
Pathophysiology of Influenza D Virus Infection in Specific-Pathogen-Free Lambs with or without Prior Mycoplasma ovipneumoniae Exposure

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry
2.2. Health Monitoring
2.3. M. ovipneumoniae Preparation
2.4. Virus Preparation
2.5. Intranasal Inoculations
2.6. Sampling
2.6.1. Nasal and Rectal Swabs
2.6.2. Serum Collection
2.6.3. Fecal Samples
2.7. Determining Pathogen Burden
2.7.1. M. ovipneumoniae Quantification
2.7.2. IDV Quantification
2.8. Hemagglutinin Inhibition (HI) Assay
2.9. Acute Phase Protein and L-Lactate Quantification
2.10. Statistical Analyses
3. Results
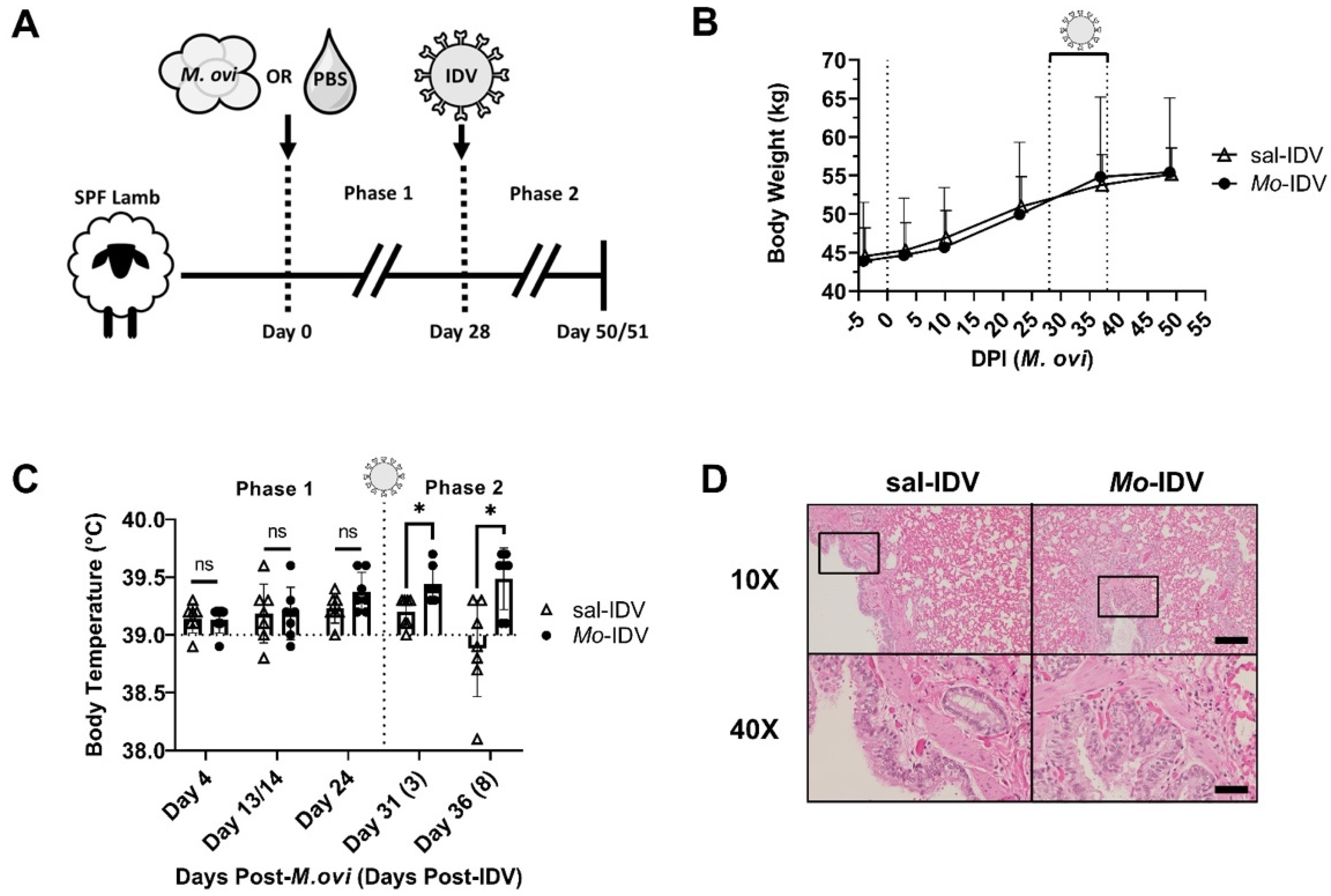
3.1. Prior M. ovipneumoniae Infection Elicits a Subclinical Response in IDV-Infected Lambs
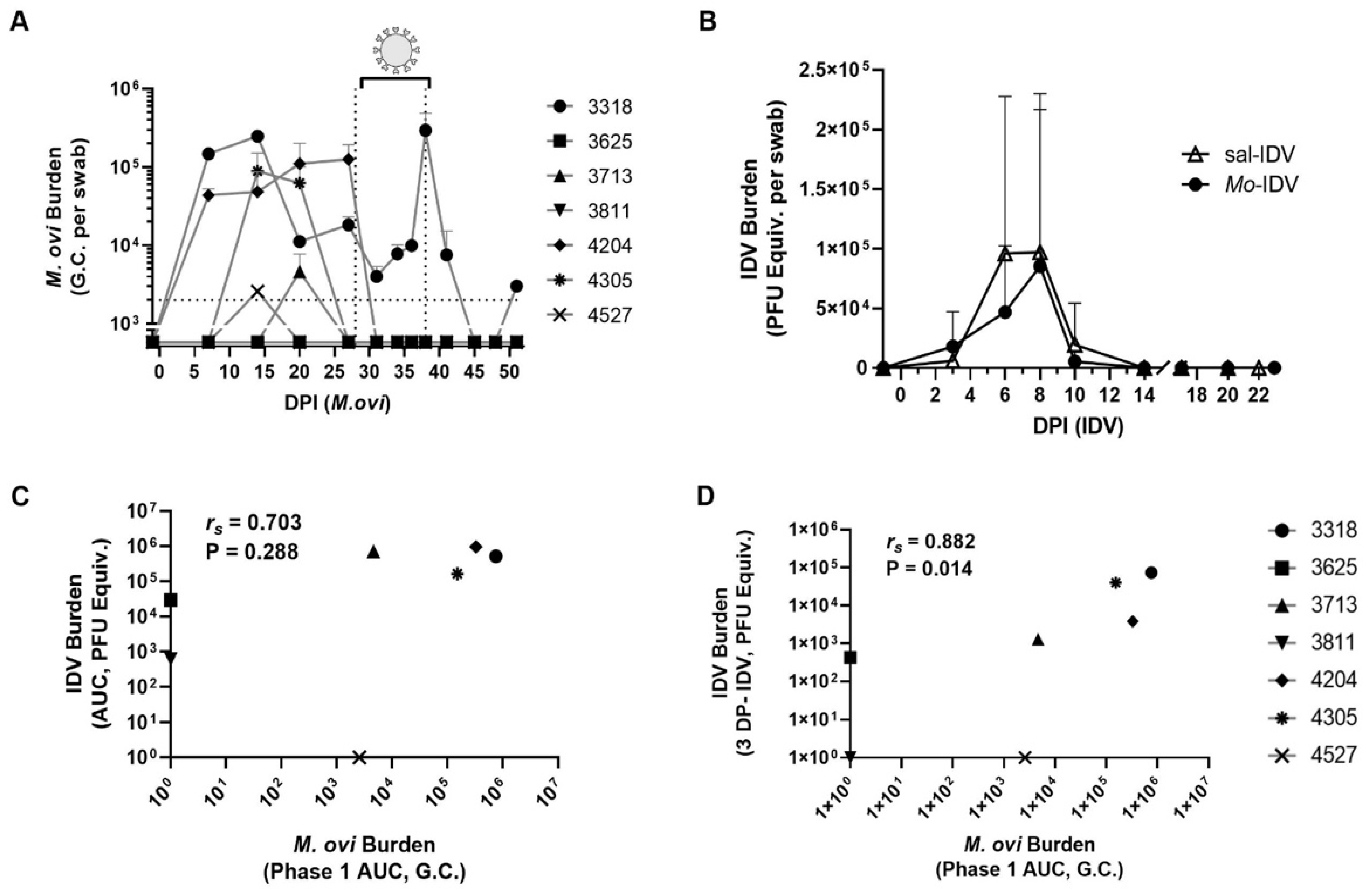
3.2. Recent M. ovipneumoniae Infection Has a Minimal Impact on IDV Shedding
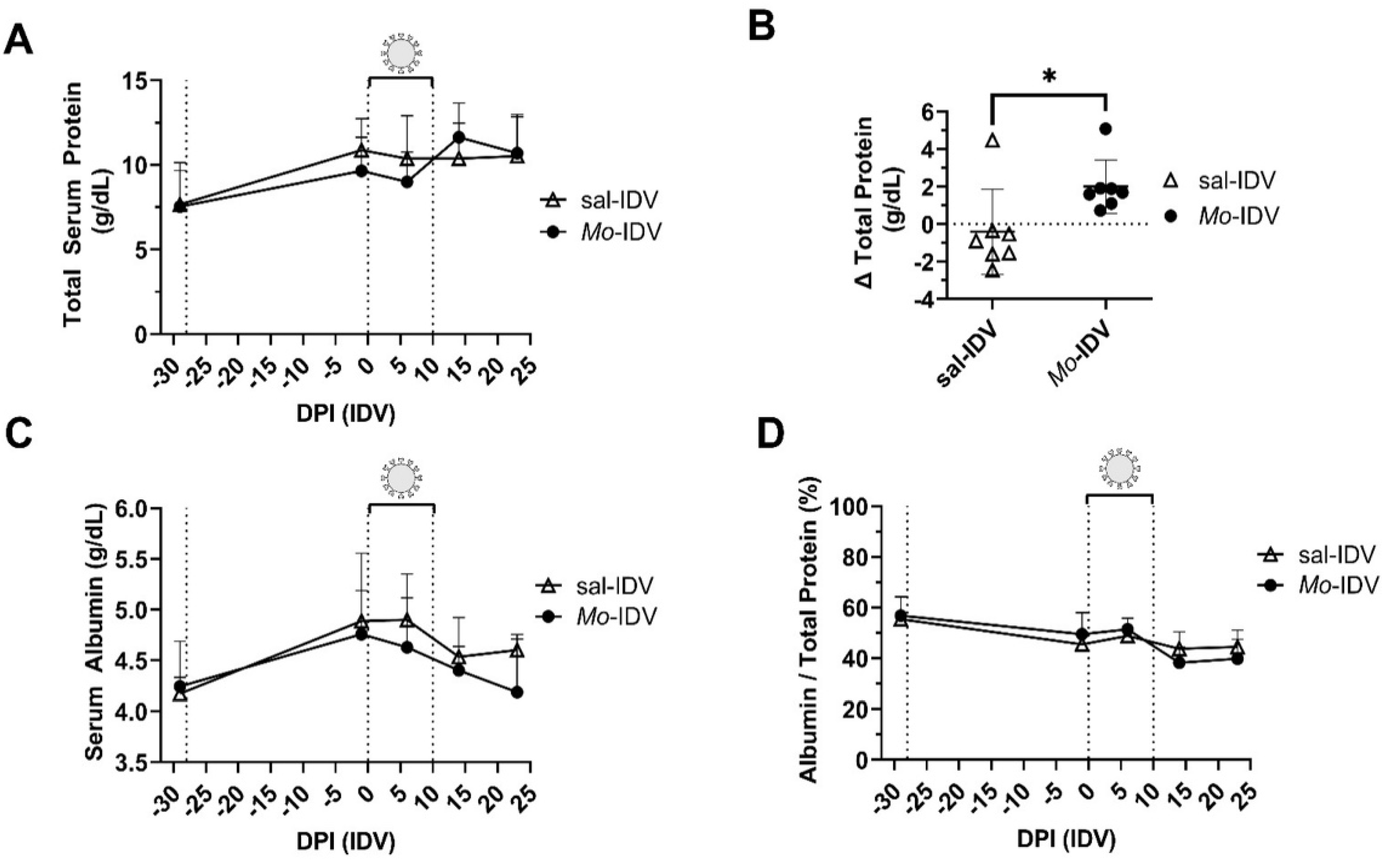
3.3. Total Serum Protein and Acute Phase Response
3.4. IDV Infection Elicits a Neutralizing Antibody Response in SPF Lambs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verma, A.K.; Bansal, S.; Bauer, C.; Muralidharan, A.; Sun, K. Influenza Infection Induces Alveolar Macrophage Dysfunction and Thereby Enables Noninvasive Streptococcus Pneumoniae to Cause Deadly Pneumonia. J. Immunol. 2020, 205, 1601–1607. [Google Scholar] [CrossRef] [PubMed]
