
Growth, Pathogenesis, and Serological Characteristics of the Japanese Encephalitis Virus Genotype IV Recent Strain 19CxBa-83-Cv
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Cell Culture
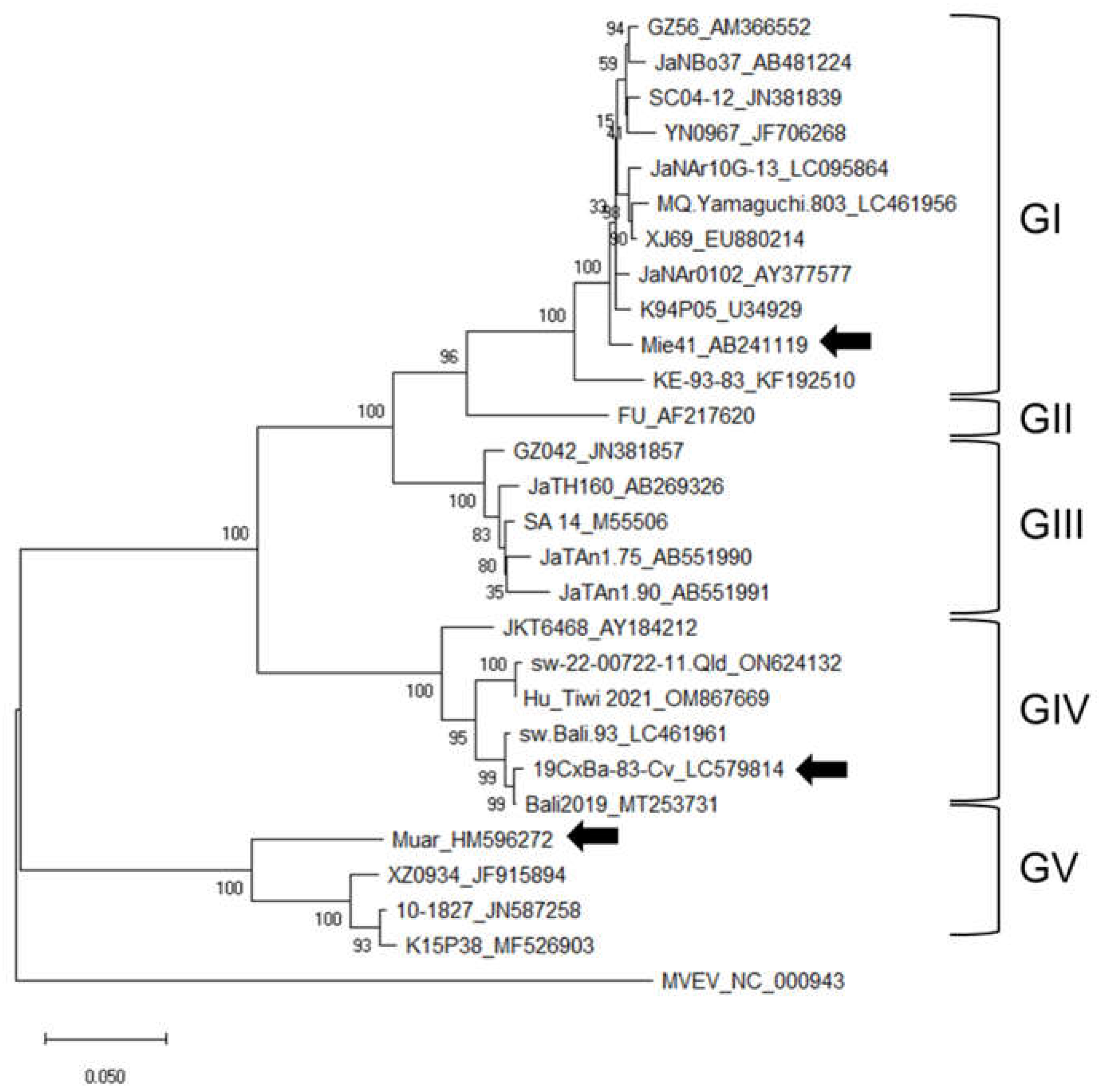
2.3. Phylogenetic Analysis
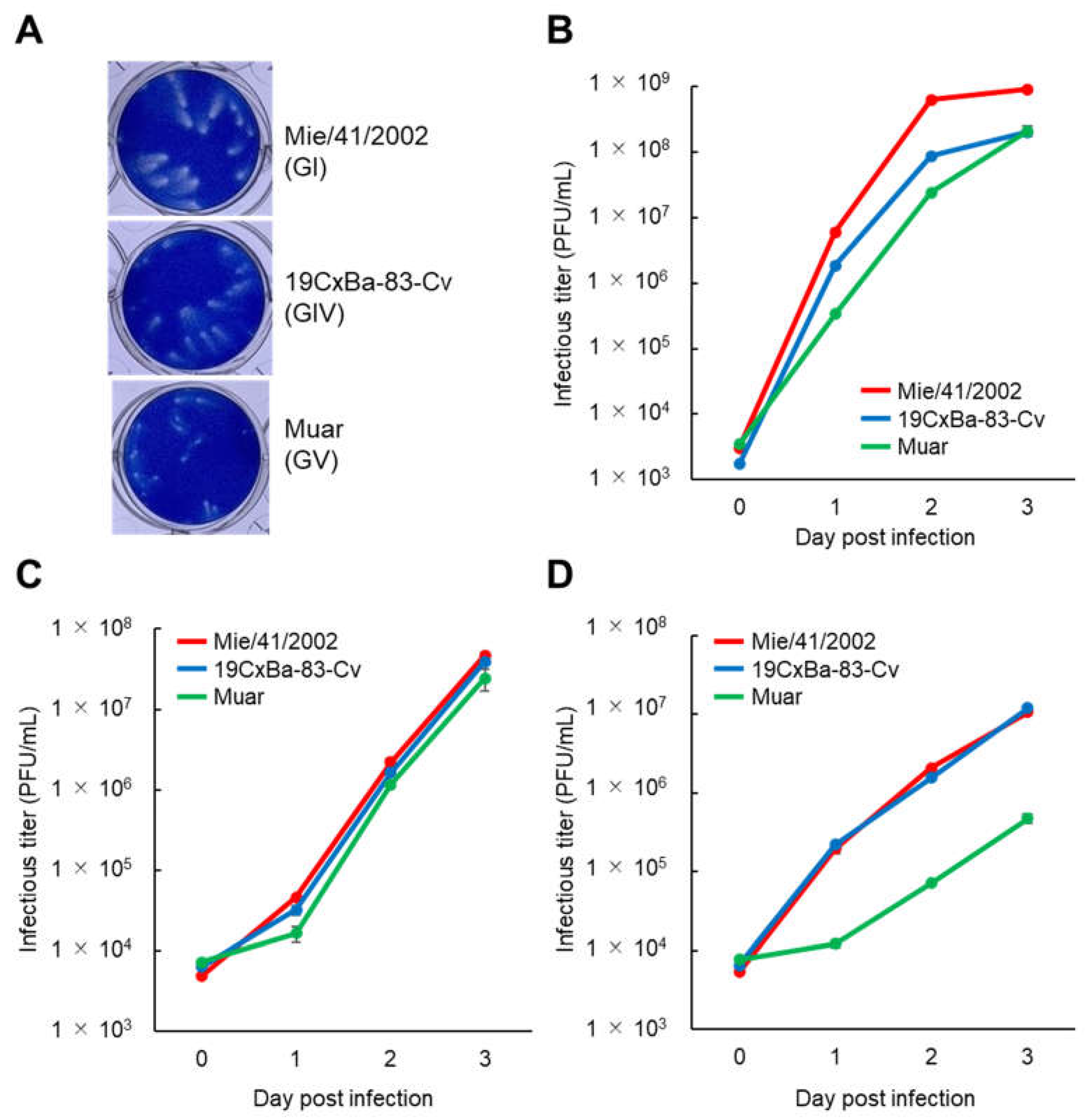
2.4. Plaque Formation Assay for Titration of Infectious Viruses and Analysis of Growth Kinetics
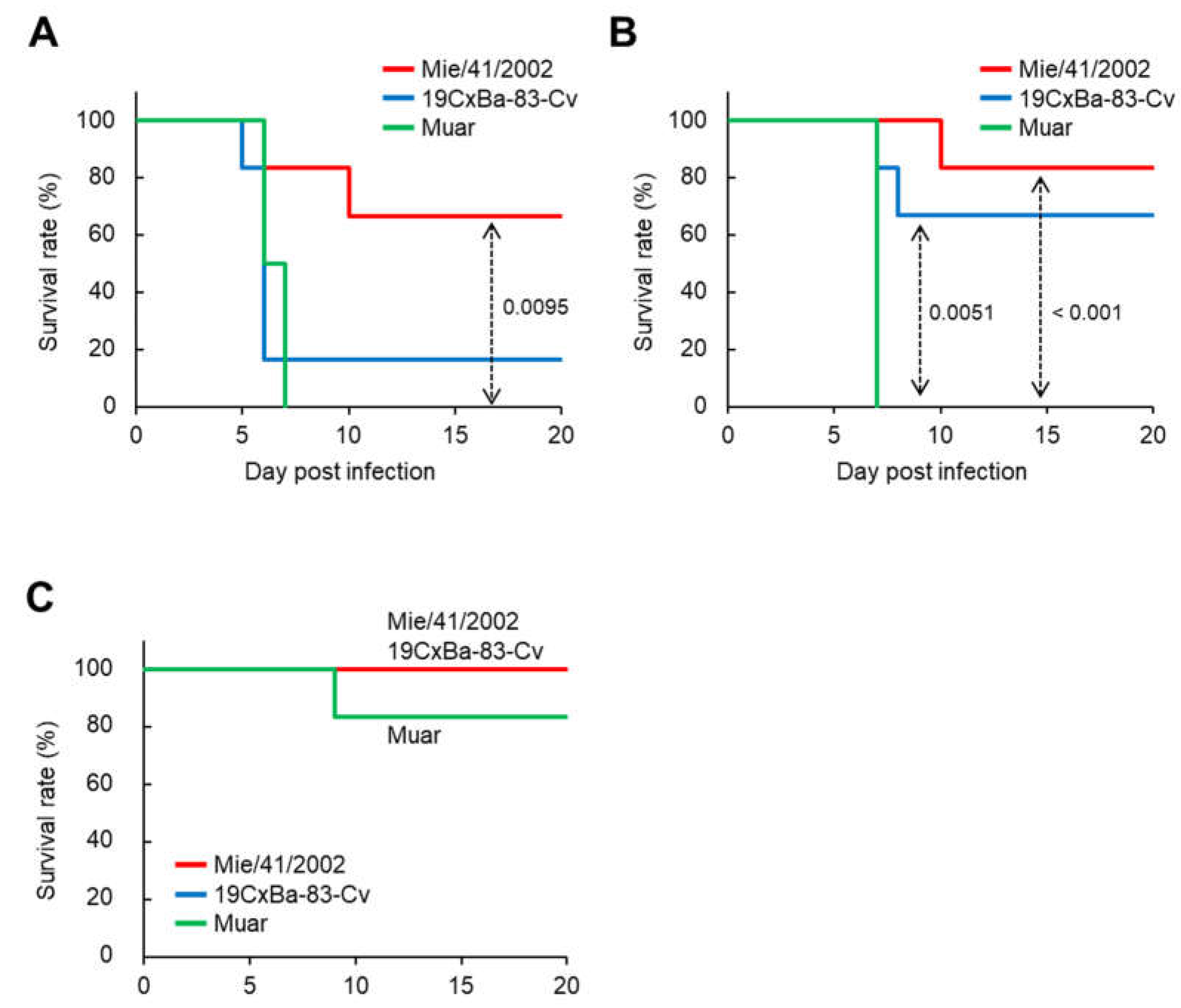
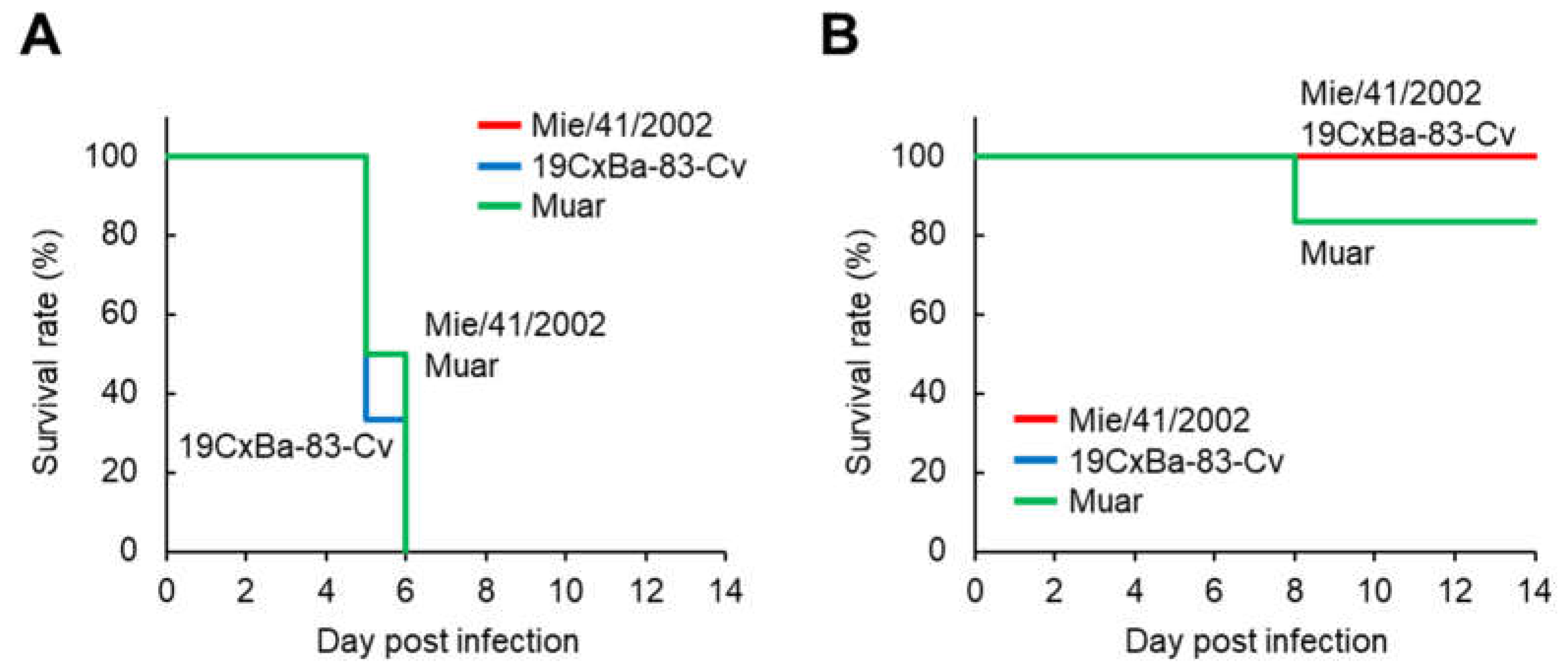
2.5. Mouse Challenge Experiment
2.6. Mouse Vaccination
2.7. Sera of JE Patients
2.8. Plaque Reduction Neutralization Test (PRNT)
3. Results
3.1. Phylogenetic Analysis of the 19CxBa-83-Cv Strain
3.2. Growth Properties of the 19CxBa-83-Cv Strain In Vitro
3.3. Virulence of the 19CxBa-83-Cv Strain in Mice
3.4. Neutralizing Ability of Sera from the 19CxBa-83-Cv-Infected Mice
3.5. Neutralizing Ability of Pooled Serum from Mice Vaccinated with Vero Cell-Derived Inactivated JE Vaccine against the 19CxBa-83-Cv
3.6. Neutralizing Ability of Sera from JE Patients against 19CxBa-83-Cv
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Erlanger, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, present, and future of Japanese encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar] [CrossRef]
- Tsai, T.F. New initiatives for the control of Japanese encephalitis by vaccination: Minutes of a WHO/CVI meeting, Bangkok, Thailand, 13–15 October 1998. Vaccine 2000, 18 (Suppl. 2), 1–25. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.D.; Flaviviruses, M.S. Fields Virology, 6th ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 747–794. [Google Scholar]
- Solomon, T.; Ni, H.; Beasley, D.W.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D. Origin and evolution of Japanese encephalitis virus in southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef]
