
PSGL-1, a Strategic Biomarker for Pathological Conditions in HIV Infection: A Hypothesis Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Hypothesis 1: A Marker of Immune Depletion Prior to ART Initiation

3. Hypothesis 2: A Marker of Immune Reconstitution after ART Initiation
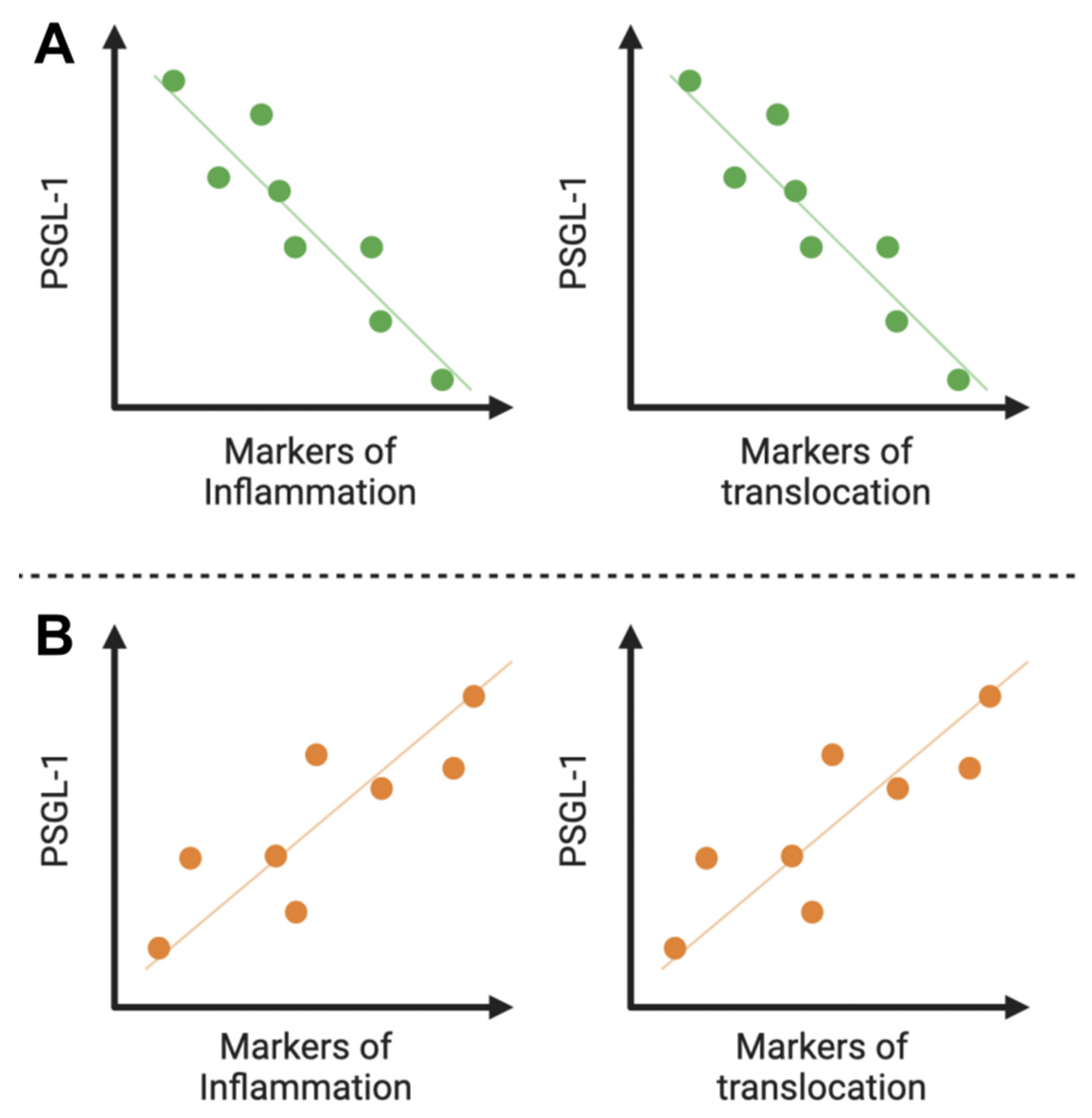
4. Hypothesis 3: A Marker of Inflammation and/or Translocation
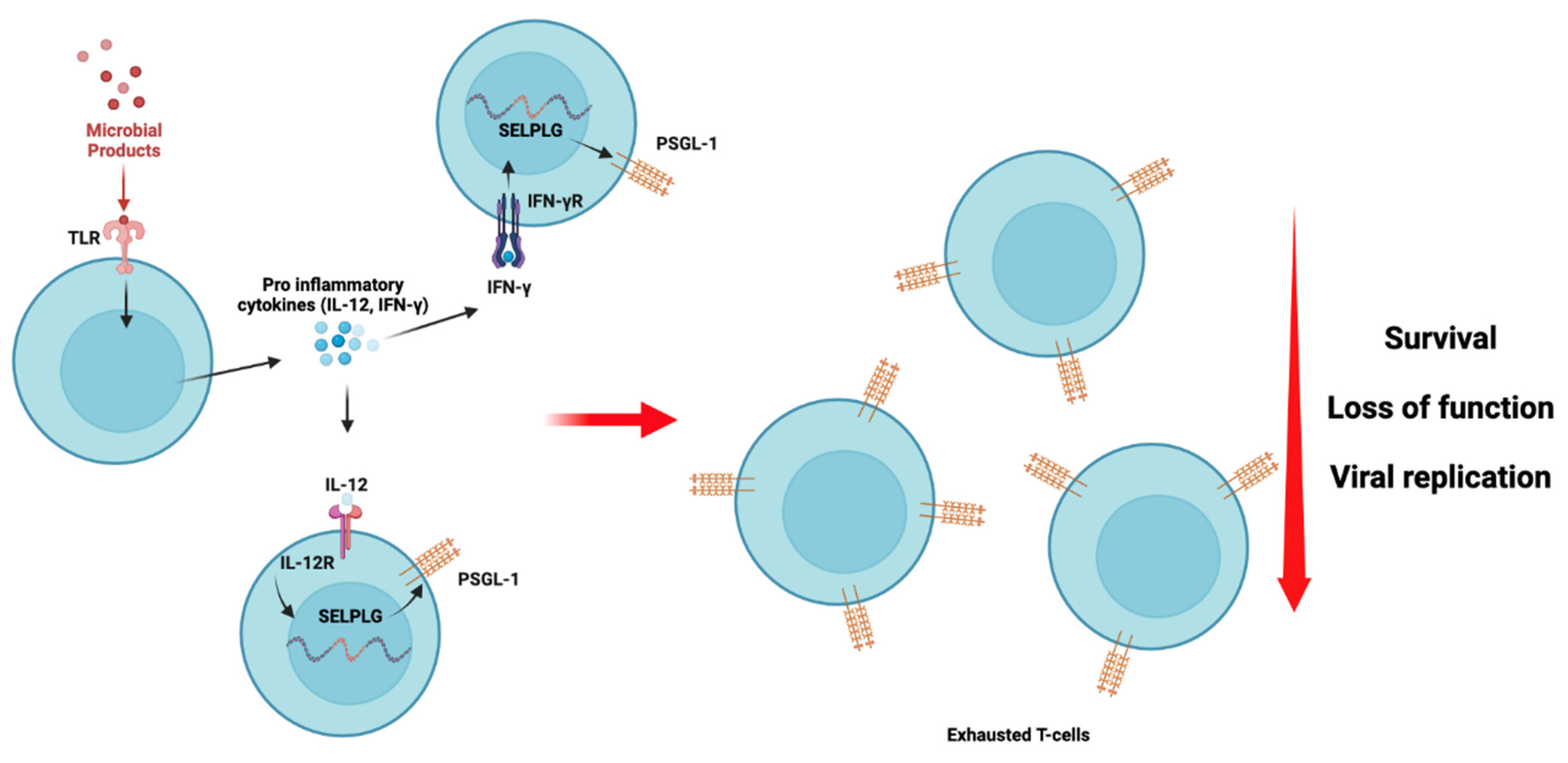
5. Hypothesis 4: A Marker of Cell Exhaustion in ART-Treated Individuals
6. Hypothesis 5: A Marker of Potential or Latent Cancer in INRs
7. Additional Challenges
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tinoco, R.; Bradley, L.M. Targeting the PSGL-1 pathway for immune modulation. Immunotherapy 2017, 9, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Tinoco, R.; Otero, D.C.; Takahashi, A.A.; Bradley, L.M. PSGL-1: A New Player in the Immune Checkpoint Landscape. Trends Immunol. 2017, 38, 323–335. [Google Scholar] [CrossRef]
- Tanaka, T. Leukocyte adhesion molecules. In Encyclopedia of Immunobiology; Ratcliffe, M., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 505–511. [Google Scholar]
- Borges, E.; Eytner, R.; Moll, T.; Steegmaier, M.; Campbell, M.A.; Ley, K.; Mossmann, H.; Vestweber, D. The P-selectin glycoprotein ligand-1 is important for recruitment of neutrophils into inflamed mouse peritoneum. Blood 1997, 90, 1934–1942. [Google Scholar] [CrossRef] [PubMed]
- Asaduzzaman, M.; Mihaescu, A.; Wang, Y.; Sato, T.; Thorlacius, H. P-selectin and P-selectin glycoprotein ligand 1 mediate rolling of activated CD8+ T cells in inflamed colonic venules. J. Investig. Med. 2009, 57, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Martín-Fontecha, A.; Baumjohann, D.; Guarda, G.; Reboldi, A.; Hons, M.; Lanzavecchia, A.; Sallusto, F. CD40L+ CD4+ memory T cells migrate in a CD62P-dependent fashion into reactive lymph nodes and license dendritic cells for T cell priming. J. Exp. Med. 2008, 205, 2561–2574. [Google Scholar] [CrossRef] [PubMed]
- Ley, K.; Kansas, G.S. Selectins in T-cell recruitment to non-lymphoid tissues and sites of inflammation. Nat. Rev. Immunol. 2004, 4, 325–335. [Google Scholar] [CrossRef]
- Lam, F.W.; Burns, A.R.; Smith, C.W.; Rumbaut, R.E. Platelets enhance neutrophil transendothelial migration via P-selectin glycoprotein ligand-1. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H468–H475. [Google Scholar] [CrossRef]
- Zaongo, S.D.; Liu, Y.; Harypursat, V.; Song, F.; Xia, H.; Ma, P.; Chen, Y. P-Selectin Glycoprotein Ligand 1: A Potential HIV-1 Therapeutic Target. Front. Immunol. 2021, 12, 710121. [Google Scholar] [CrossRef]
- Kum, W.W.; Lee, S.; Grassl, G.A.; Bidshahri, R.; Hsu, K.; Ziltener, H.J.; Finlay, B.B. Lack of functional P-selectin ligand exacerbates Salmonella serovar typhimurium infection. J. Immunol. 2009, 182, 6550–6561. [Google Scholar] [CrossRef]
- He, S.; Waheed, A.A.; Hetrick, B.; Dabbagh, D.; Akhrymuk, I.V.; Kehn-Hall, K.; Freed, E.O.; Wu, Y. PSGL-1 Inhibits the Incorporation of SARS-CoV and SARS-CoV-2 Spike Glycoproteins into Pseudovirions and Impairs Pseudovirus Attachment and Infectivity. Viruses 2020, 13, 46. [Google Scholar] [CrossRef]
- Fu, Y.; He, S.; Waheed, A.A.; Dabbagh, D.; Zhou, Z.; Trinité, B.; Wang, Z.; Yu, J.; Wang, D.; Li, F.; et al. PSGL-1 restricts HIV-1 infectivity by blocking virus particle attachment to target cells. Proc. Natl. Acad. Sci. USA 2020, 117, 9537–9545. [Google Scholar] [CrossRef] [PubMed]