- Sender, V.; Hentrich, K.; Henriques-Normark, B. Virus-Induced Changes of the Respiratory Tract Environment Promote Secondary Infections with Streptococcus Pneumoniae. Front. Cell. Infect. Microbiol. 2021, 11, 643326. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zeng, Y.; Wei, Y.; Wang, Q.; Liu, M.; Zhang, B.; Liu, J.; Zhu, Q.; Xu, S. Influenza D Virus Matrix Protein 1 Restricts the Type I Interferon Response by Degrading TRAF6. Virology 2022, 568, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Li, Y.; Jiang, Z.; Deng, G.; Li, M.; Liu, X.; Wang, Y. A ROS-Dependent and Caspase-3-Mediated Apoptosis in Sheep Bronchial Epithelial Cells in Response to Mycoplasma Ovipneumoniae Infections. Vet. Immunol. Immunopathol. 2017, 187, 55–63. [Google Scholar] [CrossRef]
- Niang, M.; Rosenbusch, R.F.; Lopez-Virella, J.; Kaeberle, M.L. Expression of Functions by Normal Sheep Alveolar Macrophages and Their Alteration by Interaction with Mycoplasma Ovipneumoniae. Vet. Microbiol. 1997, 58, 31–43. [Google Scholar] [CrossRef]
- Khan, M.A.; Kaeberle, M.L. Mitogenic Activity of Mycoplasma Ovipneumoniae Antigens for Sensitized Ovine Lymphocytes. Pak. Vet. J. 1995, 15, 117–120. [Google Scholar]
- Shepardson, K.M.; Larson, K.; Johns, L.L.; Stanek, K.; Cho, H.; Wellham, J.; Henderson, H.; Rynda-Apple, A. IFNAR2 Is Required for Anti-Influenza Immunity and Alters Susceptibility to Post-Influenza Bacterial Superinfections. Front. Immunol. 2018, 9, 2589. [Google Scholar] [CrossRef]