- Nam, J.H.; Chung, Y.J.; Ban, S.J.; Kim, E.J.; Park, Y.K.; Cho, H.W. Envelope gene sequence variation among Japanese encephalitis viruses isolated in Korea. Acta Virol. 1996, 40, 303–309. [Google Scholar]
- Yang, D.K.; Kim, B.H.; Kweon, C.H.; Kwon, J.H.; Lim, S.I.; Han, H.R. Molecular characterization of full-length genome of Japanese encephalitis virus (KV1899) isolated from pigs in Korea. J. Vet. Sci. 2004, 5, 197–205. [Google Scholar] [CrossRef]
- Nga, P.T.; del Carmen Parquet, M.; Cuong, V.D.; Ma, S.P.; Hasebe, F.; Inoue, S.; Makino, Y.; Takagi, M.; Nam, V.S.; Morita, K. Shift in Japanese encephalitis virus (JEV) genotype circulating in northern Vietnam: Implications for frequent introductions of JEV from Southeast Asia to East Asia. J. Gen. Virol. 2004, 85, 1625–1631. [Google Scholar] [CrossRef]
- Wang, H.Y.; Takasaki, T.; Fu, S.H.; Sun, X.H.; Zhang, H.L.; Wang, Z.X.; Hao, Z.Y.; Zhang, J.K.; Tang, Q.; Kotaki, A.; et al. Molecular epidemiological analysis of Japanese encephalitis virus in China. J. Gen. Virol. 2007, 88, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Nitatpattana, N.; Dubot-Peres, A.; Gouilh, M.A.; Souris, M.; Barbazan, P.; Yoksan, S.; de Lamballerie, X.; Gonzalez, J.P. Change in Japanese encephalitis virus distribution, Thailand. Emerg. Infect. Dis. 2008, 14, 1762–1765. [Google Scholar] [CrossRef]
- Yun, S.M.; Cho, J.E.; Ju, Y.R.; Kim, S.Y.; Ryou, J.; Han, M.G.; Choi, W.Y.; Jeong, Y.E. Molecular epidemiology of Japanese encephalitis virus circulating in South Korea, 1983–2005. Virol. J. 2010, 7, 127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Zhao, Q.M.; Zhang, P.H.; Jia, N.; Cao, W.C. Genomic sequence of a Japanese encephalitis virus isolate from southern China. Arch. Virol. 2009, 154, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Zhao, Q.M.; Guo, X.F.; Zuo, S.Q.; Cheng, J.X.; Jia, N.; Wu, C.; Dai, P.F.; Zhao, J.Y. Isolation and genetic characteristics of human genotype 1 Japanese encephalitis virus, China, 2009. PLoS ONE 2011, 6, e16418. