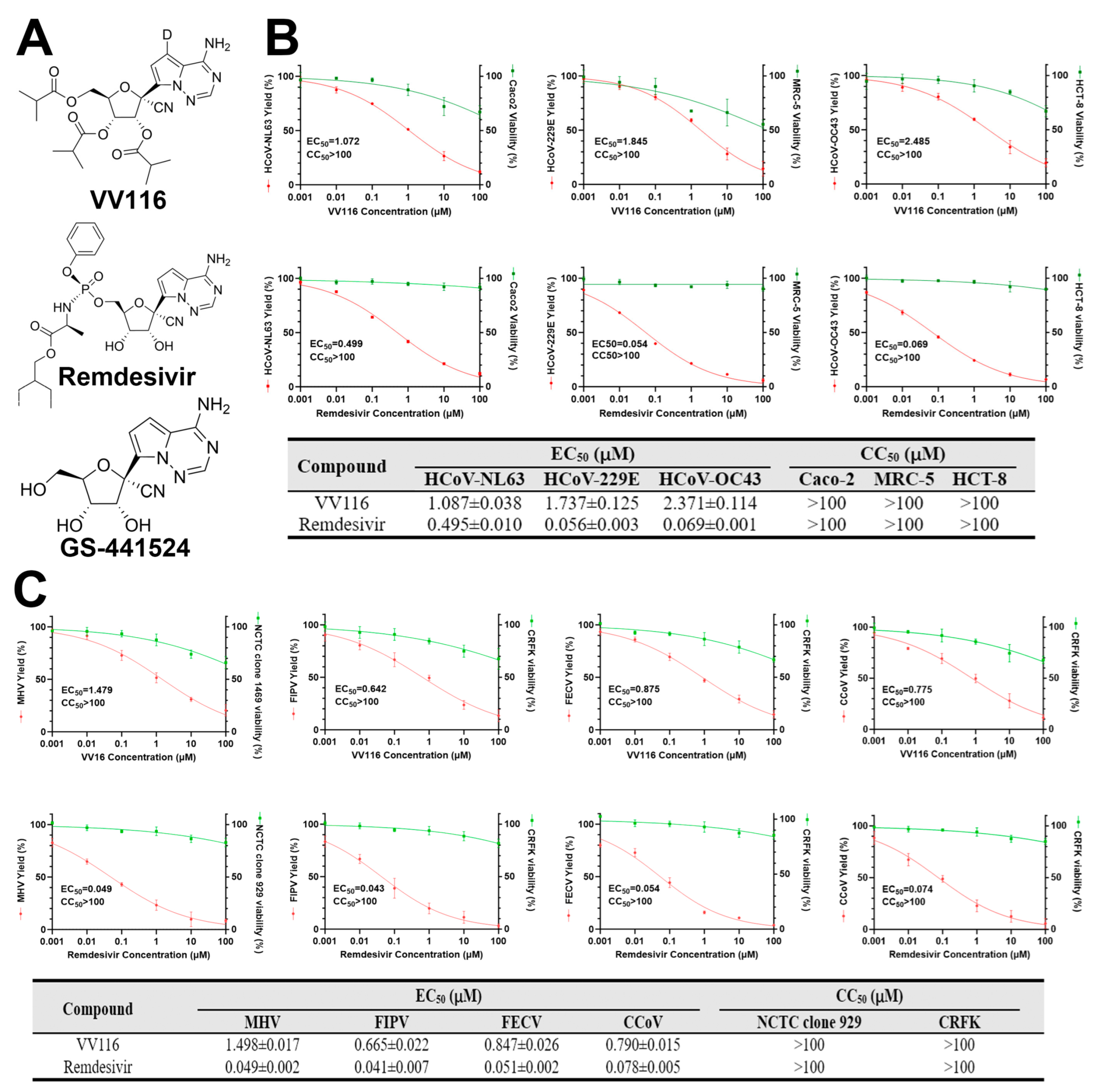
3.1. VV116 Exhibits Broad-Spectrum Antiviral Activity against Human and Animal Coronaviruses
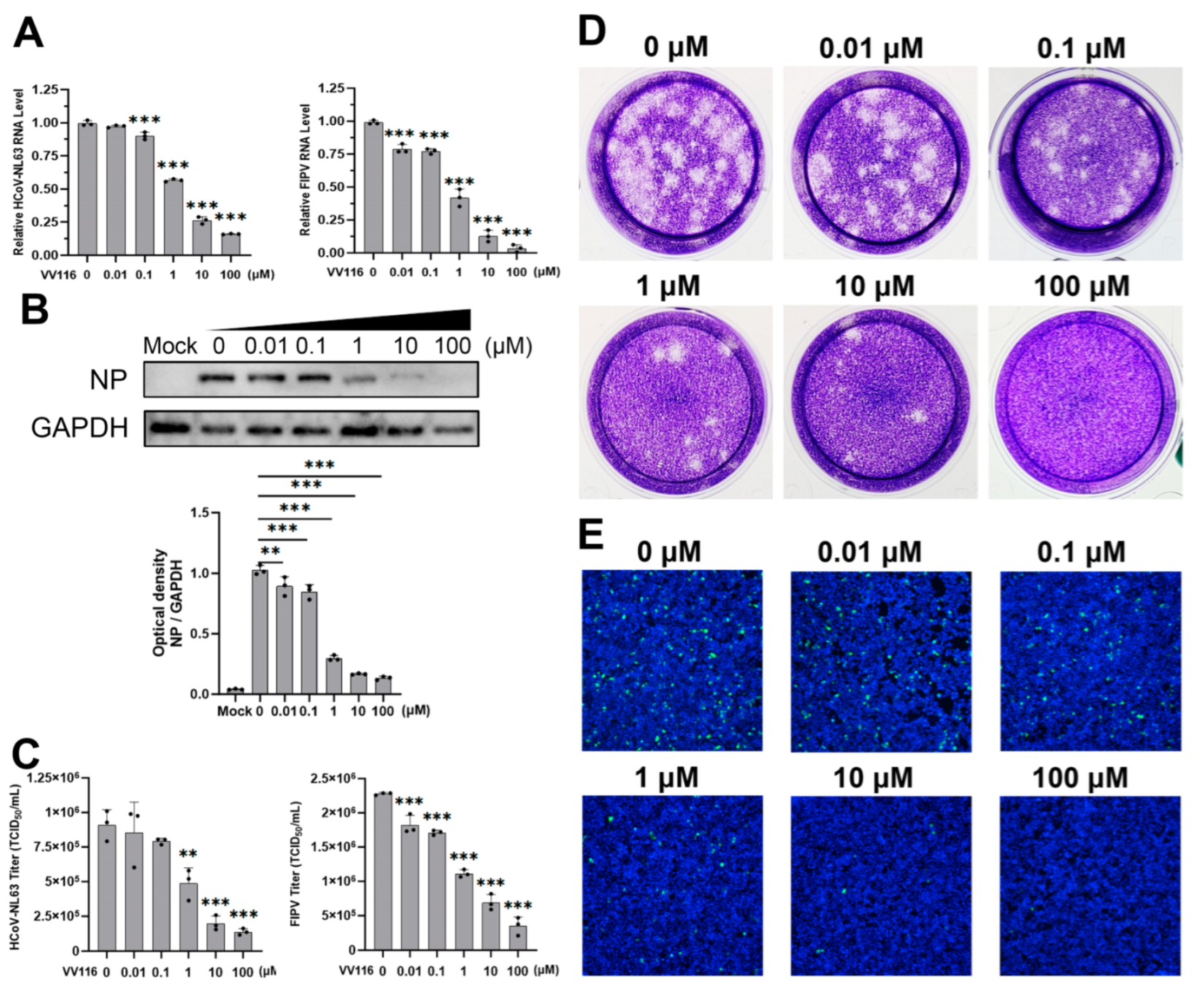
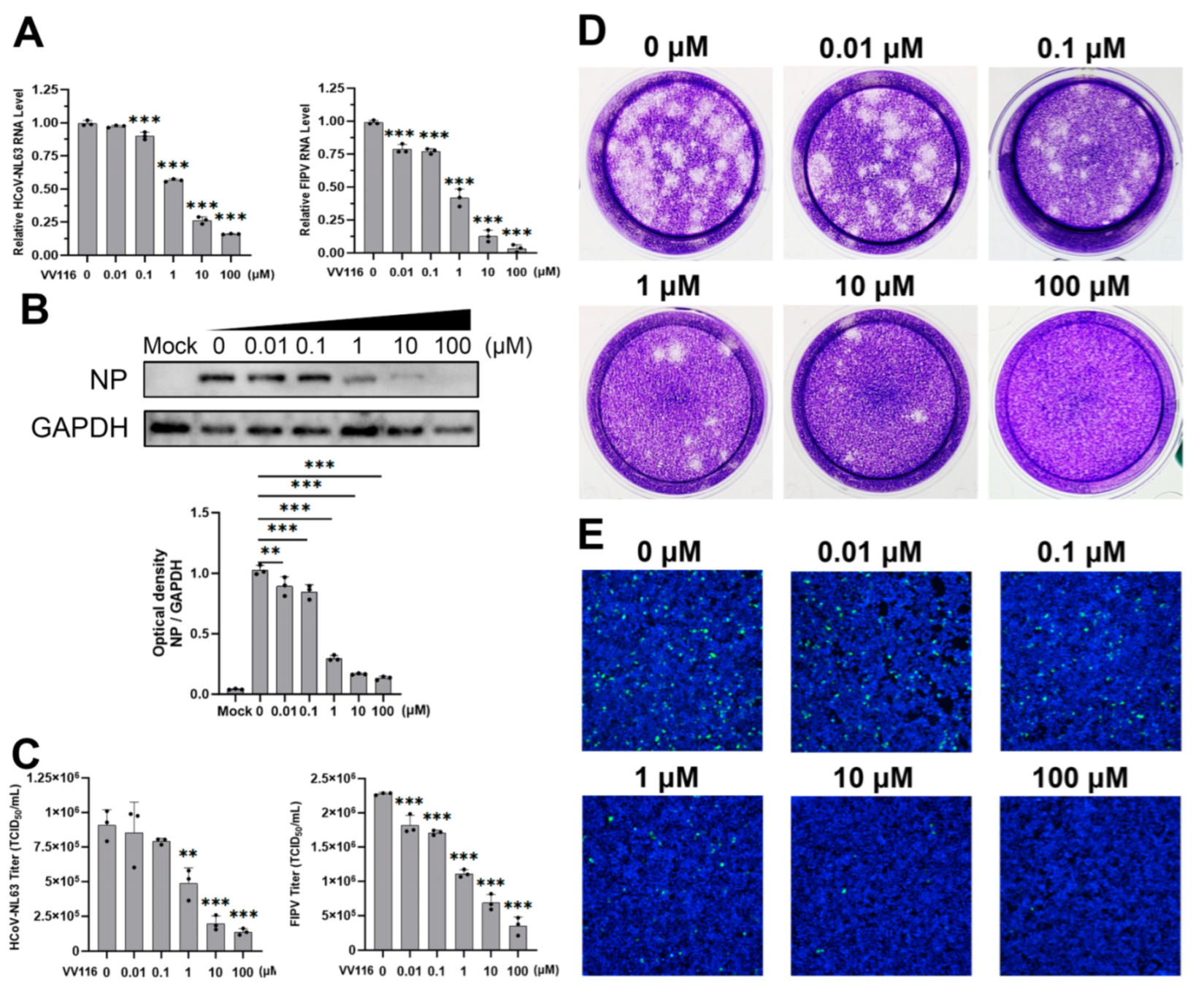
3.2. Confirmation of the Antiviral Activity of VV116 against HCoV-NL63 and FIPV
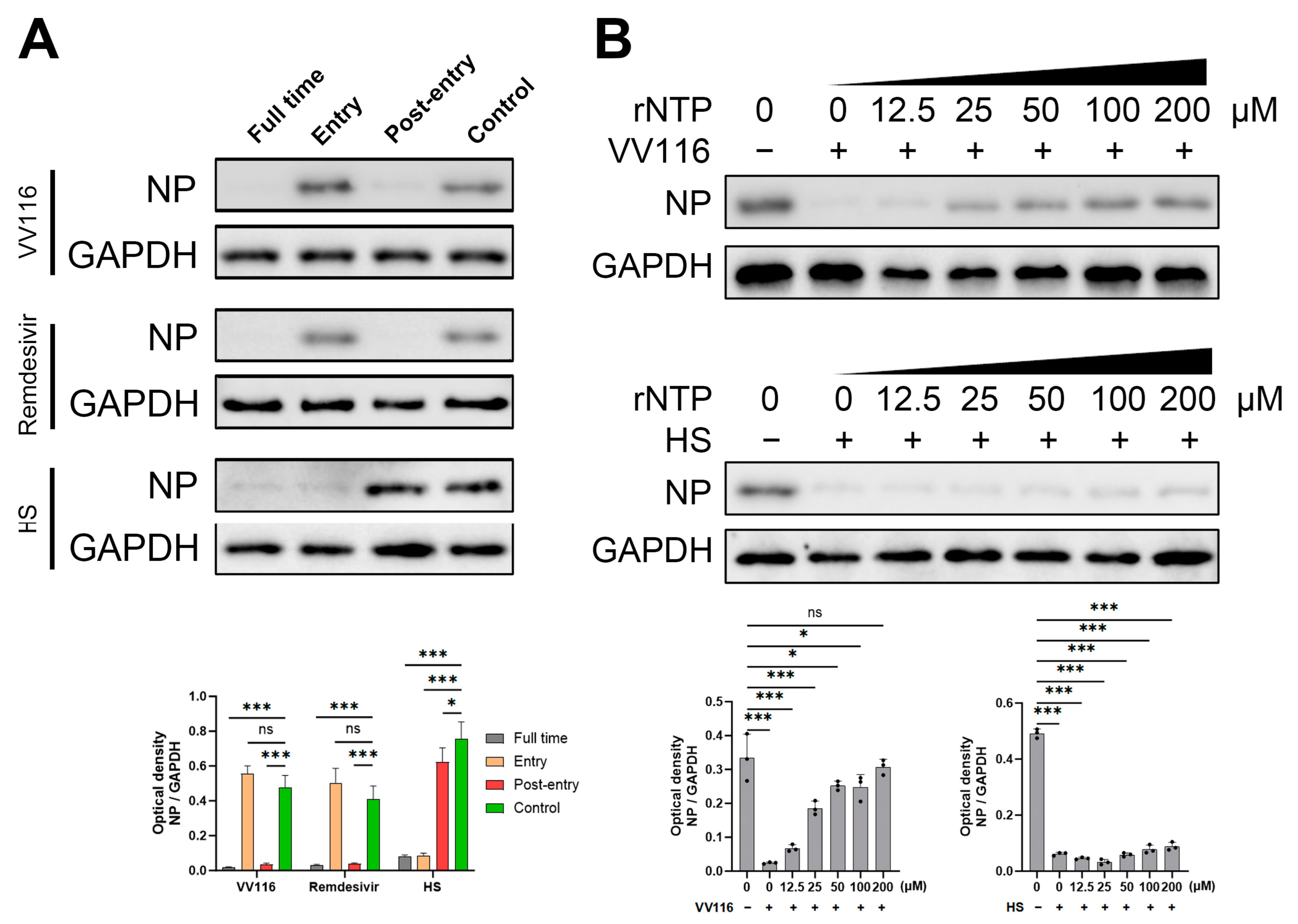
3.3. Mechanism of Antiviral Activity
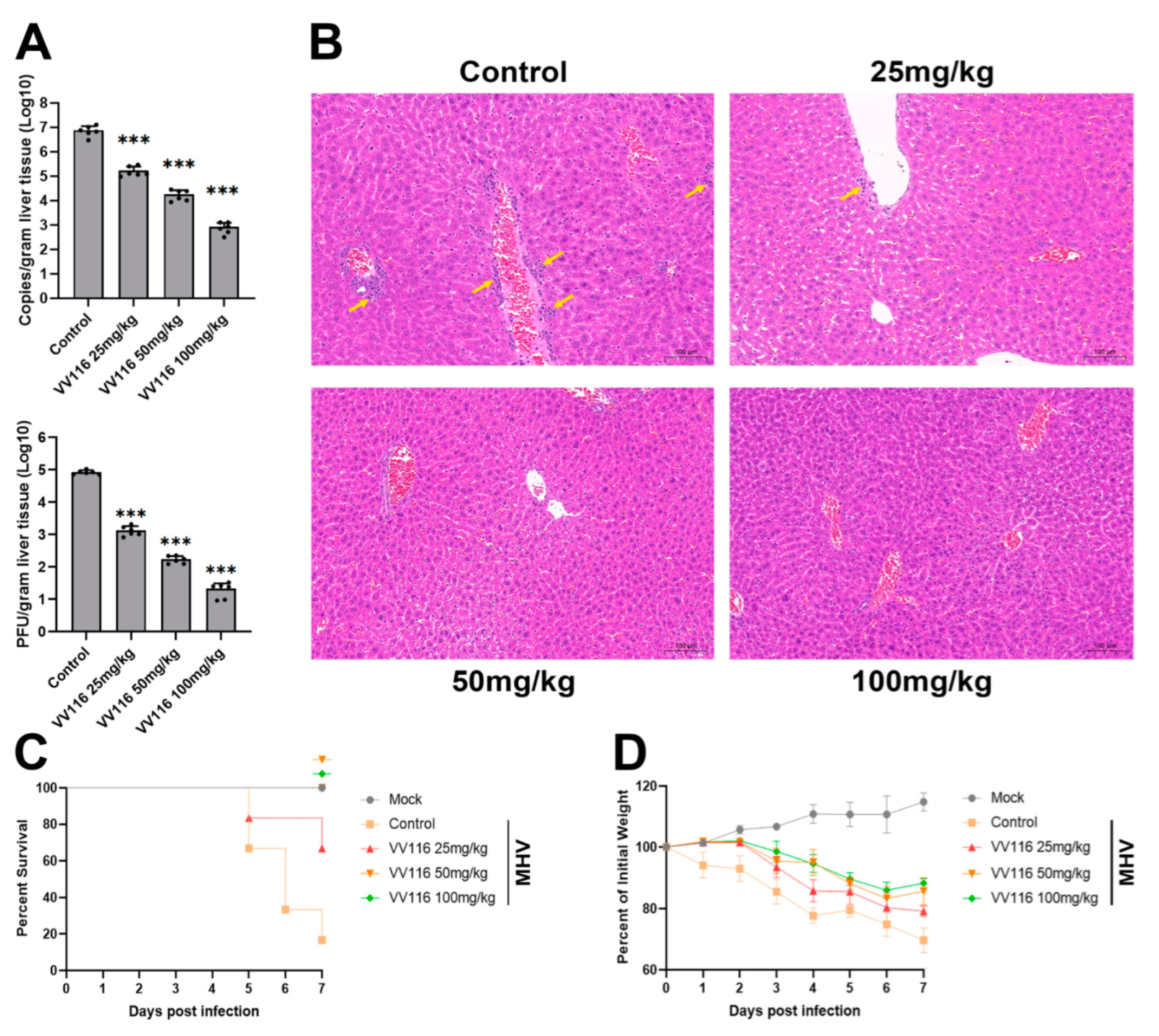
3.4. VV116 Exhibits Substantial Anti-Coronavirus Activity in an MHV-Infected Mouse Model
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tyrrell, D.A.J.; Bynoe, M.L. Cultivation of a Novel Type of Common-Cold Virus in Organ Cultures. Br. Med. J. 1965, 1, 1467–1470. [Google Scholar] [CrossRef] [PubMed]
- Mesel-Lemoine, M.; Millet, J.; Vidalain, P.-O.; Law, H.; Vabret, A.; Lorin, V.; Escriou, N.; Albert, M.L.; Nal, B.; Tangy, F. A Human Coronavirus Responsible for the Common Cold Massively Kills Dendritic Cells but Not Monocytes. J. Virol. 2012, 86, 7577–7587. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, T.; Endo, R.; Ma, X.; Ishiguro, N.; Kikuta, H. Detection of Human Coronavirus NL63 in Young Children with Bronchiolitis. J. Med. Virol. 2005, 75, 463–465. [Google Scholar] [CrossRef] [PubMed]
