
Manifestation of SARS-CoV-2 Infections in Mink Related to Host-, Virus- and Farm-Associated Factors, The Netherlands 2020

 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Sample and Data Collection on All 69 Infected Mink Farms
2.3. Additional Extensive Sample and Data Collection on Seven of the Infected Mink Farms
2.4. Data Analysis
2.4.1. Clinical and Diagnostic Data Analyses for All 69 Infected Mink Farms
2.4.2. Clinical and Diagnostic Data Analyses for the Seven Extensively Sampled Mink Farms
3. Results
3.1. Farm Characteristics
3.1.1. Type, Size, and Location of Farms
3.1.2. Humans on the Farm and Hygiene Protocols
3.1.3. Veterinary and Feed Service Contacts and Manure Management
3.1.4. Other Animals
3.2. Clinical and Diagnostic Representation of SARS-CoV-2 in Mink
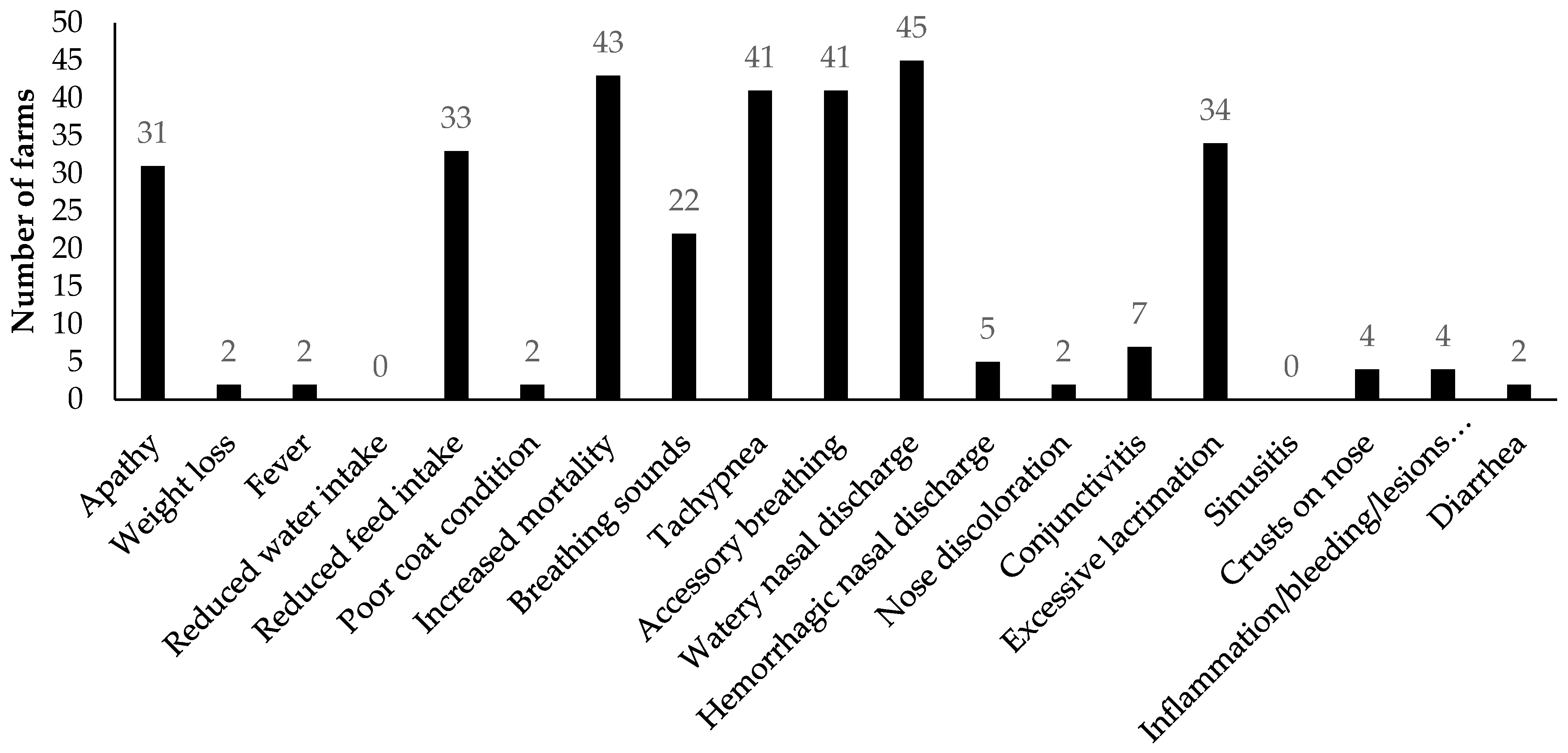
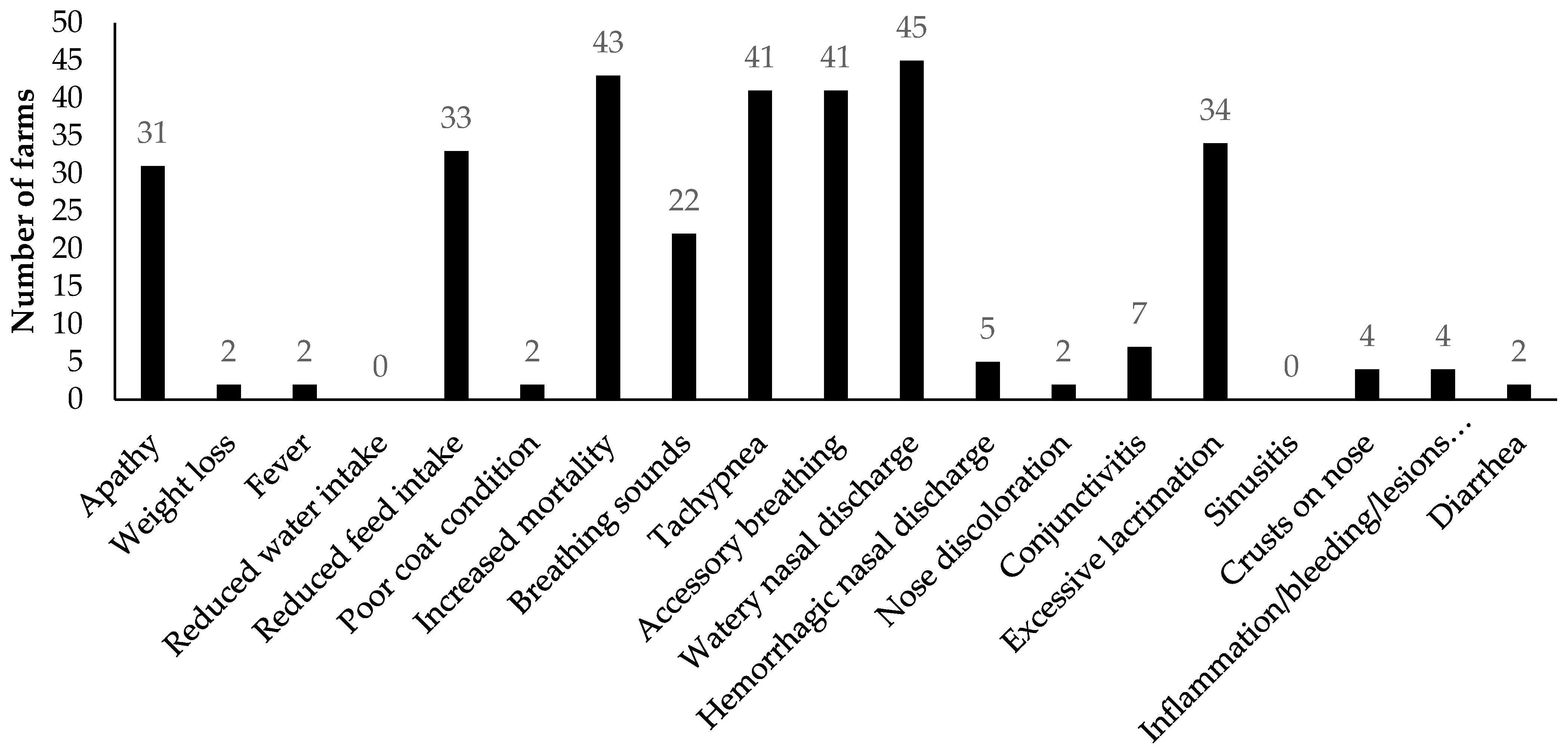
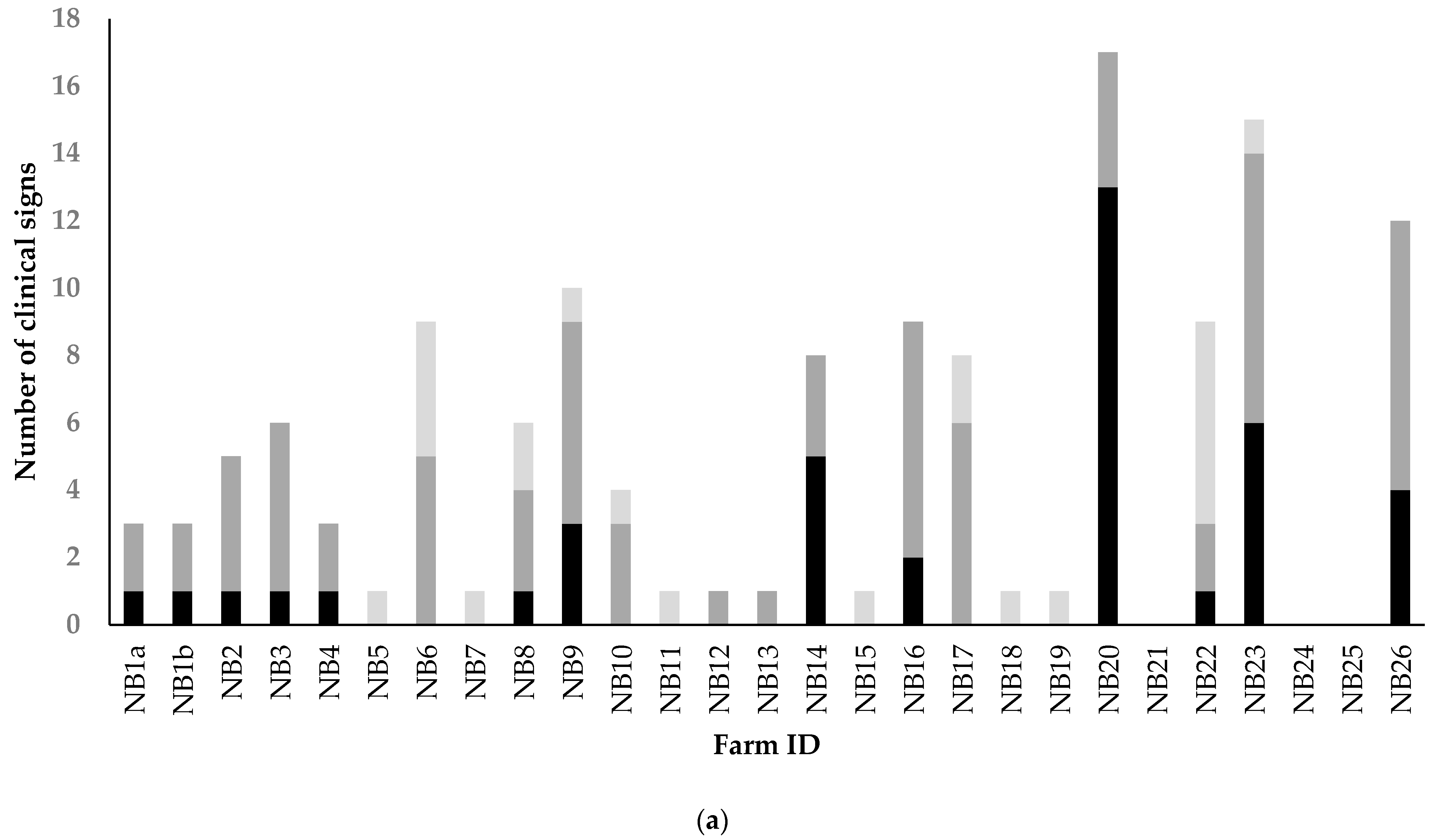
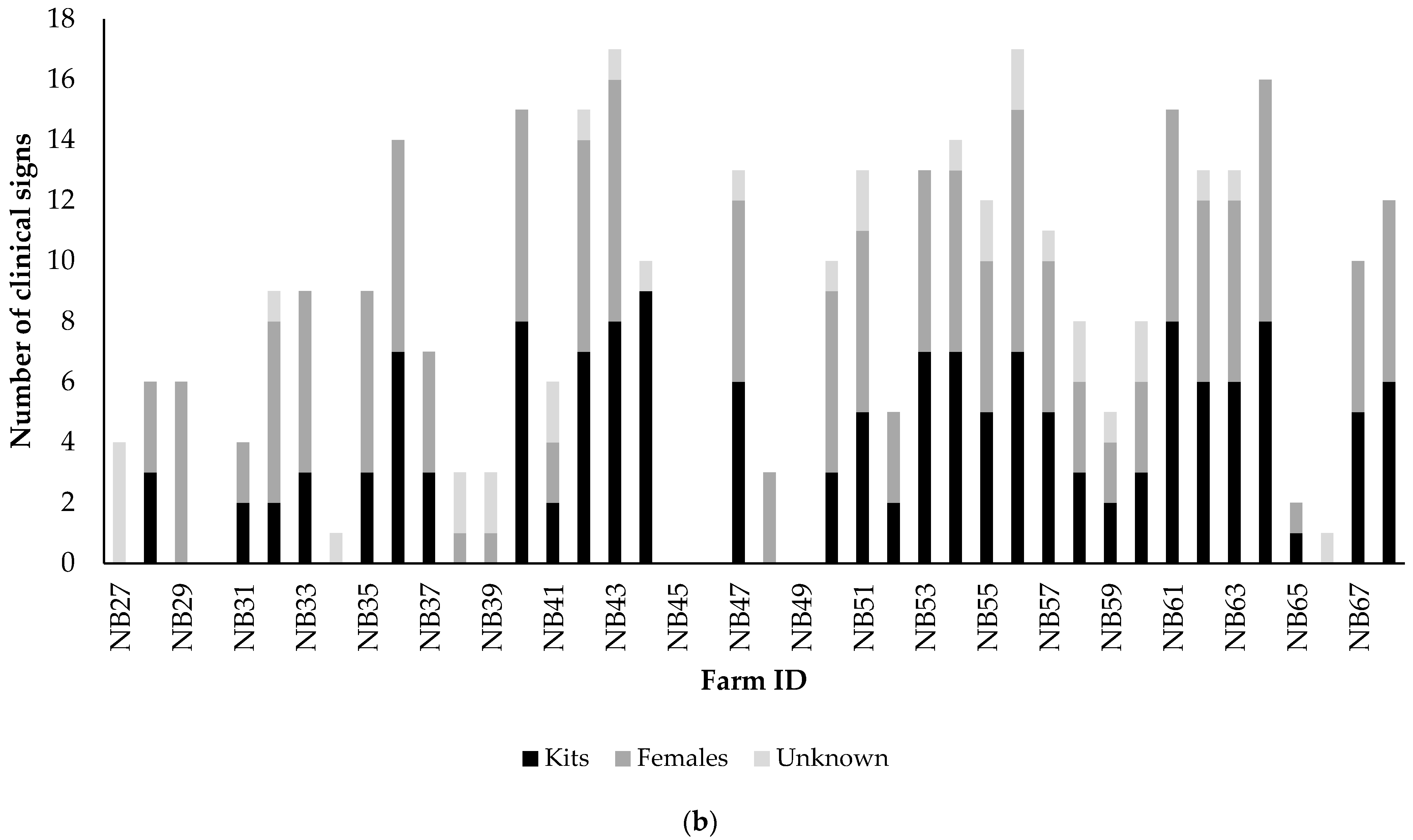