- Shepardson, K.; Larson, K.; Cho, H.; Johns, L.L.; Malkoc, Z.; Stanek, K.; Wellhman, J.; Zaiser, S.; Daggs-Olson, J.; Moodie, T.; et al. A Novel Role for PDZ-Binding Motif of Influenza A Virus Nonstructural Protein 1 in Regulation of Host Susceptibility to Postinfluenza Bacterial Superinfections. Viral Immunol. 2019, 32, 131–143. [Google Scholar] [CrossRef]
- Shepardson, K.M.; Larson, K.; Morton, R.V.; Prigge, J.R.; Schmidt, E.E.; Huber, V.C.; Rynda-Apple, A. Differential Type I Interferon Signaling Is a Master Regulator of Susceptibility to Postinfluenza Bacterial Superinfection. mBio 2016, 7, e00506-16. [Google Scholar] [CrossRef]
- Skelton, R.M.; Shepardson, K.M.; Hatton, A.; Wilson, P.T.; Sreenivasan, C.; Yu, J.; Wang, D.; Huber, V.C.; Rynda-Apple, A. Contribution of Host Immune Responses Against Influenza D Virus Infection Toward Secondary Bacterial Infection in a Mouse Model. Viruses 2019, 11, 994. [Google Scholar] [CrossRef]
- Johnson, T.; Jones, K.; Jacobson, B.T.; Schearer, J.; Adams, N.; Thornton, I.; Mosdal, C.; Jones, S.; Jutila, M.; Rynda-Apple, A.; et al. Experimental Infection of Specific-Pathogen-Free Domestic Lambs with Mycoplasma Ovipneumoniae Causes Asymptomatic Colonization of the Upper Airways That Is Resistant to Antibiotic Treatment. Vet. Microbiol. 2022, 265, 109334. [Google Scholar] [CrossRef] [PubMed]
- USDA APHIS. Mycoplasma Ovipneumoniae. Available online: https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/sa_animal_disease_information/sheep-goat/movi/mycoplasma-ovipneumoniae (accessed on 26 February 2022).