- Bukrinskaya, A.; Brichacek, B.; Mann, A.; Stevenson, M. Establishment of a functional human immunodeficiency virus type 1 (HIV-1) reverse transcription complex involves the cytoskeleton. J. Exp. Med. 1998, 188, 2113–2125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fu, Y.; Wang, Q.; Li, M.; Zhou, Z.; Dabbagh, D.; Fu, C.; Zhang, H.; Li, S.; Zhang, T.; et al. Proteomic profiling of HIV-1 infection of human CD4(+) T cells identifies PSGL-1 as an HIV restriction factor. Nat. Microbiol. 2019, 4, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, Y.; Zhang, S.; Diao, M.; Huang, S.; Li, S.; Tan, X. PSGL-1 inhibits HIV-1 infection by restricting actin dynamics and sequestering HIV envelope proteins. Cell Discov. 2020, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Burnie, J.; Persaud, A.T.; Thaya, L.; Liu, Q.; Miao, H.; Grabinsky, S.; Norouzi, V.; Lusso, P.; Tang, V.A.; Guzzo, C. P-selectin glycoprotein ligand-1 (PSGL-1/CD162) is incorporated into clinical HIV-1 isolates and can mediate virus capture and subsequent transfer to permissive cells. Retrovirology 2022, 19, 9. [Google Scholar] [CrossRef]
- Lévesque, J.P.; Zannettino, A.C.; Pudney, M.; Niutta, S.; Haylock, D.N.; Snapp, K.R.; Kansas, G.S.; Berndt, M.C.; Simmons, P.J. PSGL-1-mediated adhesion of human hematopoietic progenitors to P-selectin results in suppression of hematopoiesis. Immunity 1999, 11, 369–378. [Google Scholar] [CrossRef]
- Hope, J.L.; Otero, D.C.; Bae, E.A.; Stairiker, C.J.; Palete, A.B.; Faso, H.A.; Lin, M.; Henriquez, M.L.; Roy, S.; Seo, H.; et al. PSGL-1 attenuates early TCR signaling to suppress CD8+ T cell progenitor differentiation and elicit terminal CD8+ T cell exhaustion. Cell Rep. 2023, 42, 112436. [Google Scholar] [CrossRef]
- Hong, F.F.; Mellors, J.W. Changes in HIV reservoirs during long-term antiretroviral therapy. Curr. Opin. HIV AIDS 2015, 10, 43–48. [Google Scholar] [CrossRef]
- Lee, S.A.; Deeks, S.G. The Benefits of Early Antiretroviral Therapy for HIV Infection: How Early is Early Enough? EBioMedicine 2016, 11, 7–8. [Google Scholar] [CrossRef]
- De Paula, H.H.S.; Ferreira, A.C.G.; Caetano, D.G.; Delatorre, E.; Teixeira, S.L.M.; Coelho, L.E.; João, E.G.; de Andrade, M.M.; Cardoso, S.W.; Grinsztejn, B.; et al. Reduction of inflammation and T cell activation after 6 months of cART initiation during acute, but not in early chronic HIV-1 infection. Retrovirology 2018, 15, 76. [Google Scholar] [CrossRef]
- Boulougoura, A.; Sereti, I. HIV infection and immune activation: The role of coinfections. Curr. Opin. HIV AIDS 2016, 11, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Devi, P.; Khan, A.; Chattopadhyay, P.; Mehta, P.; Sahni, S.; Sharma, S.; Pandey, R. Co-infections as Modulators of Disease Outcome: Minor Players or Major Players? Front. Microbiol. 2021, 12, 664386. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Tracy, R.; Douek, D.C. Systemic effects of inflammation on health during chronic HIV infection. Immunity 2013, 39, 633–645. [Google Scholar] [CrossRef]
- Connor, R.; Jones, L.D.; Qiu, X.; Thakar, J.; Maggirwar, S.B. Frontline Science: C-Myc regulates P-selectin glycoprotein ligand-1 expression in monocytes during HIV-1 infection. J. Leukoc. Biol. 2017, 102, 953–964. [Google Scholar] [CrossRef]
- Liang, H.; Duan, Z.; Li, D.; Li, D.; Wang, Z.; Ren, L.; Shen, T.; Shao, Y. Higher levels of circulating monocyte-platelet aggregates are correlated with viremia and increased sCD163 levels in HIV-1 infection. Cell Mol. Immunol. 2015, 12, 435–443. [Google Scholar] [CrossRef]
- Harris, R.S.; Hultquist, J.F.; Evans, D.T. The restriction factors of human immunodeficiency virus. J. Biol. Chem. 2012, 287, 40875–40883. [Google Scholar] [CrossRef] [PubMed]
- Daugherty, M.D.; Malik, H.S. Rules of engagement: Molecular insights from host-virus arms races. Annu. Rev. Genet. 2012, 46, 677–700. [Google Scholar] [CrossRef]