- Pene, F.; Merlat, A.; Vabret, A.; Rozenberg, F.; Buzyn, A.; Dreyfus, F.; Cariou, A.; Freymuth, F.; Lebon, P. Coronavirus 229E-Related Pneumonia in Immunocompromised Patients. Clin. Infect. Dis. 2003, 37, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Chan-Yeung, M.; Xu, R.-H. SARS: Epidemiology. Respirology 2003, 8, S9–S14. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; El Morabet, R.; Khan, R.A.; Bindajam, A.; Alqadhi, S.; Alsubih, M.; Khan, N.A. Where We Missed? Middle East Respiratory Syndrome (MERS-CoV) Epidemiology in Saudi Arabia; 2012–2019. Sci. Total Environ. 2020, 747, 141369. [Google Scholar] [CrossRef] [PubMed]
- Gaye, B.; Fanidi, A.; Jouven, X. Denominator Matters in Estimating COVID-19 Mortality Rates. Eur. Heart J. 2020, 41, 3500. [Google Scholar] [CrossRef]
- Grabherr, S.; Ludewig, B.; Pikor, N.B. Insights into Coronavirus Immunity Taught by the Murine Coronavirus. Eur. J. Immunol. 2021, 51, 1062–1070. [Google Scholar] [CrossRef]
- Ward, J.M. Morphogenesis of a Virus in Cats with Experimental Feline Infectious Peritonitis. Virology 1970, 41, 191–194. [Google Scholar] [CrossRef]
- Rottier, P.J.M.; Nakamura, K.; Schellen, P.; Volders, H.; Haijema, B.J. Acquisition of Macrophage Tropism during the Pathogenesis of Feline Infectious Peritonitis Is Determined by Mutations in the Feline Coronavirus Spike Protein. J. Virol. 2005, 79, 14122–14130. [Google Scholar] [CrossRef]