- Alley, M.R.; Quinlan, J.R.; Clarke, J.K. The Prevalence of Mycoplasma Ovipneumoniae and Mycoplasma Arginini in the Respiratory Tract of Sheep. N. Z. Vet. J. 1975, 23, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Manlove, K.; Branan, M.; Baker, K.; Bradway, D.; Cassirer, E.F.; Marshall, K.L.; Miller, R.S.; Sweeney, S.; Cross, P.C.; Besser, T.E. Risk Factors and Productivity Losses Associated with Mycoplasma Ovipneumoniae Infection in United States Domestic Sheep Operations. Prev. Vet. Med. 2019, 168, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Alley, M.R.; Clarke, J.K. The Experimental Transmission of Ovine Chronic Non-Progressive Pneumonia. N. Z. Vet. J. 1979, 27, 217–220. [Google Scholar] [CrossRef]
- St. George, T.D.; Carmichael, L.E. Isolation of Mycoplasma Ovipneumoniae from Sheep with Chronic Pneumonia. Vet. Rec. 1975, 97, 205–206. [Google Scholar] [CrossRef]
- Fernández, S.; Galapero, J.; Rey, J.; Pérez, C.J.; Ramos, A.; Rosales, R.; Ayling, R.; Alonso, J.M.; Gómez, L. Investigations into the Seasonal Presence of Mycoplasma Species in Fattening Lambs. Vet. J. 2016, 212, 80–82. [Google Scholar] [CrossRef]
- Alley, M. The Effect of Chronic Non-Progressive Pneumonia on Weight Gain of Pasture-Fed Lambs. N. Z. Vet. J. 1987, 35, 163–166. [Google Scholar] [CrossRef]
- Jones, G.E.; Gilmour, J.S.; Rae, A.G. The Effect of Mycoplasma Ovipneumoniae and Pasteurella Haemolytica on Specific Pathogen-Free Lambs. J. Comp. Pathol. 1982, 92, 261–266. [Google Scholar] [CrossRef]
- Brogden, K.; Lehmkuhl, H.; Cutlip, R. Pasteurella Haemolytica Complicated Respiratory Infections in Sheep and Goats. Vet. Res. 1998, 29, 233–254. [Google Scholar]
- DaMassa, A.J.; Wakenell, P.S.; Brooks, D.L. Mycoplasmas of Goats and Sheep. J. Vet. Diagn. Invest. 1992, 4, 101–113. [Google Scholar] [CrossRef]
- Besser, T.E.; Levy, J.; Ackerman, M.; Nelson, D.; Manlove, K.; Potter, K.A.; Busboom, J.; Benson, M. A Pilot Study of the Effects of Mycoplasma Ovipneumoniae Exposure on Domestic Lamb Growth and Performance. PLoS ONE 2019, 14, e0207420. [Google Scholar] [CrossRef]
- Dassanayake, R.P.; Shanthalingam, S.; Herndon, C.N.; Subramaniam, R.; Lawrence, P.K.; Bavananthasivam, J.; Cassirer, E.F.; Haldorson, G.J.; Foreyt, W.J.; Rurangirwa, F.R.; et al. Mycoplasma Ovipneumoniae Can Predispose Bighorn Sheep to Fatal Mannheimia Haemolytica Pneumonia. Vet. Microbiol. 2010, 145, 354–359. [Google Scholar] [CrossRef]
- Li, Z.; Du, Z.; Li, J.; Sun, Y. Comparative Analysis on Lung Transcriptome of Mycoplasma Ovipneumoniae (Mo)-Infected Bashbay Sheep and Argali Hybrid Sheep. BMC Vet. Res. 2021, 17, 327. [Google Scholar] [CrossRef]
- Besser, T.E.; Cassirer, E.F.; Potter, K.A.; Lahmers, K.; Oaks, J.L.; Shanthalingam, S.; Srikumaran, S.; Foreyt, W.J. Epizootic Pneumonia of Bighorn Sheep Following Experimental Exposure to Mycoplasma Ovipneumoniae. PLoS ONE 2014, 9, e110039. [Google Scholar] [CrossRef]
- Niang, M.; Rosenbusch, R.F.; DeBey, M.C.; Niyo, Y.; Andrews, J.J.; Kaeberle, M.L. Field Isolates of Mycoplasma Ovipneumoniae Exhibit Distinct Cytopathic Effects in Ovine Tracheal Organ Cultures. J. Vet. Med. Ser. A 1998, 45, 29–40. [Google Scholar] [CrossRef]
- Jones, G.E.; Keir, W.A.; Gilmour, J.S. The Pathogenicity of Mycoplasma Ovipneumoniae and Mycoplasma Arginini in Ovine and Caprine Tracheal Organ Cultures. J. Comp. Pathol. 1985, 95, 477–487. [Google Scholar] [CrossRef]
- Niang, M.; Rosenbusch, R.F.; Andrews, J.J.; Lopez-Virella, J.; Kaeberle, M.L. Occurrence of Autoantibodies to Cilia in Lambs with a “Coughing Syndrome”. Vet. Immunol. Immunopathol. 1998, 64, 191–205. [Google Scholar] [CrossRef]
- Al-Kaissi, A.; Alley, M.R. Electron Microscopic Studies of the Interaction between Ovine Alveolar Macrophages and Mycoplasma Ovipneumoniae In Vitro. Vet. Microbiol. 1983, 8, 571–584. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, Y.; Zhu, E.; Yang, P.; Li, M.; Zhang, S.; Yue, J.; Wen, M.; Wang, K.; Cheng, Z. Mycoplasma Ovipneumoniae Induces Caspase-8-Dependent Extrinsic Apoptosis and P53- and ROS-Dependent Intrinsic Apoptosis in Murine Alveolar Macrophages. Virulence 2021, 12, 2703–2720. [Google Scholar] [CrossRef]
- Shahzad, W.; Ajuwape, A.T.P.; Rosenbusch, R.F. Global Suppression of Mitogen-Activated Ovine Peripheral Blood Mononuclear Cells by Surface Protein Activity from Mycoplasma Ovipneumoniae. Vet. Immunol. Immunopathol. 2010, 136, 116–121. [Google Scholar] [CrossRef]
- Einarsdottir, T.; Gunnarsson, E.; Hjartardottir, S. Icelandic Ovine Mycoplasma Ovipneumoniae Are Variable Bacteria That Induce Limited Immune Responses In Vitro and In Vivo. J. Med. Microbiol. 2018, 67, 1480–1490. [Google Scholar] [CrossRef]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef]
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a Novel Influenza Virus in Cattle and Swine: Proposal for a New Genus in the Orthomyxoviridae Family. mBio 2014, 5. [Google Scholar] [CrossRef]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B.M. Metagenomic Characterization of the Virome Associated with Bovine Respiratory Disease in Feedlot Cattle Identified Novel Viruses and Suggests an Etiologic Role for Influenza D Virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef]
- Ahlgren, E. Retrospective Serological and Virological Survey of Influenza D Virus among Cattle in Sweden. 2019. Available online: http://www.diva-portal.org/smash/get/diva2:1337747/FULLTEXT01.pdf (accessed on 26 February 2022).