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Fan, Y.C.; Tu, W.C.; Chang, R.Y.; Shih, C.C.; Lu, I.H.; Chien, M.S.; Lee, W.C.; Chen, T.H.; Chang, G.J.; et al. Japanese encephalitis virus genotype replacement, Taiwan, 2009–2010. Emerg. Infect. Dis. 2011, 17, 2354–2356. [Google Scholar] [CrossRef]
- Pan, X.L.; Liu, H.; Wang, H.Y.; Fu, S.H.; Liu, H.Z.; Zhang, H.L.; Li, M.H.; Gao, X.Y.; Wang, J.L.; Sun, X.H.; et al. Emergence of genotype I of Japanese encephalitis virus as the dominant genotype in Asia. J. Virol. 2011, 85, 9847–9853. [Google Scholar] [CrossRef]
- Fulmali, P.V.; Sapkal, G.N.; Athawale, S.; Gore, M.M.; Mishra, A.C.; Bondre, V.P. Introduction of Japanese encephalitis virus genotype I, India. Emerg. Infect. Dis. 2011, 17, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Schuh, A.J.; Ward, M.J.; Leigh Brown, A.J.; Barrett, A.D. Dynamics of the Emergence and Establishment of a Newly Dominant Genotype of Japanese Encephalitis Virus throughout Asia. J. Virol. 2014, 88, 4522–4532. [Google Scholar] [CrossRef]
- Hameed, M.; Wahaab, A.; Nawaz, M.; Khan, S.; Nazir, J.; Liu, K.; Wei, J.; Ma, Z. Potential Role of Birds in Japanese Encephalitis Virus Zoonotic Transmission and Genotype Shift. Viruses 2021, 13, 357. [Google Scholar] [CrossRef] [PubMed]
- Schuh, A.J.; Guzman, H.; Tesh, R.B.; Barrett, A.D. Genetic diversity of Japanese encephalitis virus isolates obtained from the Indonesian archipelago between 1974 and 1987. Vector-Borne Zoonotic Dis. 2013, 13, 479–488. [Google Scholar] [CrossRef]
- Kuwata, R.; Torii, S.; Shimoda, H.; Supriyono, S.; Phichitraslip, T.; Prasertsincharoen, N.; Takemae, H.; Bautista, R.; Ebora, V.; Abella, J.A.C.; et al. Distribution of Japanese Encephalitis Virus, Japan and Southeast Asia, 2016-2018. Emerg. Infect. Dis. 2020, 26, 125–128. [Google Scholar] [CrossRef]