3.2.1. Analyses of Clinical Signs Observed on All Farms in Kits and Adult Mink
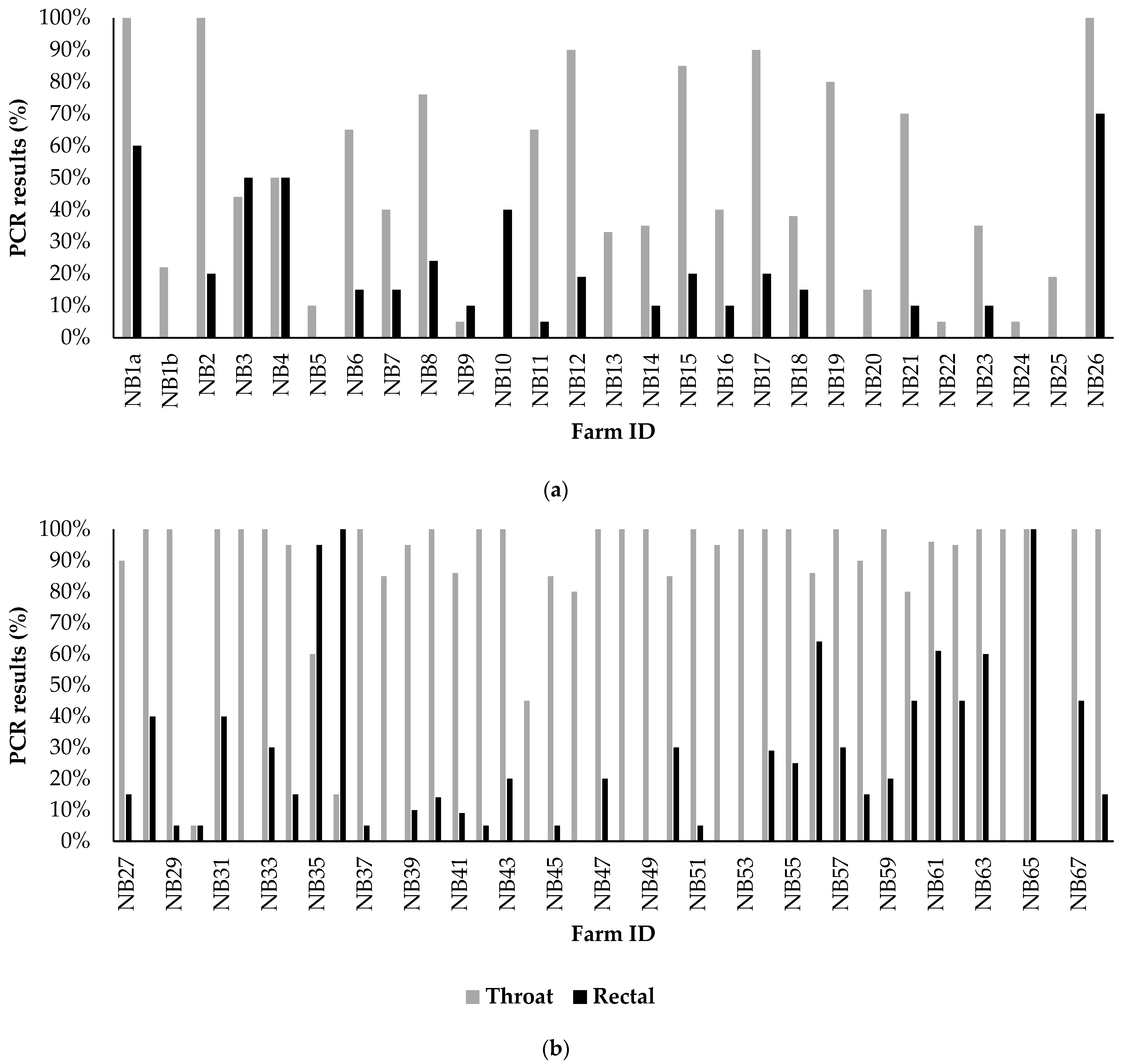
3.2.2. Analyses of Diagnostic Results on All Farms
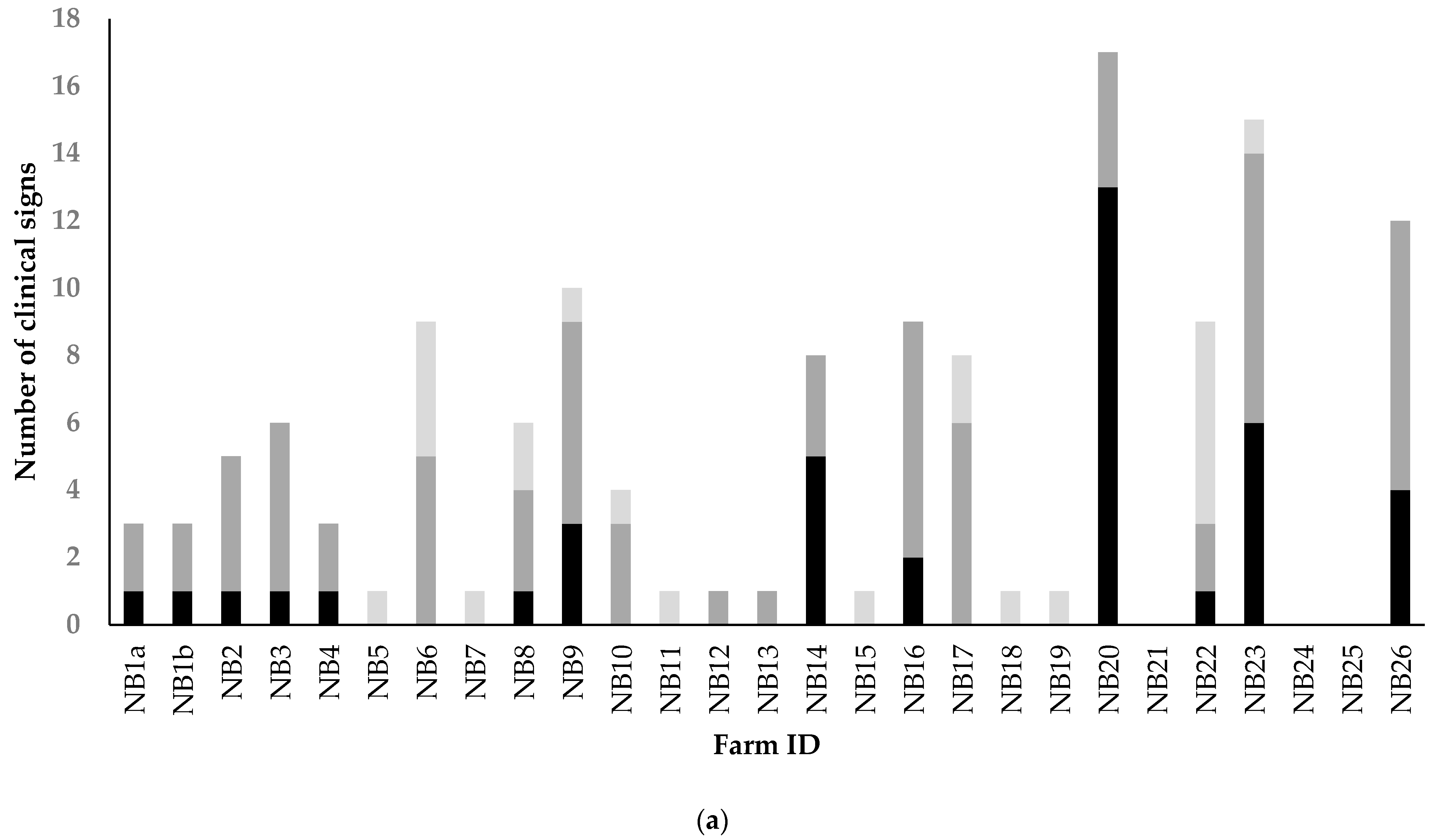
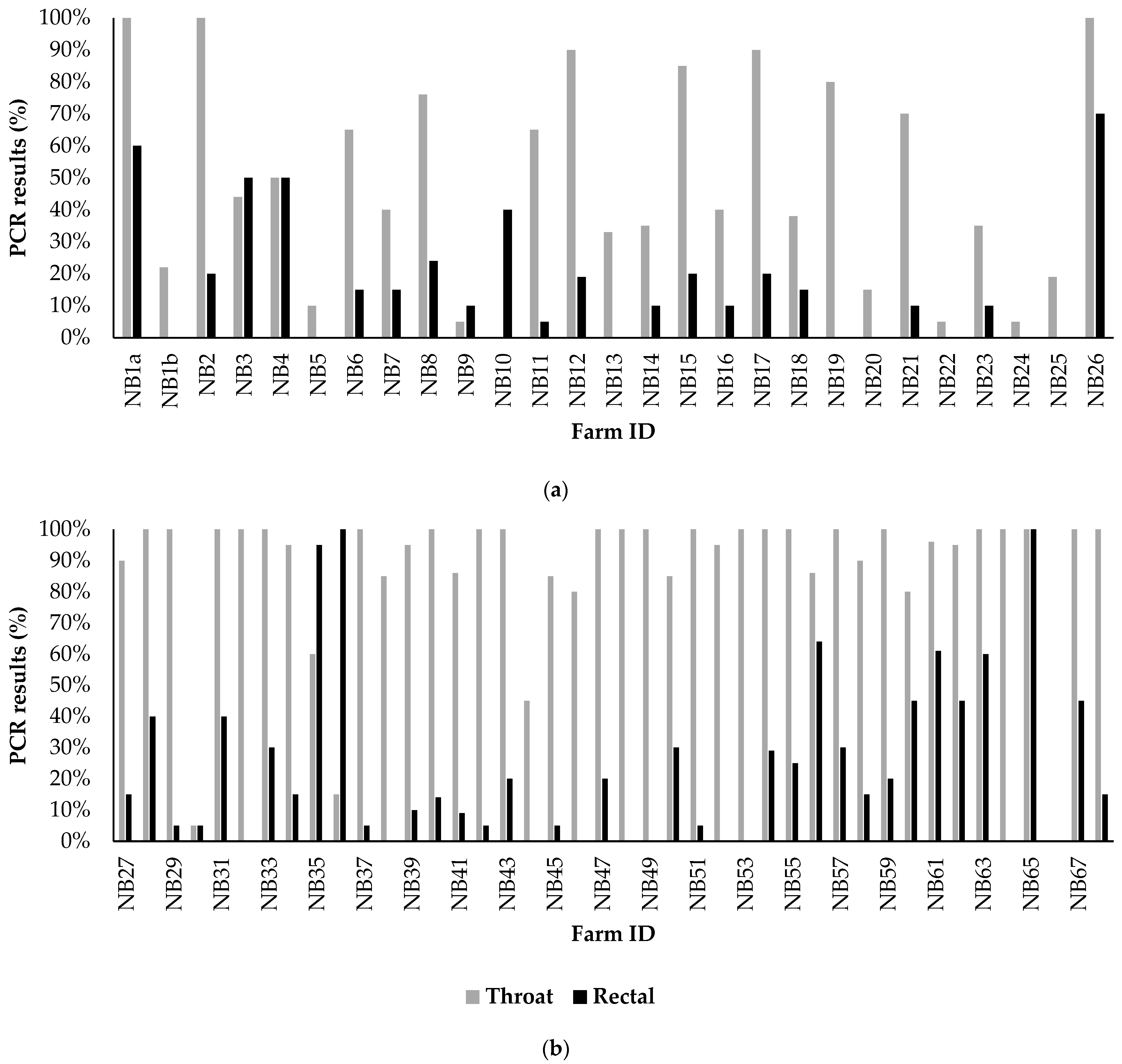
3.3. Results of Analyses of Extensive Random Sampling on Seven Mink Farms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bialek, S.; Boundy, E.; Bowen, V.; Chow, N.; Cohn, A.; Dowling, N.; Ellington, S.; Gierke, R.; Hall, A.; MacNeil, J.; et al. Severe Outcomes Among Patients with Coronavirus Disease 2019 (COVID-19)—United States, February 12–March 16, 2020. MMWR. Morb. Mortal. Wkly. Rep. 2020, 69, 343–346. [Google Scholar] [CrossRef]
- Emergency Situational Updates Weekly Operational Update on COVID-19—23 October 2020. 2020. Available online: https://www.who.int/publications/m/item/weekly-update-on-COVID-19—23-october (accessed on 26 July 2022).