- Ducatez, M.F.; Pelletier, C.; Meyer, G. Influenza D Virus in Cattle, France, 2011–2014. Emerg. Infect. Dis. 2015, 21, 368–371. [Google Scholar] [CrossRef]
- Oliva, J.; Eichenbaum, A.; Belin, J.; Gaudino, M.; Guillotin, J.; Alzieu, J.-P.; Nicollet, P.; Brugidou, R.; Gueneau, E.; Michel, E.; et al. Serological Evidence of Influenza D Virus Circulation among Cattle and Small Ruminants in France. Viruses 2019, 11, 516. [Google Scholar] [CrossRef]
- Sanogo, I.N.; Kouakou, C.; Batawui, K.; Djegui, F.; Byarugaba, D.K.; Adjin, R.; Adjabli, K.; Wabwire-Mangen, F.; Erima, B.; Atim, G.; et al. Serological Surveillance of Influenza D Virus in Ruminants and Swine in West and East Africa, 2017–2020. Viruses 2021, 13, 1749. [Google Scholar] [CrossRef]
- Gorin, S.; Fablet, C.; Quéguiner, S.; Barbier, N.; Paboeuf, F.; Hervé, S.; Rose, N.; Simon, G. Assessment of Influenza D Virus in Domestic Pigs and Wild Boars in France: Apparent Limited Spread within Swine Populations Despite Serological Evidence of Breeding Sow Exposure. Viruses 2019, 12, 25. [Google Scholar] [CrossRef]
- Nedland, H.; Wollman, J.; Sreenivasan, C.; Quast, M.; Singrey, A.; Fawcett, L.; Christopher-Hennings, J.; Nelson, E.; Kaushik, R.S.; Wang, D.; et al. Serological Evidence for the Co-Circulation of Two Lineages of Influenza D Viruses in Equine Populations of the Midwest United States. Zoonoses Public Health 2018, 65, e148–e154. [Google Scholar] [CrossRef]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological Evidence for the Presence of Influenza D Virus in Small Ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef]
- O’Donovan, T.; Donohoe, L.; Ducatez, M.F.; Meyer, G.; Ryan, E. Seroprevalence of Influenza D Virus in Selected Sample Groups of Irish Cattle, Sheep and Pigs. Ir. Vet. J. 2019, 72, 11. [Google Scholar] [CrossRef]
- Salem, E.; Cook, E.A.J.; Lbacha, H.A.; Oliva, J.; Awoume, F.; Aplogan, G.L.; Hymann, E.C.; Muloi, D.; Deem, S.L.; Alali, S.; et al. Serologic Evidence for Influenza C and D Virus among Ruminants and Camelids, Africa, 1991–2015. Emerg. Infect. Dis. 2017, 23, 1556–1559. [Google Scholar] [CrossRef]
- Zhai, S.-L.; Zhang, H.; Chen, S.-N.; Zhou, X.; Lin, T.; Liu, R.; Lv, D.-H.; Wen, X.-H.; Wei, W.-K.; Wang, D.; et al. Influenza D Virus in Animal Species in Guangdong Province, Southern China. Emerg. Infect. Dis. 2017, 23, 1392–1396. [Google Scholar] [CrossRef]
- Murakami, S.; Odagiri, T.; Melaku, S.K.; Bazartseren, B.; Ishida, H.; Takenaka-Uema, A.; Muraki, Y.; Sentsui, H.; Horimoto, T. Influenza D Virus Infection in Dromedary Camels, Ethiopia. Emerg. Infect. Dis. 2019, 25, 1224–1226. [Google Scholar] [CrossRef]
- Ferguson, L.; Luo, K.; Olivier, A.K.; Cunningham, F.L.; Blackmon, S.; Hanson-Dorr, K.; Sun, H.; Baroch, J.; Lutman, M.W.; Quade, B.; et al. Influenza D Virus Infection in Feral Swine Populations, United States. Emerg. Infect. Dis. 2018, 24, 1020–1028. [Google Scholar] [CrossRef]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic Evidence of Exposure to Influenza D Virus among Persons with Occupational Contact with Cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Marchi, S.; Manini, I.; Kistner, O.; Li, F.; Piu, P.; Manenti, A.; Biuso, F.; Sreenivasan, C.; Druce, J.; et al. Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses 2020, 12, 30. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Montomoli, E.; Di Bartolo, I.; Ostanello, F.; Chiapponi, C.; Marchi, S. Detection of Antibodies against Influenza D Virus in Swine Veterinarians in Italy in 2004. J. Med. Virol. 2021, 94, 2855–2859. [Google Scholar] [CrossRef]
- Oliva, J.; Mettier, J.; Sedano, L.; Delverdier, M.; Bourgès-Abella, N.; Hause, B.; Loupias, J.; Pardo, I.; Bleuart, C.; Bordignon, P.J.; et al. Murine Model for the Study of Influenza D Virus. J. Virol. 2020, 94, e01662-19. [Google Scholar] [CrossRef]
- Sreenivasan, C.; Thomas, M.; Sheng, Z.; Hause, B.M.; Collin, E.A.; Knudsen, D.E.B.; Pillatzki, A.; Nelson, E.; Wang, D.; Kaushik, R.S.; et al. Replication and Transmission of the Novel Bovine Influenza D Virus in a Guinea Pig Model. J. Virol. 2015, 89, 11990–12001. [Google Scholar] [CrossRef]
- Ferguson, L.; Olivier, A.K.; Genova, S.; Epperson, W.B.; Smith, D.R.; Schneider, L.; Barton, K.; McCuan, K.; Webby, R.J.; Wan, X.-F. Pathogenesis of Influenza D Virus in Cattle. J. Virol. 2016, 90, 5636–5642. [Google Scholar] [CrossRef]