- Li, M.H.; Fu, S.H.; Chen, W.X.; Wang, H.Y.; Guo, Y.H.; Liu, Q.Y.; Li, Y.X.; Luo, H.M.; Da, W.; Duo Ji, D.Z.; et al. Genotype v Japanese encephalitis virus is emerging. PLoS Negl. Trop. Dis. 2011, 5, e1231. [Google Scholar] [CrossRef]
- Takhampunya, R.; Kim, H.C.; Tippayachai, B.; Kengluecha, A.; Klein, T.A.; Lee, W.J.; Grieco, J.; Evans, B.P. Emergence of Japanese encephalitis virus genotype V in the Republic of Korea. Virol. J. 2011, 8, 449. [Google Scholar] [CrossRef]
- Kim, H.; Cha, G.W.; Jeong, Y.E.; Lee, W.G.; Chang, K.S.; Roh, J.Y.; Yang, S.C.; Park, M.Y.; Park, C.; Shin, E.H. Detection of Japanese encephalitis virus genotype V in Culex orientalis and Culex pipiens (Diptera: Culicidae) in Korea. PLoS ONE 2015, 10, e0116547. [Google Scholar] [CrossRef] [PubMed]
- Bae, W.; Kim, J.H.; Kim, J.; Lee, J.; Hwang, E.S. Changes of Epidemiological Characteristics of Japanese Encephalitis Viral Infection and Birds as a Potential Viral Transmitter in Korea. J. Korean Med. Sci. 2018, 33, e70. [Google Scholar] [CrossRef]
- de Wispelaere, M.; Frenkiel, M.P.; Despres, P. A Japanese Encephalitis virus genotype 5 molecular clone is highly neuropathogenic in a mouse model: Implication of the structural proteins region in virulence. J. Virol. 2015, 89, 5862–5875. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Fu, S.; Gao, X.; Li, M.; Cui, S.; Li, X.; Cao, Y.; Lei, W.; Lu, Z.; He, Y.; et al. Low Protective Efficacy of the Current Japanese Encephalitis Vaccine against the Emerging Genotype 5 Japanese Encephalitis Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004686. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Yagasaki, K.; Kotaki, A.; Tomikawa, T.; Nakayama, E.; Moi, M.L.; Lim, C.K.; Saijo, M.; Kurane, I.; Takasaki, T. In vitro growth, pathogenicity and serological characteristics of the Japanese encephalitis virus genotype V Muar strain. J. Gen. Virol. 2015, 96, 2661–2669. [Google Scholar] [CrossRef]