- World Health Organisation Coronavirus Disease (COVID-19) Pandemic; Numbers at a Glance. 2021. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019?gclid=EAIaIQobChMIpZaJ4vz_7QIVUOJ3Ch3r9ws5EAAYASAAEgI_r_D_BwE (accessed on 26 July 2022).
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.; Wang, W.; Song, Z.; Hu, Y.; Tao, Z.; Tian, J.; Pei, Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention, and Control; Boklund, A.; Gortázar, C.; Pasquali, P.; Helen, R.; Stahl, K.; Stegeman, A.; Baldinelli, F.; Broglia, A.; Van Der Stede, Y.; et al. Monitoring of SARS-CoV-2 infection in mustelids. EFSA J. 2021, 19, e06459. [Google Scholar] [CrossRef]
- Koeppel, K.N.; Mendes, A.; Strydom, A.; Rotherham, L.; Mulumba, M.; Venter, M. SARS-CoV-2 Reverse Zoonoses to Pumas and Lions, South Africa. Viruses 2022, 14, 120. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. SARS-CoV-2 in Animals-Situation Report 12, 30 April 2022. Available online: https://www.woah.org/app/uploads/2022/05/sars-cov-2-situation-report-12.pdf (accessed on 26 July 2022).
- Melo, F.L.; Bezerra, B.; Luna, F.O.; Barragan, N.A.N.; Arcoverde, R.M.L.; Umeed, R.; Lucchini, K.; Attademo, F.L.N. Coronavirus (SARS-CoV-2) in Antillean Manatees (Trichechus Manatusmanatus). Res. Sq. 2022. [Google Scholar] [CrossRef]
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Oude Munnink, B.B.; Hakze-van der Honing, R.W.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.S.; et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Euro Surveill. 2020, 25, 2001005. [Google Scholar] [CrossRef] [PubMed]
- Pomorska-Mól, M.; Włodarek, J.; Gogulski, M.; Rybska, M. Review: SARS-CoV-2 infection in farmed minks—An overview of current knowledge on occurrence, disease and epidemiology. Animal 2021, 15, 100272. [Google Scholar] [CrossRef]
- van Aart, A.E.; Velkers, F.C.; Fischer, E.A.J.; Broens, E.M.; Egberink, H.; Zhao, S.; Engelsma, M.; Hakze-van der Honing, R.W.; Harders, F.; de Rooij, M.M.T.; et al. SARS-CoV-2 infection in cats and dogs in infected mink farms. Transbound. Emerg. Dis. 2021, 1–7. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-Coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Sikkema, R.S.; Velkers, F.C.; Nieuwenhuijse, D.F.; Fischer, E.A.J.; Meijer, P.A.; Bouwmeester-Vincken, N.; Rietveld, A.; Wegdam-Blans, M.C.A.; Tolsma, P.; et al. Adaptation, spread and transmission of SARS-CoV-2 in farmed minks and associated humans in the Netherlands. Nat. Commun. 2021, 12, 6802. [Google Scholar] [CrossRef] [PubMed]
- Badiola, J.J.; Otero, A.; Sevilla, E.; Marín, B.; García Martínez, M.; Betancor, M.; Sola, D.; Pérez Lázaro, S.; Lozada, J.; Velez, C.; et al. SARS-CoV-2 Outbreak on a Spanish Mink Farm: Epidemiological, Molecular, and Pathological Studies. Front. Vet. Sci. 2021, 8, 805004. [Google Scholar] [CrossRef] [PubMed]
- Boklund, A.; Hammer, A.S.; Quaade, M.L.; Rasmussen, T.B.; Lohse, L.; Strandbygaard, B.; Jørgensen, C.S.; Olesen, A.S.; Hjerpe, F.B.; Petersen, H.H.; et al. SARS-CoV-2 in Danish Mink Farms: Course of the Epidemic and a Descriptive Analysis of the Outbreaks in 2020. Animals 2021, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- More than 1 Million Mink Will Be Killed to Help Contain a Series of COVID-19 Outbreaks on Danish Farms. Available online: https://www.cnn.com/2020/10/13/world/denmark-mink-farms-covid-trnd/index.html (accessed on 26 July 2022).
- Chaintoutis, S.C.; Thomou, Z.; Mouchtaropoulou, E.; Tsiolas, G.; Chassalevris, T.; Stylianaki, I.; Lagou, M.; Michailidou, S.; Moutou, E.; Koenen, J.J.H.; et al. Outbreaks of SARS-CoV-2 in naturally infected mink farms: Impact, transmission dynamics, genetic patterns, and environmental contamination. PLoS Pathog. 2021, 17, 1009883. [Google Scholar] [CrossRef] [PubMed]
- Domańska-Blicharz, K.; Orłowska, A.; Smreczak, M.; Niemczuk, K.; Iwan, E.; Bomba, A.; Lisowska, A.; Opolska, J.; Trębas, P.; Potyrało, P.; et al. Mink SARS-CoV-2 Infection in Poland—Short Communication. J. Vet. Res. 2021, 65, 1–5. [Google Scholar] [CrossRef]
- World Health Organization; World Organisation for Animal Health; Food and Agriculture Organization of the United Nations. SARS-CoV-2 in Animals Used for Fur Farming. GLEWS+. 2021. Available online: https://www.woah.org/app/uploads/2021/03/glews-risk-assessment-fur-animals-sars-cov-2.pdf (accessed on 26 July 2022).