- Mazzetto, E.; Bortolami, A.; Fusaro, A.; Mazzacan, E.; Maniero, S.; Vascellari, M.; Beato, M.S.; Schiavon, E.; Chiapponi, C.; Terregino, C.; et al. Replication of Influenza D Viruses of Bovine and Swine Origin in Ovine Respiratory Explants and Their Attachment to the Respiratory Tract of Bovine, Sheep, Goat, Horse, and Swine. Front. Microbiol. 2020, 11, 1136. [Google Scholar] [CrossRef]
- Nemanichvili, N.; Berends, A.J.; Wubbolts, R.W.; Gröne, A.; Rijks, J.M.; de Vries, R.P.; Verheije, M.H. Tissue Microarrays to Visualize Influenza D Attachment to Host Receptors in the Respiratory Tract of Farm Animals. Viruses 2021, 13, 586. [Google Scholar] [CrossRef]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. An Open Receptor-Binding Cavity of Hemagglutinin-Esterase-Fusion Glycoprotein from Newly-Identified Influenza D Virus: Basis for Its Broad Cell Tropism. PLoS Pathog. 2016, 12, e1005411. [Google Scholar] [CrossRef]
- Liu, R.; Sreenivasan, C.; Yu, H.; Sheng, Z.; Newkirk, S.J.; An, W.; Smith, D.F.; Chen, X.; Wang, D.; Li, F. Influenza D Virus Diverges from Its Related Influenza C Virus in the Recognition of 9-O-Acetylated N-Acetyl- or N-Glycolyl-Neuraminic Acid-Containing Glycan Receptors. Virology 2020, 545, 16–23. [Google Scholar] [CrossRef]
- Zhang, X.; Outlaw, C.; Olivier, A.K.; Woolums, A.; Epperson, W.; Wan, X.-F. Pathogenesis of Co-Infections of Influenza D Virus and Mannheimia Haemolytica in Cattle. Vet. Microbiol. 2019, 231, 246–253. [Google Scholar] [CrossRef]
- Zhang, M.; Hill, J.E.; Fernando, C.; Alexander, T.W.; Timsit, E.; van der Meer, F.; Huang, Y. Respiratory Viruses Identified in Western Canadian Beef Cattle by Metagenomic Sequencing and Their Association with Bovine Respiratory Disease. Transbound. Emerg. Dis. 2019, 66, 1379–1386. [Google Scholar] [CrossRef]
- Mekata, H.; Yamamoto, M.; Hamabe, S.; Tanaka, H.; Omatsu, T.; Mizutani, T.; Hause, B.M.; Okabayashi, T. Molecular Epidemiological Survey and Phylogenetic Analysis of Bovine Influenza D Virus in Japan. Transbound. Emerg. Dis. 2018, 65, e355–e360. [Google Scholar] [CrossRef]
- Salem, E.; Hägglund, S.; Cassard, H.; Corre, T.; Näslund, K.; Foret, C.; Gauthier, D.; Pinard, A.; Delverdier, M.; Zohari, S.; et al. Pathogenesis, Host Innate Immune Response, and Aerosol Transmission of Influenza D Virus in Cattle. J. Virol. 2019, 93, e01853-18. [Google Scholar] [CrossRef]
- Ng, T.F.F.; Kondov, N.O.; Deng, X.; Van Eenennaam, A.; Neibergs, H.L.; Delwart, E. A Metagenomics and Case-Control Study to Identify Viruses Associated with Bovine Respiratory Disease. J. Virol. 2015, 89, 5340–5349. [Google Scholar] [CrossRef]
- Nissly, R.H.; Zaman, N.; Ibrahim, P.A.S.; McDaniel, K.; Lim, L.; Kiser, J.N.; Bird, I.; Chothe, S.K.; Bhushan, G.L.; Vandegrift, K.; et al. Influenza C and D Viral Load in Cattle Correlates with Bovine Respiratory Disease (BRD): Emerging Role of Orthomyxoviruses in the Pathogenesis of BRD. Virology 2020, 551, 10–15. [Google Scholar] [CrossRef]
- Lion, A.; Secula, A.; Rançon, C.; Boulesteix, O.; Pinard, A.; Deslis, A.; Hägglund, S.; Salem, E.; Cassard, H.; Näslund, K.; et al. Enhanced Pathogenesis Caused by Influenza D Virus and Mycoplasma Bovis Coinfection in Calves: A Disease Severity Linked with Overexpression of IFN-γ as a Key Player of the Enhanced Innate Immune Response in Lungs. Microbiol. Spectr. 2021, 9, e01690-21. [Google Scholar] [CrossRef]
- Foggie, A.; Jones, G.E.; Buxton, D. The Experimental Infection of Specific Pathogen Free Lambs with Mycoplasma Ovipneumoniae. Res. Vet. Sci. 1976, 21, 28–35. [Google Scholar] [CrossRef]
- Jones, G.E.; Gilmour, J.S.; Rae, A.G. The Effects of Different Strains of Mycoplasma Ovipneumoniae on Specific Pathogen-Free and Conventionally-Reared Lambs. J. Comp. Pathol. 1982, 92, 267–272. [Google Scholar] [CrossRef]
- Brown, M.R.; Hands, C.L.; Coello-Garcia, T.; Sani, B.S.; Ott, A.I.G.; Smith, S.J.; Davenport, R.J. A Flow Cytometry Method for Bacterial Quantification and Biomass Estimates in Activated Sludge. J. Microbiol. Methods 2019, 160, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Littlefield, K. Evaluating the Zoonotic Potential of Non-Human Influenza D Virus. Master’s Thesis, Johns Hopkins University, Baltimore, MD, USA, 2021. [Google Scholar]