- Tajima, S.; Shibasaki, K.I.; Taniguchi, S.; Nakayama, E.; Maeki, T.; Lim, C.K.; Saijo, M. E and prM proteins of genotype V Japanese encephalitis virus are required for its increased virulence in mice. Heliyon 2019, 5, e02882. [Google Scholar] [CrossRef]
- Tajima, S.; Taniguchi, S.; Nakayama, E.; Maeki, T.; Inagaki, T.; Lim, C.K.; Saijo, M. Amino Acid at Position 166 of NS2A in Japanese Encephalitis Virus (JEV) is Associated with In Vitro Growth Characteristics of JEV. Viruses 2020, 12, 709. [Google Scholar] [CrossRef] [PubMed]
- Faizah, A.N.; Kobayashi, D.; Maekawa, Y.; Amoa-Bosompem, M.; Fauziyah, S.; Mulyatno, K.C.; Subekti, S.; Rohmah, E.A.; Lusida, M.I.; Mori, Y.; et al. Identification and Isolation of Japanese Encephalitis Virus Genotype IV from Culex vishnui Collected in Bali, Indonesia in 2019. Am. J. Trop. Med. Hyg. 2021, 105, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Pyke, A.T.; Choong, K.; Moore, F.; Schlebusch, S.; Taylor, C.; Hewitson, G.; McMahon, J.; Nair, N.; Moore, P.; Finger, M.; et al. A Case of Japanese Encephalitis with a Fatal Outcome in an Australian Who Traveled from Bali in 2019. Trop. Med. Infect. Dis. 2020, 5, 133. [Google Scholar] [CrossRef]
- Hanna, J.N.; Ritchie, S.A.; Phillips, D.A.; Shield, J.; Bailey, M.C.; Mackenzie, J.S.; Poidinger, M.; McCall, B.J.; Mills, P.J. An outbreak of Japanese encephalitis in the Torres Strait, Australia, 1995. Med. J. Aust. 1996, 165, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Phillips, D.; Broom, A.; Mackenzie, J.; Poidinger, M.; van den Hurk, A. Isolation of Japanese encephalitis virus from Culex annulirostris in Australia. Am. J. Trop. Med. Hyg. 1997, 56, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.T.; Wang, L.F.; Daniels, P.W.; Mackenzie, J.S. Molecular characterization of the first Australian isolate of Japanese encephalitis virus, the FU strain. J. Gen. Virol. 