
Targeted Therapy of HPV Positive and Negative Tonsillar Squamous Cell Carcinoma Cell Lines Reveals Synergy between CDK4/6, PI3K and Sometimes FGFR Inhibitors, but Rarely between PARP and WEE1 Inhibitors

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Seeding
2.2. Drugs, and Treatments
2.3. WST-1 Viability Assay
2.4. Cell Proliferation and Cytotoxicity Assays
2.5. Statistical Analysis
3. Results
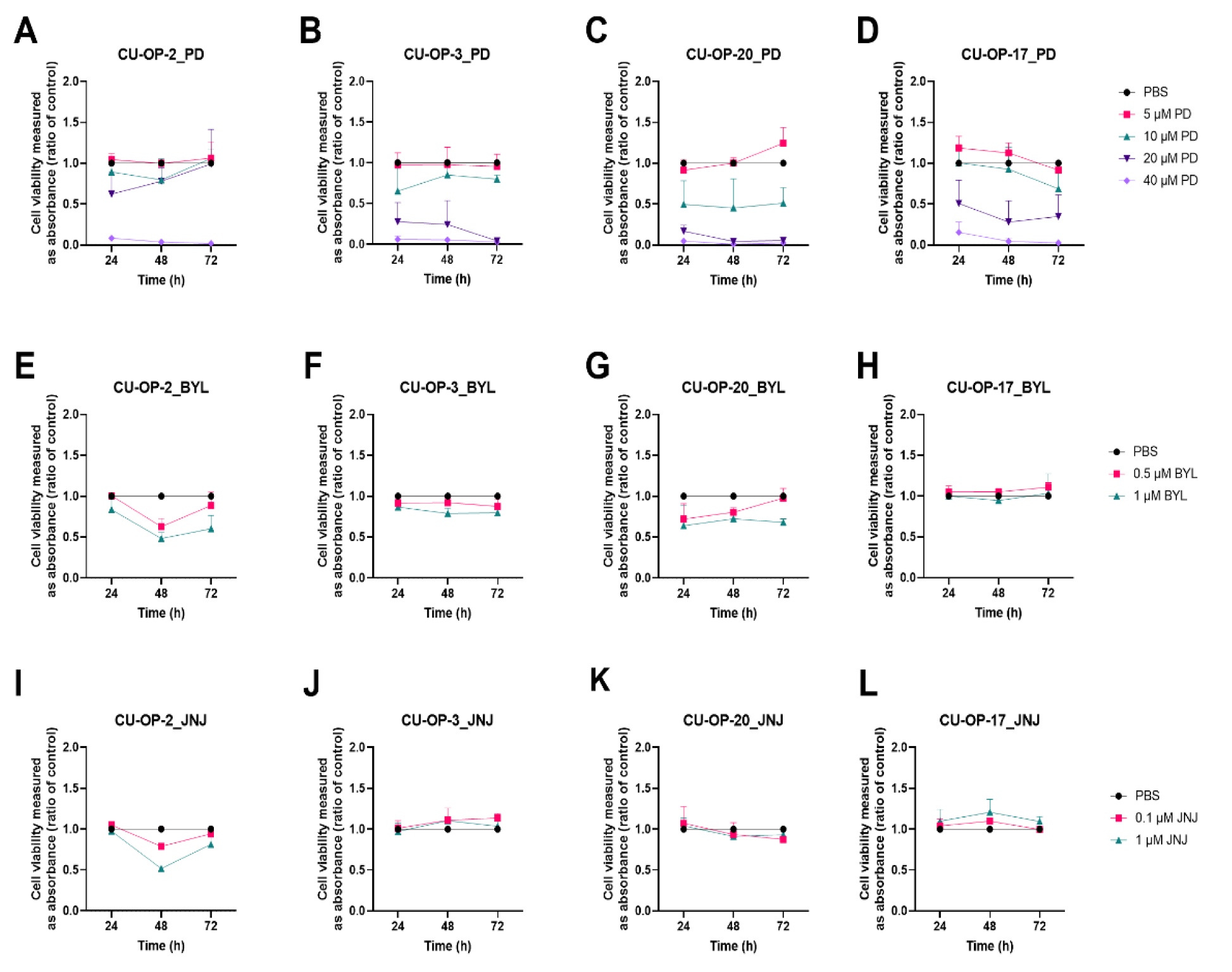
3.1. Viability after Single Exposure to FDA Approved CDK4/6, PI3K, FGFR, PARP Inhibitors (PD-332991, BYL719, JNJ-42756493, BMN-673 Resp) and a WEE-1 Inhibitor (MK-1775) Measured by WST-1 Assays in HPV+ and HPV− TSCC Lines