- Binn, L.N.; Lazar, E.C.; Keenan, K.P.; Huxsoll, D.L.; Marchwicki, R.H.; Strano, A.J. Recovery and Characterization of a Coronavirus from Military Dogs with Diarrhea. In Proceedings of the 78th Annual Meeting of the United States Animal Health Association, Roanoke, Virginia, 13–17 October 1974; pp. 359–366. [Google Scholar]
- Tennant, B.J.; Gaskell, R.M.; Kelly, D.F.; Carter, S.D.; Gaskell, C.J. Canine Coronavirus Infection in the Dog Following Oronasal Inoculation. Res. Vet. Sci. 1991, 51, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Seley-Radtke, K.L.; Yates, M.K. The Evolution of Nucleoside Analogue Antivirals: A Review for Chemists and Non-Chemists. Part 1: Early Structural Modifications to the Nucleoside Scaffold. Antivir. Res. 2018, 154, 66–86. [Google Scholar] [CrossRef] [PubMed]
- Yates, M.K.; Seley-Radtke, K.L. The Evolution of Antiviral Nucleoside Analogues: A Review for Chemists and Non-Chemists. Part II: Complex Modifications to the Nucleoside Scaffold. Antivir. Res. 2019, 162, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of COVID-19—Final Report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef] [PubMed]
- Warren, T.K.; Jordan, R.; Lo, M.K.; Ray, A.S.; Mackman, R.L.; Soloveva, V.; Siegel, D.; Perron, M.; Bannister, R.; Hui, H.C.; et al. Therapeutic Efficacy of the Small Molecule GS-5734 against Ebola Virus in Rhesus Monkeys. Nature 2016, 531, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, T.P.; Sims, A.C.; Leist, S.R.; Schäfer, A.; Won, J.; Brown, A.J.; Montgomery, S.A.; Hogg, A.; Babusis, D.; Clarke, M.O.; et al. Comparative Therapeutic Efficacy of Remdesivir and Combination Lopinavir, Ritonavir, and Interferon Beta against MERS-CoV. Nat. Commun. 2020, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, T.P.; Sims, A.C.; Graham, R.L.; Menachery, V.D.; Gralinski, L.E.; Case, J.B.; Leist, S.R.; Pyrc, K.; Feng, J.Y.; Trantcheva, I.; et al. Broad-Spectrum Antiviral GS-5734 Inhibits Both Epidemic and Zoonotic Coronaviruses. Sci. Transl. Med. 2017, 9, eaal3653. [Google Scholar] [CrossRef]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Smith, E.C.; Case, J.B.; Feng, J.Y.; Jordan, R.; et al. Coronavirus Susceptibility to the Antiviral Remdesivir (GS-5734) Is Mediated by the Viral Polymerase and the Proofreading Exoribonuclease. mBio 2018, 9, e00221-18. [Google Scholar] [CrossRef]
- Bravo, J.P.K.; Dangerfield, T.L.; Taylor, D.W.; Johnson, K.A. Remdesivir Is a Delayed Translocation Inhibitor of SARS-CoV-2 Replication. Mol. Cell 2021, 81, 1548–1552.e4. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and Chloroquine Effectively Inhibit the Recently Emerged Novel Coronavirus (2019-nCoV) in Vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Pizzorno, A.; Padey, B.; Dubois, J.; Julien, T.; Traversier, A.; Dulière, V.; Brun, P.; Lina, B.; Rosa-Calatrava, M.; Terrier, O. In