- Koopmans, M. SARS-CoV-2 and the Human-Animal Interface: Outbreaks on Mink Farms. 2020. Available online: https://www.thelancet.com/journals/laninf/article/PIIS1473-3099(20)30912-9/fulltext (accessed on 26 July 2022).
- United States Department of Agriculture Lithuania: COVID-19 Outbreak on Lithuanian Mink Farm. 2020. Available online: https://www.fas.usda.gov/data/lithuania-COVID-19-outbreak-lithuanian-mink-farm (accessed on 26 July 2022).
- Detection of New SARS-CoV-2 Variants Related to Mink—12 November 2020. Available online: https://www.ecdc.europa.eu/en/publications-data/detection-new-sars-cov-2-variants-mink (accessed on 26 July 2022).
- Eklund, C.M.; Hadlow, W.J.; Kennedy, R.C.; Boyle, C.C.; Jackson, T.A. Aleutian disease of mink: Properties of the etiologic agent and the host responses. J. Infect. Dis. 1968, 118, 510–526. [Google Scholar] [CrossRef] [PubMed]
- Fertner, M.; Pedersen, K.; Jensen, V.F.; Larsen, G.; Lindegaard, M.; Hansen, J.E.; Chriél, M. Within-farm prevalence and environmental distribution of livestock-associated methicillin-resistant Staphylococcus aureus in farmed mink (Neovison vison). Vet. Microbiol. 2019, 231, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Kashtanov, S.; Salnikova, L. Aleutian Mink Disease: Epidemiological and Genetic Aspects. Biol. Bull. Rev. 2018, 8, 104–113. [Google Scholar] [CrossRef]
- Gorham, J.R.; Evermann, J.F.; Ward, A.; Pearson, R.; Shen, D.; Hartsough, G.R.; Leathers, C. “Detection of coronavirus-like particles from mink with epizootic catarrhal gastroenteritis. Can. J. Vet. Res. 1990, 54, 383–384. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1255674/ (accessed on 26 July 2022). [PubMed]
- Anistoroaei, R.; Krogh, A.K.; Christensen, K. A frameshift mutation in the LYST gene is responsible for the Aleutian color and the associated Chédiak-Higashi syndrome in American mink. Anim. Genet. 2013, 44, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Blume, R.S.; Padgett, G.A.; Wolff, S.M.; Bennett, J.M. Giant neutrophil granules in the Chediak-Higashi syndrome of man, mink, cattle and mice. Can. J. Comp. Med. 1969, 33, 271–274. [Google Scholar] [PubMed]
- Jespersen, A.; Jensen, H.E.; Agger, J.F.; Heegaard, P.M.H.; Damborg, P.; Aalbæk, B.; Hammer, A.S. The effect of color type on early wound healing in farmed mink (Neovison vison). BMC Vet. Res. 2017, 13, 135. [Google Scholar] [CrossRef] [PubMed]
- Oude Munnink, B.B.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; van der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M.; et al. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef]
- Díaz, A.V.; Walker, M.; Webster, J.P. Surveillance and control of SARS-CoV-2 in mustelids: An evolutionary perspective. Evol. Appl. 2021, 14, 2715–2725. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. 2020, 25, 2000045. [Google Scholar] [CrossRef]
- Van der Heijden, H. Serological Screening of Dutch Mink for SARS-CoV-2 by ELISA; Royal GD: Deventer, The Netherlands, 2022; in preparation. [Google Scholar]
- Oude Munnink, B.B.; Sikkema, R.S.; Nieuwhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; van der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M.; et al. Jumping Back and Forth: Anthropozoonotic and Zoonotic Transmission of SARS-CoV-2 on Mink Farms. BioRxiv 2020. [Google Scholar] [CrossRef]
- Velkers, F.; Molenaar, R.J.; La Haye, M.; Spierenburg, M.; Smit, L.; Sikkema, R. SARS-CoV-2 on mink farms in the Netherlands; a One Health investigation. Ned. Tijdschr. Med. Microbiol. 2021, 2021, 105–111. Available online: https://www.nvmm.nl/media/4277/105-111-velkers.pdf (accessed on 26 July 2022).
- RStudio Team. Integrated Development Environment for R; RStudio PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 26 July 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Software 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Velkers, F.C.; One Health Mink Outbreak Investigation Consortium. Onderzoek NAAR Besmettingsroutes van SARS-CoV-2 op nertsenbedrijven. Research Report for Dutch Ministries of Health, Welfare and Sport, and Agriculture, Nature and Food Quality. 2021. Available online: https://open.overheid.nl/repository/ronl-b5de3a58-82d7-4717-b358-b92a9c888c3c/1/pdf/onderzoek-naar-besmettingsroutes-van-sarscov-2-op-nertsenbedrijven.pdf (accessed on 26 July 2022). (In Dutch).