- Ishida, H.; Murakami, S.; Kamiki, H.; Matsugo, H.; Takenaka-Uema, A.; Horimoto, T. Establishment of a Reverse Genetics System for Influenza D Virus. J. Virol. 2020, 94, e01767-19. [Google Scholar] [CrossRef]
- Klebe, R.J.; Harriss, J.V. A Technically Simple “Non-Lethal” Vital Staining Procedure for Viral Plaque and Cell Transformation Assays. Brief Report. Arch. Virol. 1984, 81, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, H.; Zhao, N.; Wang, J.; Yang, Y.; Sun, Y. Therapeutic Effects of Recombinant SPLUNC1 on Mycoplasma Ovipneumoniae-Infected Argali Hybrid Sheep. Res. Vet. Sci. 2020, 133, 174–179. [Google Scholar] [CrossRef]
- Du, Z.; Sun, Y.; Wang, J.; Liu, H.; Yang, Y.; Zhao, N. Comprehensive RNA-Seq Profiling of the Lung Transcriptome of Bashbay Sheep in Response to Experimental Mycoplasma Ovipneumoniae Infection. PLoS ONE 2020, 15, e0214497. [Google Scholar] [CrossRef]
- Khan, M.A. Respiratory Infection of Lambs with Mycoplasma Ovipneumoniae. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 1993. [Google Scholar]
- Kaplan, B.S.; Falkenberg, S.; Dassanayake, R.; Neill, J.; Velayudhan, B.; Li, F.; Vincent, A.L. Virus Strain Influenced the Interspecies Transmission of Influenza D Virus between Calves and Pigs. Transbound. Emerg. Dis. 2021, 68, 3396–3404. [Google Scholar] [CrossRef]
- Yang, F.; Dao, X.; Rodriguez-Palacios, A.; Feng, X.; Tang, C.; Yang, X.; Yue, H. A Real-Time PCR for Detection and Quantification of Mycoplasma Ovipneumoniae. J. Vet. Med. Sci. 2014, 76, 1631–1634. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Britton, G.J.; Chen-Liaw, A.; Cossarini, F.; Livanos, A.E.; Spindler, M.P.; Plitt, T.; Eggers, J.; Mogno, I.; Gonzalez-Reiche, A.S.; Siu, S.; et al. Limited Intestinal Inflammation despite Diarrhea, Fecal Viral RNA and SARS-CoV-2-Specific IgA in Patients with Acute COVID-19. Sci. Rep. 2021, 11, 13308. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, L.; Syedbasha, M.; Vogt, D.; Hollenstein, Y.; Hartmann, J.; Linnik, J.E.; Egli, A. An Optimized Hemagglutination Inhibition (HI) Assay to Quantify Influenza-Specific Antibody Titers. J. Vis. Exp. 2017, 55833. [Google Scholar] [CrossRef] [PubMed]
- Table: Normal Rectal Temperature Ranges. Available online: https://www.merckvetmanual.com/multimedia/table/normal-rectal-temperature-ranges (accessed on 19 April 2022).
- Chiapponi, C.; Faccini, S.; De Mattia, A.; Baioni, L.; Barbieri, I.; Rosignoli, C.; Nigrelli, A.; Foni, E. Detection of Influenza D Virus among Swine and Cattle, Italy. Emerg. Infect. Dis. 2016, 22, 352–354. [Google Scholar] [CrossRef]
- Guan, M.; Blackmon, S.; Olivier, A.K.; Zhang, X.; Liu, L.; Woolums, A.; Crenshaw, M.A.; Liao, S.F.; Webby, R.; Epperson, W.; et al. Time-Dependent Proinflammatory Responses Shape Virus Interference during Coinfections of Influenza A Virus and Influenza D Virus. Viruses 2022, 14, 224. [Google Scholar] [CrossRef] [PubMed]
- Ionas, G.; Mew, A.J.; Alley, M.R.; Clarke, J.K.; Robinson, A.J.; Marshall, R.B. Colonisation of the Respiratory Tract of Lambs by Strains of Mycoplasma Ovipneumoniae. Vet. Microbiol. 1985, 10, 533–539. [Google Scholar] [CrossRef]
- Thirkell, D.; Spooner, R.K.; Jones, G.E.; Russell, W.C. The Humoral Immune Response of Lambs Experimentally Infected with Mycoplasma Ovipneumoniae. Vet. Microbiol. 1990, 24, 143–153. [Google Scholar] [CrossRef]
- Buddle, B.M.; Herceg, M.; Davies, D.H. Experimental Infection of Sheep with Mycoplasma Ovipneumoniae and Pasteurella Haemolytica. Vet. Microbiol. 1984, 9, 543–548. [Google Scholar] [CrossRef]
- Niang, M.; Rosenbusch, R.F.; Andrews, J.J.; Kaeberle, M.L. Demonstration of a Capsule on Mycoplasma Ovipneumoniae. Am. J. Vet. Res. 1998, 59, 557–562. [Google Scholar]
- Saegerman, C.; Salem, E.; Ait Lbacha, H.; Alali, S.; Zouagui, Z.; Meyer, G.; Ducatez, M.F. Formal Estimation of the Seropositivity Cut-off of the Hemagglutination Inhibition Assay in Field Diagnosis of Influenza D Virus in Cattle and Estimation of the Associated True Prevalence in Morocco. Transbound. Emerg. Dis. 2021, 68, 1392–1399. [Google Scholar] [CrossRef]
- Ionas, G.; Norman, N.G.; Clarke, J.K.; Marshall, R.B. A Study of the Heterogeneity of Isolates of Mycoplasma Ovipneumoniae from Sheep in New Zealand. Vet. Microbiol. 1991, 29, 339–347. [Google Scholar] [CrossRef]