Vitro Evaluation of Antiviral Activity of Single and Combined Repurposable Drugs against SARS-CoV-2. Antivir. Res. 2020, 181, 104878. [Google Scholar] [CrossRef]
- Ogando, N.S.; Dalebout, T.J.; Zevenhoven-Dobbe, J.C.; Limpens, R.W.A.L.; van der Meer, Y.; Caly, L.; Druce, J.; de Vries, J.J.C.; Kikkert, M.; Bárcena, M.; et al. SARS-Coronavirus-2 Replication in Vero E6 Cells: Replication Kinetics, Rapid Adaptation and Cytopathology. J. Gen. Virol. 2020, 101, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yin, W.; Zhang, Y.; Shang, W.; Wang, Z.; Luan, X.; Tian, G.; Aisa, H.A.; Xu, Y.; Xiao, G.; et al. Design and Development of an Oral Remdesivir Derivative VV116 against SARS-CoV-2. Cell Res. 2021, 31, 1212–1214. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.W. VV116 as a Potential Treatment for COVID-19. Expert Opin. Pharmacother. 2023, 24, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Gao, W.; Bao, H.; Feng, H.; Mei, S.; Chen, P.; Gao, Y.; Cui, Z.; Zhang, Q.; Meng, X.; et al. VV116 versus Nirmatrelvir–Ritonavir for Oral Treatment of COVID-19. N. Engl. J. Med. 2023, 388, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, Y.; Zheng, W.; Shang, W.; Wu, Y.; Li, N.; Xiong, J.; Jiang, H.; Shen, J.; Xiao, G.; et al. Oral Remdesivir Derivative VV116 Is a Potent Inhibitor of Respiratory Syncytial Virus with Efficacy in Mouse Model. Signal Transduct. Target. Ther. 2022, 7, 123. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Wang, Y.; Zhang, M.; Xie, Y.; Wu, Q.; Liang, L.; Cao, Y.; Duan, H.; Tian, G.; Ma, J.; et al. Safety, Tolerability, and Pharmacokinetics of VV116, an Oral Nucleoside Analog against SARS-CoV-2, in Chinese Healthy Subjects. Acta Pharmacol. Sin. 2022, 43, 3130–3138. [Google Scholar] [CrossRef]
- Shen, Y.; Ai, J.; Lin, N.; Zhang, H.; Li, Y.; Wang, H.; Wang, S.; Wang, Z.; Li, T.; Sun, F.; et al. An Open, Prospective Cohort Study of VV116 in Chinese Participants Infected with SARS-CoV-2 Omicron Variants. Emerg. Microbes Infect. 2022, 11, 1518–1523. [Google Scholar] [CrossRef]
- Zhu, K.-W. Deuremidevir and Simnotrelvir–Ritonavir for the Treatment of COVID-19. ACS Pharmacol. Transl. Sci. 2023, 6, 1306–1309. [Google Scholar] [CrossRef]
- Milewska, A.; Zarebski, M.; Nowak, P.; Stozek, K.; Potempa, J.; Pyrc, K. Human Coronavirus NL63 Utilizes Heparan Sulfate Proteoglycans for Attachment to Target Cells. J. Virol. 2014, 88, 13221–13230. [Google Scholar] [CrossRef]
- Liu, D.X.; Liang, J.Q.; Fung, T.S. Human Coronavirus-229E, -OC43, -NL63, and -HKU1 (Coronaviridae). In Encyclopedia of Virology; Academic Press: Cambridge, MA, USA, 2021; Volume 2, pp. 428–440. [Google Scholar] [CrossRef]