- Antinori, A.; Cicalini, S.; Meschi, S.; Bordoni, V.; Lorenzini, P.; Vergori, A.; Lanini, S.; De Pascale, L.; Matusali, G.; Mariotti, D.; et al. Humoral and Cellular Immune Response Elicited by mRNA Vaccination Against Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in People Living With Human Immunodeficiency Virus Receiving Antiretroviral Therapy Based on Current CD4 T-Lymphocyte Count. Clin. Infect. Dis. 2022, 75, e552–e563. [Google Scholar] [CrossRef]
- Damtie, D.; Yismaw, G.; Woldeyohannes, D.; Anagaw, B. Common opportunistic infections and their CD4 cell correlates among HIV-infected patients attending at antiretroviral therapy clinic of Gondar University Hospital, Northwest Ethiopia. BMC Res. Notes 2013, 6, 534. [Google Scholar] [CrossRef]
- Woldegeorgis, B.Z.; Zekarias, Z.; Adem, B.G.; Obsa, M.S.; Kerbo, A.A. Prevalence and determinants of opportunistic infections among HIV-infected adults receiving antiretroviral therapy in Ethiopia: A systematic review and meta-analysis. Front. Med. 2023, 10, 1087086. [Google Scholar] [CrossRef]
- Fox, M.P.; Sanne, I.M.; Conradie, F.; Zeinecker, J.; Orrell, C.; Ive, P.; Rassool, M.; Dehlinger, M.; van der Horst, C.; McIntyre, J.; et al. Initiating patients on antiretroviral therapy at CD4 cell counts above 200 cells/microl is associated with improved treatment outcomes in South Africa. Aids 2010, 24, 2041–2050. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Zhang, W.; Zhang, X.; Jiang, W.; Huan, X.; Meng, X.; Zhu, B.; Yang, Y.; Tao, Y.; Tian, T.; et al. HIV-1 CRF01_AE strain is associated with faster HIV/AIDS progression in Jiangsu Province, China. Sci. Rep. 2017, 7, 1570. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, Y.; Xie, J.; Gu, L.; Li, W.; Wang, H.; Lv, W.; Song, X.; Li, Y.; Routy, J.P.; et al. CRF01_AE subtype is associated with X4 tropism and fast HIV progression in Chinese patients infected through sexual transmission. Aids 2014, 28, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.Y.; Chew, K.K.; Kaur, P.; Kwan, J.Y.; Khong, W.X.; Lin, L.; Chua, A.; Tan, M.T.; Quinn, T.C.; Laeyendecker, O.; et al. High prevalence of CXCR4 usage among treatment-naive CRF01_AE and CRF51_01B-infected HIV-1 subjects in Singapore. BMC Infect. Dis. 2013, 13, 90. [Google Scholar] [CrossRef]
- Li, X.; Xue, Y.; Zhou, L.; Lin, Y.; Yu, X.; Wang, X.; Zhen, X.; Zhang, W.; Ning, Z.; Yue, Q.; et al. Evidence that HIV-1 CRF01_AE is associated with low CD4+T cell count and CXCR4 co-receptor usage in recently infected young men who have sex with men (MSM) in Shanghai, China. PLoS ONE 2014, 9, e89462. [Google Scholar] [CrossRef]
- Almulki, L.; Noda, K.; Amini, R.; Schering, A.; Garland, R.C.; Nakao, S.; Nakazawa, T.; Hisatomi, T.; Thomas, K.L.; Masli, S.; et al. Surprising up-regulation of P-selectin glycoprotein ligand-1 (PSGL-1) in endotoxin-induced uveitis. FASEB J. 2009, 23, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Somers, W.S.; Tang, J.; Shaw, G.D.; Camphausen, R.T. Insights into the molecular basis of leukocyte tethering and rolling revealed by structures of P- and E-selectin bound to SLeX and PSGL-1. Cell 2000, 103, 467–479. [Google Scholar] [CrossRef]
- Xu, H.; Manivannan, A.; Jiang, H.R.; Liversidge, J.; Sharp, P.F.; Forrester, J.V.; Crane, I.J. Recruitment of IFN-gamma-producing (Th1-like) cells into the inflamed retina in vivo is preferentially regulated by P-selectin glycoprotein ligand 1:P/E-selectin interactions. J. Immunol. 2004, 172, 3215–3224. [Google Scholar] [CrossRef]
- Schumacher, A.; Liebers, U.; John, M.; Gerl, V.; Meyer, M.; Witt, C.; Wolff, G. P-selectin glycoprotein ligand-1 (PSGL-1) is up-regulated on leucocytes from patients with chronic obstructive pulmonary disease. Clin. Exp. Immunol. 2005, 142, 370–376. [Google Scholar] [CrossRef]