- Hissel, B.; Slaterus, R.; Janssen, R.; van Dijk, J. Vogels op en rond Nertsenbedrijven; Sovon-Report 2021/20; Sovon Vogelonderzoek: Nijmegen, The Netherlands, 2021; pp. 1–26. [Google Scholar]
- Molenaar, R.J.; Vreman, S.; Hakze-van der Honing, R.W.; Zwart, R.; de Rond, J.; Weesendorp, E.; Smit, L.A.M.; Koopmans, M.; Bouwstra, R.; Stegeman, A.; et al. Clinical and Pathological Findings in SARS-CoV-2 Disease Outbreaks in Farmed Mink (Neovison vison). Vet. Pathol. 2020, 57, 653–657. [Google Scholar] [CrossRef]
- Clayton, E.; Ackerley, J.; Aelmans, M.; Ali, N.; Ashcroft, Z.; Ashton, C.; Barker, R.; Budryte, V.; Burrows, C.; Cai, S.; et al. Structural Bases of Zoonotic and Zooanthroponotic Transmission of SARS-CoV-2. Viruses 2022, 14, 418. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.C.S.; Lam, S.D.; Richard, D.; Owen, C.J.; Berchtold, D.; Orengo, C.; Nair, M.S.; Kuchipudi, S.V.; Kapur, V.; van Dorp, L.; et al. Transmission of SARS-CoV-2 from humans to animals and potential host adaptation. Nat. Commun. 2022, 13, 2988. [Google Scholar] [CrossRef]
- Devaux, C.A.; Pinault, L.; Delerce, J.; Raoult, D.; Levasseur, A.; Frutos, R. Spread of Mink SARS-CoV-2 Variants in Humans: A Model of Sarbecovirus Interspecies Evolution. Front. Microbiol. 2021, 12, 675528. [Google Scholar] [CrossRef] [PubMed]
- Devaux, C.A.; Pinault, L.; Osman, I.O.; Raoult, D. Can ACE2 Receptor Polymorphism Predict Species Susceptibility to SARS-CoV-2? Front. Public Health 2021, 8, 608765. [Google Scholar] [CrossRef]
- Fantini, J.; Yahi, N.; Azzaz, F.; Chahinian, H. Structural dynamics of SARS-CoV-2 variants: A health monitoring strategy for anticipating Covid-19 outbreaks. J. Infection 2021, 83, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, G.; Wang, Y.; Ren, W.; Zhao, X.; Ji, F.; Zhu, Y.; Feng, F.; Gong, M.; Ju, X.; et al. Functional and genetic analysis of viral receptor ACE2 orthologs reveals a broad potential host range of SARS-CoV-2. Proc. Natl. Acad. Sci. USA. 2021, 118, e2025373118. [Google Scholar] [CrossRef]
- Frutos, R.; Pliez, O.; Gavotte, L.; Devaux, C.A. There is no “origin” to SARS-CoV-2. Environ. Res. 2022, 207, 112173. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Lean, F.Z.X.; Núñez, A.; Spiro, S.; Priestnall, S.L.; Vreman, S.; Bailey, D.; James, J.; Wrigglesworth, E.; Suarez-Bonnet, A.; Conceicao, C.; et al. Differential susceptibility of SARS-CoV-2 in animals: Evidence of ACE2 host receptor distribution in companion animals, livestock and wildlife by immunohistochemical characterisation. Transbound. Emerg. Dis. 2022, 69, 2275–2286. [Google Scholar] [CrossRef]
- Deering, K. Information from the WVDL Regarding Mustelid Species Affected by SARS-CoV-2. 2020. Available online: https://www.wvdl.wisc.edu/index.php/mink-and-suspected-sars-cov-2-infection/ (accessed on 26 July 2022).
- Shuai, L.; Zhong, G.; Yuan, Q.; Wen, Z.; Wang, C.; He, X.; Liu, R.; Wang, J.; Zhao, Q.; Liu, Y.; et al. Replication, pathogenicity, and transmission of SARS-CoV-2 in minks. Natl. Sci. Rev. 2021, 8, nwaa291. [Google Scholar] [CrossRef]
- Prince, T.; Smith, S.L.; Radford, A.D.; Solomon, T.; Hughes, G.L.; Patterson, E.I. SARS-CoV-2 Infections in Animals: Reservoirs for Reverse Zoonosis and Models for Study. Viruses 2021, 13, 494. [Google Scholar] [CrossRef]
- Bwire, G.M. Coronavirus: Why Men are More Vulnerable to Covid-19 Than Women? SN Compr. Clin. Med. 2020, 2, 874–876. [Google Scholar] [CrossRef]
- Pradhan, A.; Olsson, P. Sex differences in severity and mortality from COVID-19: Are males more vulnerable? Biol. Sex. Differ. 2020, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Gorham, J.R.; Leader, R.W.; Padgett, G.A.; Burger, D.; Henson, J.B. Some Observations of the Natural Occurrence of Aleutian Disease. Slow Latent Temp. Virus Infect. NINDB Monogr. 1965, 2, 279–285. [Google Scholar]
- Jensen, T.H.; Hammer, A.S.; Chriél, M. Monitoring chronic infection with a field strain of Aleutian mink disease virus. Vet. Microbiol. 2014, 168, 420–427. [Google Scholar] [CrossRef]
- Osterrieder, N.; Bertzbach, L.D.; Dietert, K.; Abdelgawad, A.; Vladimirova, D.; Kunec, D.; Hoffmann, D.; Beer, M.; Gruber, A.D.; Trimpert, J. Age-Dependent Progression of SARS-CoV-2 Infection in Syrian Hamsters. Viruses 2020, 12, 779. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Liu, F.; Ye, Z.; Song, W.; Lee, A.C.Y.; Shuai, H.; Lu, L.; To, K.K.; Chan, J.F.; et al. Age-associated SARS-CoV-2 breakthrough infection and changes in immune response in a mouse model. Emerg. Microbes Infect. 2022, 11, 368–383. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Baum, A.; Fulton, B.O.; Wloga, E.; Copin, R.; Pascal, K.E.; Russo, V.; Giordano, S.; Lanza, K.; Negron, N.; Ni, M.; et al. Antibody cocktail to SARS-CoV-2 spike protein prevents rapid mutational escape seen with individual antibodies. Science 2020, 369, 1014–1018. [Google Scholar] [CrossRef]
- Gryseels, S.; De Bruyn, L.; Gyselings, R.; Calvignac-Spencer, S.; Leendertz, F.H.; Leirs, H. Risk of human-to-wildlife transmission of SARS-CoV-2. Mamm. Rev. 2020, 51, 272–292. [Google Scholar] [CrossRef] [PubMed]
- de Rooij, M.M.T.; Hakze-Van der Honing, R.W.; Hulst, M.M.; Harders, F.; Engelsma, M.; van de Hoef, W.; Me-liefste, K.; Nieuwenweg, S.; Oude Munnink, B.B.; van Schothorst, I.; et al. Occupational and environmental exposure to SARS-CoV-2 in and around infected mink farms. Occup. Environ. Med. 2021, 78, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Boender, G.J.; Hagenaars, T.J.; Elbers, A.R.; Gethmann, J.M.; Meroc, E.; Guis, H.; de Koeijer, A.A. Confirmation of spatial patterns and temperature effects in Bluetongue virus serotype-8 transmission in NW-Europe from the 2007 reported case data. Vet. Res. 2014, 45, 75. [Google Scholar] [CrossRef]