2000, 81, 2471–2480. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.N.; Ritchie, S.A.; Phillips, D.A.; Lee, J.M.; Hills, S.L.; van den Hurk, A.F.; Pyke, A.T.; Johansen, C.A.; Mackenzie, J.S. Japanese encephalitis in north Queensland, Australia, 1998. Med. J. Aust. 1999, 170, 533–536. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Pyke, A.T.; Mackenzie, J.S.; Hall-Mendelin, S.; Ritchie, S.A. Japanese Encephalitis Virus in Australia: From Known Known to Known Unknown. Trop. Med. Infect. Dis. 2019, 4, 38. [Google Scholar] [CrossRef]
- Pyke, A.T.; Williams, D.T.; Nisbet, D.J.; van den Hurk, A.F.; Taylor, C.T.; Johansen, C.A.; Macdonald, J.; Hall, R.A.; Simmons, R.J.; Mason, R.J.; et al. The appearance of a second genotype of Japanese encephalitis virus in the Australasian region. Am. J. Trop. Med. Hyg. 2001, 65, 747–753. [Google Scholar] [CrossRef]
- Waller, C.; Tiemensma, M.; Currie, B.J.; Williams, D.T.; Baird, R.W.; Krause, V.L. Japanese Encephalitis in Australia—A Sentinel Case. N. Engl. J. Med. 2022, 387, 661–662. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Williams, D.T.; van den Hurk, A.F.; Smith, D.W.; Currie, B.J. Japanese Encephalitis Virus: The Emergence of Genotype IV in Australia and Its Potential Endemicity. Viruses 2022, 14, 2480. [Google Scholar] [CrossRef]
- Sikazwe, C.; Neave, M.J.; Michie, A.; Mileto, P.; Wang, J.; Cooper, N.; Levy, A.; Imrie, A.; Baird, R.W.; Currie, B.J.; et al. Molecular detection and characterisation of the first Japanese encephalitis virus belonging to genotype IV acquired in Australia. PLoS Negl. Trop. Dis. 2022, 16, e0010754. [Google Scholar] [CrossRef]
- Australian Government Department of Health Japanese Encephalitis Virus (JEV). Available online: https://www.health.gov.au/health-alerts/japanese-encephalitis-virus-jev/japanese-encephalitis-virus-jev?language=en (accessed on 22 October 2022).