- Lewis, D.E.; Couturier, J.P. Chapter 6—Chronic Inflammation in HIV Pathogenesis: Effects on Immune Cells, Organ Systems, and Systemic Consequences. In Translational Inflammation; Actor, J.K., Smith, K.C., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 111–131. [Google Scholar]
- van Wijk, F.; Cheroutre, H. Mucosal T cells in gut homeostasis and inflammation. Expert. Rev. Clin. Immunol. 2010, 6, 559–566. [Google Scholar] [CrossRef]
- Vidya Vijayan, K.K.; Karthigeyan, K.P.; Tripathi, S.P.; Hanna, L.E. Pathophysiology of CD4+ T-Cell Depletion in HIV-1 and HIV-2 Infections. Front. Immunol. 2017, 8, 580. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Xu, K.; Xiao, Q.; Tuo, L.; Luo, T.; Wang, S.; Yang, R.; Zhang, F.; Yang, X. Cellular and molecular insights into incomplete immune recovery in HIV/AIDS patients. Front. Immunol. 2023, 14, 1152951. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Y.; Huang, H.H.; Zhen, C.; Chen, S.Y.; Song, B.; Cao, W.J.; Shen, L.L.; Zhou, M.J.; Zhang, X.C.; Xu, R.; et al. Distinct inflammation-related proteins associated with T cell immune recovery during chronic HIV-1 infection. Emerg. Microbes Infect. 2023, 12, 2150566. [Google Scholar] [CrossRef]
- Kelly, C.; Gaskell, K.M.; Richardson, M.; Klein, N.; Garner, P.; MacPherson, P. Discordant Immune Response with Antiretroviral Therapy in HIV-1: A Systematic Review of Clinical Outcomes. PLoS ONE 2016, 11, e0156099. [Google Scholar] [CrossRef]
- Lapadula, G.; Cozzi-Lepri, A.; Marchetti, G.; Antinori, A.; Chiodera, A.; Nicastri, E.; Parruti, G.; Galli, M.; Gori, A.; Monforte, A. Risk of clinical progression among patients with immunological nonresponse despite virological suppression after combination antiretroviral treatment. Aids 2013, 27, 769–779. [Google Scholar] [CrossRef]
- Lichtenstein, K.A.; Armon, C.; Buchacz, K.; Chmiel, J.S.; Buckner, K.; Tedaldi, E.M.; Wood, K.; Holmberg, S.D.; Brooks, J.T. Low CD4+ T cell count is a risk factor for cardiovascular disease events in the HIV outpatient study. Clin. Infect. Dis. 2010, 51, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Lederman, M.M.; Calabrese, L.; Funderburg, N.T.; Clagett, B.; Medvik, K.; Bonilla, H.; Gripshover, B.; Salata, R.A.; Taege, A.; Lisgaris, M.; et al. Immunologic failure despite suppressive antiretroviral therapy is related to activation and turnover of memory CD4 cells. J. Infect. Dis. 2011, 204, 1217–1226. [Google Scholar] [CrossRef]
- Piconi, S.; Trabattoni, D.; Gori, A.; Parisotto, S.; Magni, C.; Meraviglia, P.; Bandera, A.; Capetti, A.; Rizzardini, G.; Clerici, M. Immune activation, apoptosis, and Treg activity are associated with persistently reduced CD4+ T-cell counts during antiretroviral therapy. Aids 2010, 24, 1991–2000. [Google Scholar] [CrossRef]
- Ramirez, C.M.; Sinclair, E.; Epling, L.; Lee, S.A.; Jain, V.; Hsue, P.Y.; Hatano, H.; Conn, D.; Hecht, F.M.; Martin, J.N.; et al. Immunologic profiles distinguish aviremic HIV-infected adults. Aids 2016, 30, 1553–1562. [Google Scholar] [CrossRef]
- Stiksrud, B.; Lorvik, K.B.; Kvale, D.; Mollnes, T.E.; Ueland, P.M.; Trøseid, M.; Taskén, K.; Dyrhol-Riise, A.M. Plasma IP-10 Is Increased in Immunological NonResponders and Associated With Activated Regulatory T Cells and Persisting Low CD4 Counts. J. Acquir. Immune Defic. Syndr. 2016, 73, 138–148. [Google Scholar] [CrossRef]