- He, W.-T.; Lu, M.; Xing, G.; Shao, Y.; Zhang, M.; Yang, Y.; Li, X.; Zhang, L.; Li, G.; Cao, Z.; et al. Emergence and Adaptive Evolution of Influenza D Virus. Microb. Pathog. 2021, 160, 105193. [Google Scholar] [CrossRef] [PubMed]
- Saegerman, C.; Gaudino, M.; Savard, C.; Broes, A.; Ariel, O.; Meyer, G.; Ducatez, M.F. Influenza D Virus in Respiratory Disease in Canadian, Province of Québec, Cattle: Relative Importance and Evidence of New Reassortment between Different Clades. Transbound. Emerg. Dis. 2021, 69, 1227–1245. [Google Scholar] [CrossRef] [PubMed]
- Nemanichvili, N.; Berends, A.J.; Tomris, I.; Barnard, K.N.; Parrish, C.R.; Gröne, A.; Rijks, J.M.; Verheije, M.H.; de Vries, R.P. Influenza D Binding Properties Vary amongst the Two Major Virus Clades and Wildlife Species. Vet. Microbiol. 2022, 264, 109298. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPI (M. ovi) | DPI (IDV) | sal-IDV (SD) | Mo-IDV (SD) | |
|---|---|---|---|---|
| Attitude a | 2 | - | 0 (0) | 0.14 (0.38) |
| 3 | - | 0.14 (0.38) | 0 (0) | |
| 14 | - | 0 (0) | 0.17 (0.41) | |
| 21 | - | 0 (0) | 0.14 (0.38) | |
| 23 | - | 0 (0) | 0.14 (0.38) | |
| 42 | 14 | 0 (0) | 0.14 (0.38) | |
| Body Condition b | 2 | - | 3.29 (0.27) | 3.36 (0.24) |
| 9 | - | 3.29 (0.39) | 3.29 (0.39) | |
| 16 | - | 3.21 (0.27) | 3.07 (0.35) | |
| 23 | - | 3.36 (0.38) | 3.57 (0.45) | |
| 30 | 2 | 3.64 (0.38) | 3.5 (0.29) | |
| 37 | 9 | 3.86 (0.24) | 3.79 (0.39) | |
| 44 | 16 | 3.42 (0.2) | 3.57 (0.35) | |
| Clinical Signs c | 2 | - | 0 (0) | 0.14 (0.38) |
| 3 | - | 0.14 (0.38) | 0 (0) | |
| 23 | - | 0 (0) | 0.14 (0.38) | |
| 25 | - | 0.14 (0.38) | 0 (0) | |
| 27 | - | 0 (0) | 0.14 (0.38) | |
| 28 | 0 | 0 (0) | 0.14 (0.38) | |
| 28 | 0 | 0 (0) | 0.14 (0.38) |
| Timepoint | sal-IDV | Mo-IDV | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean | Median | Range | Mean | Median | Range | p | ||
| Total Serum Protein (g/dL) | Start | 7.66 | 7.3 | 6.2–8.8 | 7.6 | 7.6 | 6.5–8.7 | >0.999 |
| Day-1 | 10.8 | 11 | 9.1–11.7 | 9.6 | 9.7 | 9.1–10.1 | 0.244 | |
| Day 6 | 10.4 | 9.9 | 7.8–13.1 | 9 | 9.5 | 6.8–10 | 0.236 | |
| Day 14 | 10.4 | 10.3 | 8–13.5 | 11.6 | 11.4 | 10.5–14.2 | 0.581 | |
| Endpoint | 10.5 | 10.7 | 8.6–12.2 | 10.7 | 10.5 | 8.8–13 | >0.999 | |
| Albumin (g/dL) | Start | 4.2 | 4.2 | 3.9–4.4 | 4.2 | 4.3 | 3.3–4.7 | >0.999 |
| Day-1 | 4.9 | 4.9 | 4.6–5.5 | 4.8 | 4.8 | 3.3–5.9 | >0.999 | |
| Day 6 | 4.9 | 4.9 | 4.7–5.3 | 4.6 | 4.8 | 3.3–5.5 | >0.999 | |
| Day 14 | 4.4 | 4.4 | 3.9–4.6 | 4.4 | 4.4 | 3.6–5.2 | >0.999 | |
| Endpoint | 4.6 | 4.6 | 4.4–4.7 | 4.2 | 4.3 | 3.1–5 | 0.546 | |
| SAA (ng/mL) | Day-1 | 618.5 | 58.5 | 3.4–2902.2 | 499.7 | 42.4 | 0–2936.8 | >0.999 |
| Day 6 | 158.4 | 30.8 | 0–838 | 667.3 | 231.2 | 36.2–2793.5 | 0.587 | |
| L-lactate (mM) | Day-1 | 1.9 | 2 | 1.4–2.2 | 1.7 | 1.8 | 1.1–2.3 | 0.495 |
| Day 6 | 1.8 | 1.9 | 0.3–3 | 1.9 | 1.8 | 0.8–2.8 | 0.838 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, E.; Schulein, C.; Jacobson, B.T.; Jones, K.; Sago, J.; Huber, V.; Jutila, M.; Bimczok, D.; Rynda-Apple, A. Pathophysiology of Influenza D Virus Infection in Specific-Pathogen-Free Lambs with or without Prior Mycoplasma ovipneumoniae Exposure. Viruses 2022, 14, 1422. https://doi.org/10.3390/v14071422
Robinson E, Schulein C, Jacobson BT, Jones K, Sago J, Huber V, Jutila M, Bimczok D, Rynda-Apple A. Pathophysiology of Influenza D Virus Infection in Specific-Pathogen-Free Lambs with or without Prior Mycoplasma ovipneumoniae Exposure. Viruses. 2022; 14(7):1422. https://doi.org/10.3390/v14071422
Chicago/Turabian StyleRobinson, Ema, Clyde Schulein, B. Tegner Jacobson, Kerri Jones, Jonathon Sago, Victor Huber, Mark Jutila, Diane Bimczok, and Agnieszka Rynda-Apple. 2022. "Pathophysiology of Influenza D Virus Infection in Specific-Pathogen-Free Lambs with or without Prior Mycoplasma ovipneumoniae Exposure" Viruses 14, no. 7: 1422. https://doi.org/10.3390/v14071422
APA StyleRobinson, E., Schulein, C., Jacobson, B. T., Jones, K., Sago, J., Huber, V., Jutila, M., Bimczok, D., & Rynda-Apple, A. (2022). Pathophysiology of Influenza D Virus Infection in Specific-Pathogen-Free Lambs with or without Prior Mycoplasma ovipneumoniae Exposure. Viruses, 14(7), 1422. https://doi.org/10.3390/v14071422