- Vijgen, L.; Keyaerts, E.; Moës, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.-M.; Van Ranst, M. Complete Genomic Sequence of Human Coronavirus OC43: Molecular Clock Analysis Suggests a Relatively Recent Zoonotic Coronavirus Transmission Event. J. Virol. 2005, 79, 1595–1604. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Eckerle, I.; Memish, Z.A.; Liljander, A.M.; Dijkman, R.; Jonsdottir, H.; Juma Ngeiywa, K.J.; Kamau, E.; Younan, M.; Al Masri, M.; et al. Link of a Ubiquitous Human Coronavirus to Dromedary Camels. Proc. Natl. Acad. Sci. USA 2016, 113, 9864–9869. [Google Scholar] [CrossRef] [PubMed]
- Nova, N. Cross-Species Transmission of Coronaviruses in Humans and Domestic Mammals, What Are the Ecological Mechanisms Driving Transmission, Spillover, and Disease Emergence? Front. Public Health 2021, 9, 717941. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.S.; Mullis, L.B.; Pereira, O.; Saif, L.J.; Vlasova, A.; Zhang, X.; Owens, R.J.; Paulson, D.; Taylor, D.; Haynes, L.M.; et al. Human Respiratory Coronaviruses Detected In Patients with Influenza-Like Illness in Arkansas, USA. Virol. Mycol. 2014, 2014, 004. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Diaz, A.; Damtie, D.; Xiu, L.; Toh, T.-H.; Lee, J.S.-Y.; Saif, L.J.; Gray, G.C. Novel Canine Coronavirus Isolated from a Hospitalized Patient With Pneumonia in East Malaysia. Clin. Infect. Dis. 2022, 74, 446–454. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′ to 3′) |