- Van Lelyveld, S.F.; Gras, L.; Kesselring, A.; Zhang, S.; De Wolf, F.; Wensing, A.M.; Hoepelman, A.I. Long-term complications in patients with poor immunological recovery despite virological successful HAART in Dutch ATHENA cohort. Aids 2012, 26, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, G.R.; Furrer, H.; Ledergerber, B.; Perrin, L.; Opravil, M.; Vernazza, P.; Cavassini, M.; Bernasconi, E.; Rickenbach, M.; Hirschel, B.; et al. Characteristics, determinants, and clinical relevance of CD4 T cell recovery to <500 cells/microL in HIV type 1-infected individuals receiving potent antiretroviral therapy. Clin. Infect. Dis. 2005, 41, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Lewden, C.; Chene, G.; Morlat, P.; Raffi, F.; Dupon, M.; Dellamonica, P.; Pellegrin, J.L.; Katlama, C.; Dabis, F.; Leport, C. HIV-infected adults with a CD4 cell count greater than 500 cells/mm3 on long-term combination antiretroviral therapy reach same mortality rates as the general population. J. Acquir. Immune Defic. Syndr. 2007, 46, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Kroeze, S.; Ondoa, P.; Kityo, C.M.; Siwale, M.; Akanmu, S.; Wellington, M.; de Jager, M.; Ive, P.; Mandaliya, K.; Stevens, W.; et al. Suboptimal immune recovery during antiretroviral therapy with sustained HIV suppression in sub-Saharan Africa. Aids 2018, 32, 1043–1051. [Google Scholar] [CrossRef]
- Zaongo, S.D.; Chen, Y. Metformin may be a viable adjunctive therapeutic option to potentially enhance immune reconstitution in HIV-positive immunological non-responders. Chin. Med. J. 2023, 136, 2147–2155. [Google Scholar] [CrossRef]
- Dai, X.P.; Wu, F.Y.; Cui, C.; Liao, X.J.; Jiao, Y.M.; Zhang, C.; Song, J.W.; Fan, X.; Zhang, J.Y.; He, Q.; et al. Increased Platelet-CD4+ T Cell Aggregates Are Correlated With HIV-1 Permissiveness and CD4+ T Cell Loss. Front. Immunol. 2021, 12, 799124. [Google Scholar] [CrossRef]
- Zarbock, A.; Lowell, C.A.; Ley, K. Spleen tyrosine kinase Syk is necessary for E-selectin-induced alpha(L)beta(2) integrin-mediated rolling on intercellular adhesion molecule-1. Immunity 2007, 26, 773–783. [Google Scholar] [CrossRef]
- Chen, S.C.; Huang, C.C.; Chien, C.L.; Jeng, C.J.; Su, H.T.; Chiang, E.; Liu, M.R.; Wu, C.H.; Chang, C.N.; Lin, R.H. Cross-linking of P-selectin glycoprotein ligand-1 induces death of activated T cells. Blood 2004, 104, 3233–3242. [Google Scholar] [CrossRef]
- Carlow, D.A.; Gossens, K.; Naus, S.; Veerman, K.M.; Seo, W.; Ziltener, H.J. PSGL-1 function in immunity and steady state homeostasis. Immunol. Rev. 2009, 230, 75–96. [Google Scholar] [CrossRef]
- Zevin, A.S.; McKinnon, L.; Burgener, A.; Klatt, N.R. Microbial translocation and microbiome dysbiosis in HIV-associated immune activation. Curr. Opin. HIV AIDS 2016, 11, 182–190. [Google Scholar] [CrossRef]
- Konrad, S.; Skinner, S.; Kazadi, G.B.; Gartner, K.; Lim, H.J. HIV disease progression to CD4 count <200 cells/μL and death in Saskatoon, Saskatchewan. Can. J. Infect. Dis. Med. Microbiol. 2013, 24, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Sim, J. The laboratory diagnosis of HIV infection. S. Afr. Med. J. 2000, 90, 105–109. [Google Scholar] [PubMed]
- McCune, J.M. The dynamics of CD4+ T-cell depletion in HIV disease. Nature 2001, 410, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.; García, M.; Petrovas, C.; Yamamoto, T.; Koup, R.A.; Nabel, G.J. HIV-1 causes CD4 cell death through DNA-dependent protein kinase during viral integration. Nature 2013, 498, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Doitsh, G.; Cavrois, M.; Lassen, K.G.; Zepeda, O.; Yang, Z.; Santiago, M.L.; Hebbeler, A.M.; Greene, W.C. Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue. Cell 2010, 143, 789–801. [Google Scholar] [CrossRef]
- Chávez-Galán, L.; Arenas-Del Angel, M.C.; Zenteno, E.; Chávez, R.; Lascurain, R. Cell death mechanisms induced by cytotoxic lymphocytes. Cell Mol. Immunol. 2009, 6, 15–25. [Google Scholar] [CrossRef]