- de Koeijer, A.A.; Boender, G.J.; Nodelijk, G.; Staubach, C.; Meroc, E.; Elbers, A.R.W. Quantitative analysis of transmission parameters for bluetongue virus serotype 8 in Western Europe in 2006. Vet. Res. 2011, 42, 53. [Google Scholar] [CrossRef]
- Boender, G.J.; Hagenaars, T.J.; Bouma, A.; Nodelijk, G.; Elbers, A.R.W.; de Jong, M.C.M.; van Boven, M. Risk Maps for the Spread of Highly Pathogenic Avian Influenza in Poultry. PLoS Comput. Biol. 2007, 3, e71. [Google Scholar] [CrossRef]
- Boender, G.J.; van den Hengel, R.; van Roermund, H.J.W.; Hagenaars, T.J. The influence of between-farm distance and farm size on the spread of classical swine fever during the 1997–1998 epidemic in The Netherlands. PLoS ONE 2014, 9, e95278. [Google Scholar] [CrossRef]
- Boender, G.J.; van Roermund, H.J.W.; de Jong, M.C.M.; Hagenaars, T.J. Transmission risks and control of foot-and-mouth disease in The Netherlands: Spatial patterns. Epidemics 2010, 2, 36–47. [Google Scholar] [CrossRef]
- Xiang, B.; Yang, L.; Ye, Z.; Ren, T.; Ye, Y. Vaccination of susceptible animals against SARS-CoV-2. J. Infect. 2022, 84, e48–e49. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observations | Yes | No | Unknown | Total |
|---|---|---|---|---|
| Farm characteristics | ||||
| Farm distance (<3 km) 1 | 51 | 18 | 0 | 69 |
| Farm distance with the same virus cluster (<3 km) 2 | 43 | 25 | 3 | 69 |
| Type of housing | Sheds = 39; Halls = 21; Both = 6 | 3 | 69 | |
| Farm size 3 | Small = 27; Medium = 18; Big = 24 | 0 | 69 | |
| Sharing of materials and vehicles | 6 | 57 | 6 | 69 |
| Same owner | 32 | 37 | 0 | 69 |
| Shed/hall visitors (≤5) | 35 | 34 | 0 | 69 |
| Exchange of employees (≥1) | 36 | 29 | 4 | 69 |
| Permanent employees (≤2) | 44 | 25 | 0 | 69 |
| Seasonal workers (≤3) | 54 | 15 | 0 | 69 |
| Temporary employees (≥1) | 18 | 51 | 0 | 69 |
| Symptomatic employees 4 | 31 | 20 | 18 | 69 |
| Manure | 21 | 39 | 9 | 69 |
| Feed supplier 5 | A = 45; B = 15; C = 9 | 0 | 69 | |
| Veterinarian 6 | A = 54 | B = 15 | 0 | 69 |
| Cats | 9 | 51 | 9 | 69 |
| Dogs | 41 | 25 | 3 | 69 |
| Feral cats or neighbor cats | 50 | 17 | 2 | 69 |
| Other animals 7 | 27 | 22 | 20 | 69 |
| Wildlife including bats and birds | 53 | 1 | 15 | 69 |
| Rodents | 18 | 32 | 19 | 69 |
| Insects | 21 | 3 | 45 | 69 |
| Outbreak characteristics | ||||
| Date of diagnosis 8 | <August = 27 | ≥August = 42 | 0 | 69 |
| Virus type | ||||
| Virus cluster 9 | A = 41; B = 1; C = 15; D = 7; E = 1 | 4 | 69 | |
| Variable | Categories | N 1 | Mean 2 | OR 3 | 95% CI 3 |
|---|---|---|---|---|---|
| Cluster | A | 41 | 5.27 | Ref 3 | |
| C vs. A | 15 | 4.00 | 0.81 | 0.45–1.45 | |
| D vs. A | 7 | 1.57 | 0.22 | 0.08–0.53 * | |
| Cluster | C | 15 | 4.00 | Ref | |
| A vs. C | 41 | 5.27 | 1.23 | 0.69–2.21 | |
| D vs. C | 7 | 1.57 | 0.27 | 0.09–0.72 * | |
| Diagnosis kits 4 | <August | 27 | 1.44 | Ref | |
| ≥August | 42 | 3.57 | 4.25 | 1.90–10.39 * | |
| Diagnosis females 5 | <August | 27 | 2.67 | Ref | |
| ≥August | 42 | 3.95 | 1.80 | 0.99–3.35 |
| Independent Variable | Categories | N 1 | Clinical Signs (%) 2 | OR 3 | 95%CI 3 |
|---|---|---|---|---|---|
| Coat color 4 | Dark: wild, brown, MAH | 750 | 11.9 | Ref 3 | |
| Light: SBL and SCR | 466 | 4.07 | 0.35 | 0.29–0.60 * | |
| Other: Jag, pearl, black, other | 187 | 2.19 | 0.41 | 0.10–1.42 | |
| Sex/age 5 | Juvenile males | 388 | 11.08 | Ref 3 | |
| Juvenile females | 521 | 4.99 | 0.33 | 0.19–0.60 * | |
| Adult females | 494 | 8.70 | 1.10 | 0.50–1.29 | |
| Sex/age 5 | Juvenile females | 521 | 4.99 | Ref 3 | |
| Juvenile males | 388 | 11.08 | 3.02 | 1.78–5.25 * | |
| Adult females | 494 | 8.70 | 2.64 | 1.44–4.19 * | |
| ELISA | Negative | 1348 | 7.03 | Ref 3 | |
| Positive | 55 | 23.64 | 2.64 | 1.24–5.38 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolters, W.J.; de Rooij, M.M.T.; Molenaar, R.J.; de Rond, J.; Vernooij, J.C.M.; Meijer, P.A.; Oude Munnink, B.B.; Sikkema, R.S.; van der Spek, A.N.; Spierenburg, M.A.H.; et al. Manifestation of SARS-CoV-2 Infections in Mink Related to Host-, Virus- and Farm-Associated Factors, The Netherlands 2020. Viruses 2022, 14, 1754. https://doi.org/10.3390/v14081754
Wolters WJ, de Rooij MMT, Molenaar RJ, de Rond J, Vernooij JCM, Meijer PA, Oude Munnink BB, Sikkema RS, van der Spek AN, Spierenburg MAH, et al. Manifestation of SARS-CoV-2 Infections in Mink Related to Host-, Virus- and Farm-Associated Factors, The Netherlands 2020. Viruses. 2022; 14(8):1754. https://doi.org/10.3390/v14081754
Chicago/Turabian StyleWolters, Wendy J., Myrna M. T. de Rooij, Robert Jan Molenaar, Jan de Rond, J. C. M. Vernooij, Paola A. Meijer, Bas B. Oude Munnink, Reina S. Sikkema, Arco N. van der Spek, Marcel A. H. Spierenburg, and et al. 2022. "Manifestation of SARS-CoV-2 Infections in Mink Related to Host-, Virus- and Farm-Associated Factors, The Netherlands 2020" Viruses 14, no. 8: 1754. https://doi.org/10.3390/v14081754
APA StyleWolters, W. J., de Rooij, M. M. T., Molenaar, R. J., de Rond, J., Vernooij, J. C. M., Meijer, P. A., Oude Munnink, B. B., Sikkema, R. S., van der Spek, A. N., Spierenburg, M. A. H., Hakze-van der Honing, R. W., van der Poel, W. H. M., Koopmans, M. P. G., Stegeman, J. A., Smit, L. A. M., Augustijn-Schretlen, M., & Velkers, F. C. (2022). Manifestation of SARS-CoV-2 Infections in Mink Related to Host-, Virus- and Farm-Associated Factors, The Netherlands 2020. Viruses, 14(8), 1754. https://doi.org/10.3390/v14081754