|---|---|
| NL63 qPCR-F | GTGATGCATATGCTAATTTG |
| NL63 qPCR-R | CTCTTGCAGGTATAATCCTA |
| FIPV qPCR-F | CCGAGGAATTACTGGTCATCGCG |
| FIPV qPCR-R | GCTCTTCCATTGTTGGCTCGTC |
| GAPDH qPCR-F | AAGGCTGTGGGCAAGG |
| GAPDH qPCR-R | TGGAGGAGTGGGTGTCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Zhang, M.; Hu, C.; Song, H.; Mei, Y.; Liu, Y.; Zhang, Q. Remdesivir Derivative VV116 Is a Potential Broad-Spectrum Inhibitor of Both Human and Animal Coronaviruses. Viruses 2023, 15, 2295. https://doi.org/10.3390/v15122295
Liu W, Zhang M, Hu C, Song H, Mei Y, Liu Y, Zhang Q. Remdesivir Derivative VV116 Is a Potential Broad-Spectrum Inhibitor of Both Human and Animal Coronaviruses. Viruses. 2023; 15(12):2295. https://doi.org/10.3390/v15122295
Chicago/Turabian StyleLiu, Weiyong, Min Zhang, Chengxiu Hu, Huijuan Song, Yi Mei, Yingle Liu, and Qi Zhang. 2023. "Remdesivir Derivative VV116 Is a Potential Broad-Spectrum Inhibitor of Both Human and Animal Coronaviruses" Viruses 15, no. 12: 2295. https://doi.org/10.3390/v15122295
APA StyleLiu, W., Zhang, M., Hu, C., Song, H., Mei, Y., Liu, Y., & Zhang, Q. (2023). Remdesivir Derivative VV116 Is a Potential Broad-Spectrum Inhibitor of Both Human and Animal Coronaviruses. Viruses, 15(12), 2295. https://doi.org/10.3390/v15122295