- Liu, Y.; McNevin, J.P.; Holte, S.; McElrath, M.J.; Mullins, J.I. Dynamics of viral evolution and CTL responses in HIV-1 infection. PLoS ONE 2011, 6, e15639. [Google Scholar] [CrossRef]
- Reuter, M.A.; Pombo, C.; Betts, M.R. Cytokine production and dysregulation in HIV pathogenesis: Lessons for development of therapeutics and vaccines. Cytokine Growth Factor. Rev. 2012, 23, 181–191. [Google Scholar] [CrossRef]
- Lackner, A.A.; Mohan, M.; Veazey, R.S. The gastrointestinal tract and AIDS pathogenesis. Gastroenterology 2009, 136, 1965–1978. [Google Scholar] [CrossRef]
- Luo, Z.; Health, S.L.; Li, M.; Yang, H.; Wu, Y.; Collins, M.; Deeks, S.G.; Martin, J.N.; Scott, A.; Jiang, W. Variation in blood microbial lipopolysaccharide (LPS) contributes to immune reconstitution in response to suppressive antiretroviral therapy in HIV. EBioMedicine 2022, 80, 104037. [Google Scholar] [CrossRef]
- Mehraj, V.; Ramendra, R.; Isnard, S.; Dupuy, F.P.; Ponte, R.; Chen, J.; Kema, I.; Jenabian, M.A.; Costinuik, C.T.; Lebouché, B.; et al. Circulating (1→3)-β-D-glucan Is Associated with Immune Activation During Human Immunodeficiency Virus Infection. Clin. Infect. Dis. 2020, 70, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Dunham, R.M.; Vujkovic-Cvijin, I.; Yukl, S.A.; Broadhurst, M.J.; Loke, P.; Albright, R.G.; Wong, J.K.; Lederman, M.M.; Somsouk, M.; Hunt, P.W.; et al. Discordance between peripheral and colonic markers of inflammation during suppressive ART. J. Acquir. Immune Defic. Syndr. 2014, 65, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Appay, V.; Sauce, D. Immune activation and inflammation in HIV-1 infection: Causes and consequences. J. Pathol. 2008, 214, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhou, T.; Zhang, Y.; Luo, S.; Chen, H.; Chen, D.; Li, C.; Li, W. The reservoir of latent HIV. Front. Cell Infect. Microbiol. 2022, 12, 945956. [Google Scholar] [CrossRef]
- Astorga-Gamaza, A.; Grau-Expósito, J.; Burgos, J.; Navarro, J.; Curran, A.; Planas, B.; Suanzes, P.; Falcó, V.; Genescà, M.; Buzon, M.J. Identification of HIV-reservoir cells with reduced susceptibility to antibody-dependent immune response. Elife 2022, 11, e78294. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, C.; Joo, V.; Jacquier, P.; Noto, A.; Banga, R.; Perreau, M.; Pantaleo, G. T-cell exhaustion in HIV infection. Immunol. Rev. 2019, 292, 149–163. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Q.; Hui, H.; Agrawal, K.; Karris, M.A.Y.; Rana, T.M. An atlas of immune cell exhaustion in HIV-infected individuals revealed by single-cell transcriptomics. Emerg. Microbes Infect. 2020, 9, 2333–2347. [Google Scholar] [CrossRef]
- Nguyen, S.; Deleage, C.; Darko, S.; Ransier, A.; Truong, D.P.; Agarwal, D.; Japp, A.S.; Wu, V.H.; Kuri-Cervantes, L.; Abdel-Mohsen, M.; et al. Elite control of HIV is associated with distinct functional and transcriptional signatures in lymphoid tissue CD8(+) T cells. Sci. Transl. Med. 2019, 11, eaax4077. [Google Scholar] [CrossRef]
- Gubin, M.M.; Zhang, X.; Schuster, H.; Caron, E.; Ward, J.P.; Noguchi, T.; Ivanova, Y.; Hundal, J.; Arthur, C.D.; Krebber, W.J.; et al. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature 2014, 515, 577–581. [Google Scholar] [CrossRef]
- DeRogatis, J.M.; Viramontes, K.M.; Neubert, E.N.; Tinoco, R. PSGL-1 Immune Checkpoint Inhibition for CD4+ T Cell Cancer Immunotherapy. Front. Immunol. 2021, 12, 636238. [Google Scholar] [CrossRef]
- Tinoco, R.; Carrette, F.; Barraza, M.L.; Otero, D.C.; Magaña, J.; Bosenberg, M.W.; Swain, S.L.; Bradley, L.M. PSGL-1 Is an Immune Checkpoint Regulator that Promotes T Cell Exhaustion. Immunity 2016, 44, 1190–1203. [Google Scholar] [CrossRef] [PubMed]