- Howard-Jones, A.R.; Pham, D.; Jeoffreys, N.; Eden, J.S.; Hueston, L.; Kesson, A.M.; Nagendra, V.; Samarasekara, H.; Newton, P.; Chen, S.C.; et al. Emerging Genotype IV Japanese Encephalitis Virus Outbreak in New South Wales, Australia. Viruses 2022, 14, 1853. [Google Scholar] [CrossRef]
- Pham, D.; Howard-Jones, A.R.; Hueston, L.; Jeoffreys, N.; Doggett, S.; Rockett, R.; Eden, J.S.; Sintchenko, V.; Chen, S.C.; O’Sullivan, M.V.; et al. Emergence of Japanese encephalitis in Australia: A diagnostic perspective. Pathology 2022, 54, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Nerome, R.; Nukui, Y.; Kato, F.; Takasaki, T.; Kurane, I. A single mutation in the Japanese encephalitis virus E protein (S123R) increases its growth rate in mouse neuroblastoma cells and its pathogenicity in mice. Virology 2010, 396, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Taniguchi, S.; Nakayama, E.; Maeki, T.; Inagaki, T.; Saijo, M.; Lim, C.K. Immunogenicity and Protective Ability of Genotype I-Based Recombinant Japanese Encephalitis Virus (JEV) with Attenuation Mutations in E Protein against Genotype V JEV. Vaccines 2021, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Maeki, T.; Tajima, S.; Ikeda, M.; Kato, F.; Taniguchi, S.; Nakayama, E.; Takasaki, T.; Lim, C.K.; Saijo, M. Analysis of cross-reactivity between flaviviruses with sera of patients with Japanese encephalitis showed the importance of neutralization tests for the diagnosis of Japanese encephalitis. J. Infect. Chemother. 2019, 25, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Nerome, R.; Tajima, S.; Takasaki, T.; Yoshida, T.; Kotaki, A.; Lim, C.K.; Ito, M.; Sugiyama, A.; Yamauchi, A.; Yano, T.; et al. Molecular epidemiological analyses of Japanese encephalitis virus isolates from swine in Japan from 2002 to 2004. J. Gen. Virol. 2007, 88, 2762–2768. [Google Scholar] [CrossRef]
- Sumiyoshi, H.; Tignor, G.H.; Shope, R.E. Characterization of a highly attenuated Japanese encephalitis virus generated from molecularly cloned cDNA. J. Infect. Dis. 1995, 171, 1144–1151. [Google Scholar] [CrossRef]
- Ni, H.; Barrett, A.D. Attenuation of Japanese encephalitis virus by selection of its mouse brain membrane receptor preparation escape variants. Virology 1998, 241, 30–36. [Google Scholar] [CrossRef]
- Monath, T.P.; Arroyo, J.; Levenbook, I.; Zhang, Z.X.; Catalan, J.; Draper, K.; Guirakhoo, F. Single mutation in the flavivirus envelope protein hinge region increases neurovirulence for mice and monkeys but decreases viscerotropism for monkeys: Relevance to development and safety testing of live, attenuated vaccines. J. Virol. 2002, 76, 1932–1943. [Google Scholar] [CrossRef]
- Zhao, Z.; Date, T.; Li, Y.; Kato, T.; Miyamoto, M.; Yasui, K.; Wakita, T. Characterization of the E-138 (Glu/Lys) mutation in Japanese encephalitis virus by using a stable, full-length, infectious cDNA clone. J. Gen. Virol. 2005, 86, 2209–2220. [Google Scholar] [CrossRef]
- Li, X.; Ma, S.J.; Liu, X.; Jiang, L.N.; Zhou, J.H.; Xiong, Y.Q.; Ding, H.; Chen, Q. Immunogenicity and safety of currently available Japanese encephalitis vaccines: A systematic review. Hum. Vaccines Immunother. 2014, 10, 3579–3593. [Google Scholar] [CrossRef]
- Monath, T.P.; Seligman, S.J.; Robertson, J.S.; Guy, B.; Hayes, E.B.; Condit, R.C.; Excler, J.L.; Mac, L.M.; Carbery, B.; Chen, R.T.; et al. Live virus vaccines based on a yellow fever vaccine backbone: Standardized template with key considerations for a risk/benefit assessment. Vaccine 2015, 33, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Satchidanandam, V. Japanese Encephalitis Vaccines. Curr. Treat. Options Infect. Dis. 2020, 12, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Erra, E.O.; Askling, H.H.; Yoksan, S.; Rombo, L.; Riutta, J.; Vene, S.; Lindquist, L.; Vapalahti, O.; Kantele, A. Cross-protective capacity of Japanese encephalitis (JE) vaccines against circulating heterologous JE virus genotypes. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2013, 56, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.P.; Yoshida, Y.; Makino, Y.; Tadano, M.; Ono, T.; Ogawa, M. Short report: A major genotype of Japanese encephalitis virus currently circulating in Japan. Am. J. Trop. Med. Hyg. 2003, 69, 151–154. [Google Scholar] [CrossRef]