- Tinoco, R.; Neubert, E.N.; Stairiker, C.J.; Henriquez, M.L.; Bradley, L.M. PSGL-1 Is a T Cell Intrinsic Inhibitor That Regulates Effector and Memory Differentiation and Responses During Viral Infection. Front. Immunol. 2021, 12, 677824. [Google Scholar] [CrossRef] [PubMed]
- Wallace, Z.; Kopycinski, J.; Yang, H.; McCully, M.L.; Eggeling, C.; Chojnacki, J.; Dorrell, L. Immune mobilising T cell receptors redirect polyclonal CD8(+) T cells in chronic HIV infection to form immunological synapses. Sci. Rep. 2022, 12, 18366. [Google Scholar] [CrossRef] [PubMed]
- Schaub, B.; Bellou, A.; Gibbons, F.K.; Velasco, G.; Campo, M.; He, H.; Liang, Y.; Gillman, M.W.; Gold, D.; Weiss, S.T.; et al. TLR2 and TLR4 stimulation differentially induce cytokine secretion in human neonatal, adult, and murine mononuclear cells. J. Interferon. Cytokine Res. 2004, 24, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Girard, A.; Vergnon-Miszczycha, D.; Depincé-Berger, A.E.; Roblin, X.; Lutch, F.; Lambert, C.; Rochereau, N.; Bourlet, T.; Genin, C.; Paul, S. Brief Report: A High Rate of β7+ Gut-Homing Lymphocytes in HIV-Infected Immunological Nonresponders is Associated With Poor CD4 T-Cell Recovery During Suppressive HAART. J. Acquir. Immune Defic. Syndr. 2016, 72, 259–265. [Google Scholar] [CrossRef]
- Norris, P.J.; Zhang, J.; Worlock, A.; Nair, S.V.; Anastos, K.; Minkoff, H.L.; Villacres, M.C.; Young, M.; Greenblatt, R.M.; Desai, S.; et al. Systemic Cytokine Levels Do Not Predict CD4+ T-Cell Recovery After Suppressive Combination Antiretroviral Therapy in Chronic Human Immunodeficiency Virus Infection. Open Forum Infect. Dis. 2016, 3, ofw025. [Google Scholar] [CrossRef]
- Massanella, M.; Gómez-Mora, E.; Carrillo, J.; Curriu, M.; Ouchi, D.; Puig, J.; Negredo, E.; Cabrera, C.; Clotet, B.; Blanco, J. Increased ex vivo cell death of central memory CD4 T cells in treated HIV infected individuals with unsatisfactory immune recovery. J. Transl. Med. 2015, 13, 230. [Google Scholar] [CrossRef]
- Gómez-Mora, E.; García, E.; Urrea, V.; Massanella, M.; Puig, J.; Negredo, E.; Clotet, B.; Blanco, J.; Cabrera, C. Preserved immune functionality and high CMV-specific T-cell responses in HIV-infected individuals with poor CD4+ T-cell immune recovery. Sci. Rep. 2017, 7, 11711. [Google Scholar] [CrossRef]
- Woelk, C.H.; Beliakova-Bethell, N.; Goicoechea, M.; Zhao, Y.; Du, P.; Rought, S.E.; Lozach, J.; Pérez-Santiago, J.; Richman, D.D.; Smith, D.M.; et al. Gene expression before HAART initiation predicts HIV-infected individuals at risk of poor CD4+ T-cell recovery. Aids 2010, 24, 217–222. [Google Scholar] [CrossRef]
- Goicoechea, M.; Smith, D.M.; Liu, L.; May, S.; Tenorio, A.R.; Ignacio, C.C.; Landay, A.; Haubrich, R. Determinants of CD4+ T cell recovery during suppressive antiretroviral therapy: Association of immune activation, T cell maturation markers, and cellular HIV-1 DNA. J. Infect. Dis. 2006, 194, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Shete, A.; Dhayarkar, S.; Sangale, S.; Medhe, U.; Panchal, N.; Rahane, G.; Yelgate, R.; Dhamanage, A.; Gangakhedkar, R. Incomplete functional T-cell reconstitution in immunological non-responders at one year after initiation of antiretroviral therapy possibly predisposes them to infectious diseases. Int. J. Infect. Dis. 2019, 81, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Lederman, M.M.; Funderburg, N.T.; Sekaly, R.P.; Klatt, N.R.; Hunt, P.W. Residual immune dysregulation syndrome in treated HIV infection. Adv. Immunol. 2013, 119, 51–83. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ramírez, R.U.; Shiels, M.S.; Dubrow, R.; Engels, E.A. Cancer risk in HIV-infected people in the USA from 1996 to 2012: A population-based, registry-linkage study. Lancet HIV 2017, 4, e495–e504. [Google Scholar] [CrossRef]
- Shiels, M.S.; Cole, S.R.; Kirk, G.D.; Poole, C. A meta-analysis of the incidence of non-AIDS cancers in HIV-infected individuals. J. Acquir. Immune Defic. Syndr. 2009, 52, 611–622. [Google Scholar] [CrossRef]
- Dubrow, R.; Silverberg, M.J.; Park, L.S.; Crothers, K.; Justice, A.C. HIV infection, aging, and immune function: Implications for cancer risk and prevention. Curr. Opin. Oncol. 2012, 24, 506–516. [Google Scholar] [CrossRef]
- Lin, Y.; Huang, S.; Qi, Y.; Xie, L.; Jiang, J.; Li, H.; Chen, Z. PSGL-1 is a novel tumor microenvironment prognostic biomarker with cervical high-grade squamous lesions and more. Front. Oncol. 2023, 13, 1052201. [Google Scholar] [CrossRef]
- Kappelmayer, J.; Nagy, B., Jr. The Interaction of Selectins and PSGL-1 as a Key Component in Thrombus Formation and Cancer Progression. Biomed. Res. Int. 2017, 2017, 6138145. [Google Scholar] [CrossRef]
- Dimitroff, C.J.; Descheny, L.; Trujillo, N.; Kim, R.; Nguyen, V.; Huang, W.; Pienta, K.J.; Kutok, J.L.; Rubin, M.A. Identification of leukocyte E-selectin ligands, P-selectin glycoprotein ligand-1 and E-selectin ligand-1, on human metastatic prostate tumor cells. Cancer Res. 2005, 65, 5750–5760. [Google Scholar] [CrossRef]
- Heidemann, F.; Schildt, A.; Schmid, K.; Bruns, O.T.; Riecken, K.; Jung, C.; Ittrich, H.; Wicklein, D.; Reimer, R.; Fehse, B.; et al. Selectins mediate small cell lung cancer systemic metastasis. PLoS ONE 2014, 9, e92327. [Google Scholar] [CrossRef]
- Hoos, A.; Protsyuk, D.; Borsig, L. Metastatic growth progression caused by PSGL-1-mediated recruitment of monocytes to metastatic sites. Cancer Res. 2014, 74, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Presti, R.; Pantaleo, G. 92-The Immunopathogenesis of HIV-1 Infection, in Infectious Diseases, 4th ed.; Cohen, J., Powderly, W.G., Opal, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 837–845.e833. [Google Scholar]
- Hildebrandt, X.; Ibrahim, M.; Peltzer, N. Cell death and inflammation during obesity: “Know my methods, WAT(son)”. Cell Death Differ. 2023, 30, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Khanna, D.; Khanna, S.; Khanna, P.; Kahar, P.; Patel, B.M. Obesity: A Chronic Low-Grade Inflammation and Its Markers. Cureus 2022, 14, e22711. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Sato, C.; Shikata, K.; Hirota, D.; Sasaki, M.; Nishishita, S.; Miyamoto, S.; Kodera, R.; Ogawa, D.; Tone, A.; Kataoka, H.U.; et al. P-selectin glycoprotein ligand-1 deficiency is protective against obesity-related insulin resistance. Diabetes 2011, 60, 189–199. [Google Scholar] [CrossRef][Green Version]
- Global HIV & AIDS Statistics—Fact Sheet|UNAIDS. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 23 October 2023).
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and aging: Signaling pathways and intervention therapies. Signal Transduct. Target. Ther. 2023, 8, 239. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaongo, S.D.; Chen, Y. PSGL-1, a Strategic Biomarker for Pathological Conditions in HIV Infection: A Hypothesis Review. Viruses 2023, 15, 2197. https://doi.org/10.3390/v15112197
Zaongo SD, Chen Y. PSGL-1, a Strategic Biomarker for Pathological Conditions in HIV Infection: A Hypothesis Review. Viruses. 2023; 15(11):2197. https://doi.org/10.3390/v15112197
Chicago/Turabian StyleZaongo, Silvere D., and Yaokai Chen. 2023. "PSGL-1, a Strategic Biomarker for Pathological Conditions in HIV Infection: A Hypothesis Review" Viruses 15, no. 11: 2197. https://doi.org/10.3390/v15112197
APA StyleZaongo, S. D., & Chen, Y. (2023). PSGL-1, a Strategic Biomarker for Pathological Conditions in HIV Infection: A Hypothesis Review. Viruses, 15(11), 2197. https://doi.org/10.3390/v15112197