- Yoshikawa, A.; Nabeshima, T.; Inoue, S.; Agoh, M.; Morita, K. Molecular and serological epidemiology of Japanese encephalitis virus (JEV) in a remote island of western Japan: An implication of JEV migration over the East China Sea. Trop. Med. Health 2016, 44, 8. [Google Scholar] [CrossRef]
- Maeki, T.; Tajima, S.; Kyaw, A.K.; Matsumoto, F.; Miura, K.; Yamashita, A.; Yoshikawa, A.; Negishi, K.; Noguchi, Y.; Tadokoro, K.; et al. Comparison of Neutralizing Antibody Titers against Japanese Encephalitis Virus Genotype V Strain with Those against Genotype I and III Strains in the Sera of Japanese Encephalitis Patients in Japan in 2016. Jpn. J. Infect. Dis. 2018, 71, 360–364. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Tajima, S.; Ikeda, M.; Nguyen, T.T.; Le, T.T.H.; Pham, H.T.T.; Pham, D.Q.; Le, M.T.Q.; Maeki, T.; Taniguchi, S.; et al. Neutralization Potency of Sera from Vietnamese Patients with Japanese Encephalitis (JE) against Genotypes I and V JE Viruses. Jpn. J. Infect. Dis. 2019, 72, 115–117. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse | Titer Inoculated | Mouse ID | PRNT50 against: | ||
|---|---|---|---|---|---|
| Mie/41/2002 (GI) | 19CxBa-83-Cv (GIV) | Muar (GV) | |||
| Mie/41/2002-inoculated group | 105 104 104 103 | PR PY PYG PW | 640 1280 640 <10 | 320 320 320 <10 | 80 80 160 <10 |
| 19CxBa-83-Cv-inoculated group | 105 104 104 104 103 | YG YW YB YG YW | 1280 320 320 160 10 | 640 160 320 320 <10 | 160 40 80 40 <10 |
| Muar-inoculated group | 103 | GW | <10 | <10 | <10 |
| Serum | PRNT50 against: | ||
|---|---|---|---|
| Mie/41/2002 (GI) | 19CxBa-83-Cv (GIV) | Muar (GV) | |
| JSS-2020 1 | 160 | 20 | 40 |
| Patient | Days after Onset | PRNT50 against: | ||
|---|---|---|---|---|
| Mie/41/2002 (GI) | 19CxBa-83-Cv (GIV) | Muar (GV) | ||
| 1 | 25 | 2560 | 2560 | 2560 |
| 2 | 7 | 160 | 80 | 80 |
| 16 | 5120 | 5120 | 5120 | |
| 3 | 10 | 640 | 160 | 160 |
| 4 | 10 | 2560 | 320 | 160 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tajima, S.; Maeki, T.; Nakayama, E.; Faizah, A.N.; Kobayashi, D.; Isawa, H.; Maekawa, Y.; Bendryman, S.S.; Mulyatno, K.C.; Rohmah, E.A.; et al. Growth, Pathogenesis, and Serological Characteristics of the Japanese Encephalitis Virus Genotype IV Recent Strain 19CxBa-83-Cv. Viruses 2023, 15, 239. https://doi.org/10.3390/v15010239
Tajima S, Maeki T, Nakayama E, Faizah AN, Kobayashi D, Isawa H, Maekawa Y, Bendryman SS, Mulyatno KC, Rohmah EA, et al. Growth, Pathogenesis, and Serological Characteristics of the Japanese Encephalitis Virus Genotype IV Recent Strain 19CxBa-83-Cv. Viruses. 2023; 15(1):239. https://doi.org/10.3390/v15010239
Chicago/Turabian StyleTajima, Shigeru, Takahiro Maeki, Eri Nakayama, Astri Nur Faizah, Daisuke Kobayashi, Haruhiko Isawa, Yoshihide Maekawa, Sri Subekti Bendryman, Kris Cahyo Mulyatno, Etik Ainun Rohmah, and et al. 2023. "Growth, Pathogenesis, and Serological Characteristics of the Japanese Encephalitis Virus Genotype IV Recent Strain 19CxBa-83-Cv" Viruses 15, no. 1: 239. https://doi.org/10.3390/v15010239
APA StyleTajima, S., Maeki, T., Nakayama, E., Faizah, A. N., Kobayashi, D., Isawa, H., Maekawa, Y., Bendryman, S. S., Mulyatno, K. C., Rohmah, E. A., Mori, Y., Sawabe, K., Ebihara, H., & Lim, C.-K. (2023). Growth, Pathogenesis, and Serological Characteristics of the Japanese Encephalitis Virus Genotype IV Recent Strain 19CxBa-83-Cv. Viruses, 15(1), 239. https://doi.org/10.3390/v15010239