3.1.1. PD-0332991
3.1.2. BYL719
3.1.3. JNJ-42756493
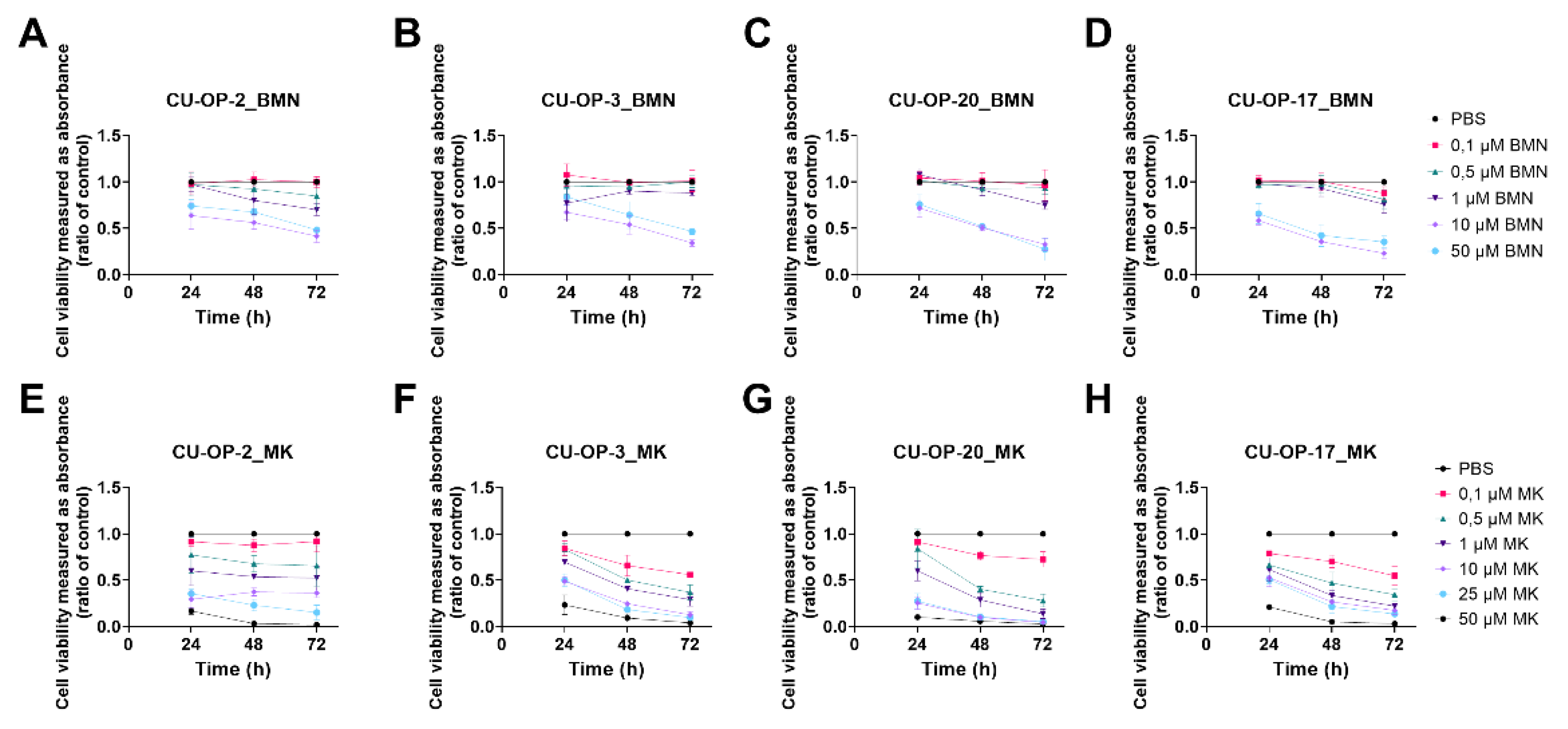
3.1.4. BMN-673
3.1.5. MK1775
3.1.6. IC50 Values of PD-0332991, BMN-673, and MK-1775 Inhibitors on the CU-OP Cell Lines
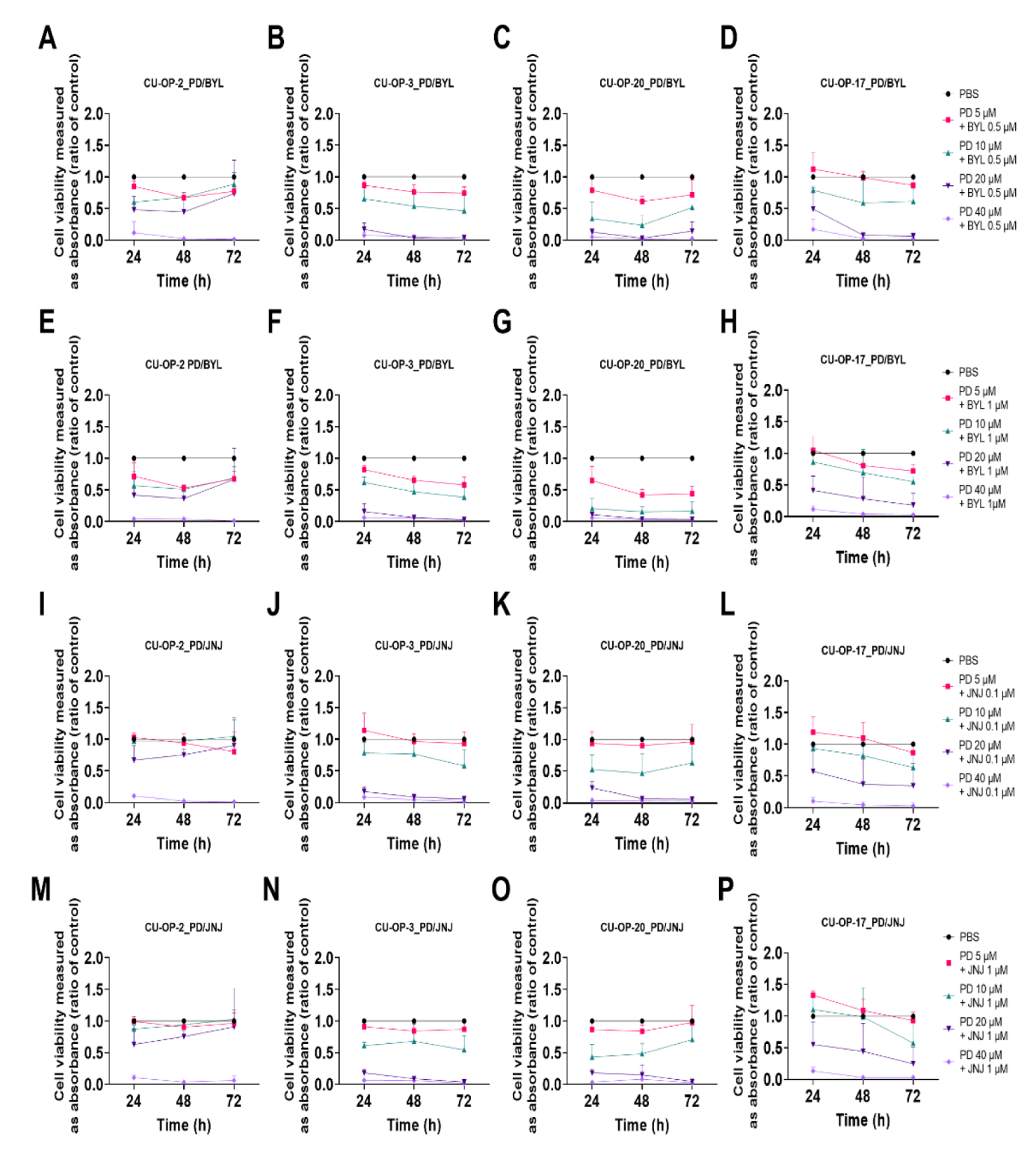
3.2. Viability after Combined Exposure of FDA Approved CDK 4/6, PI3K, and FGFR Inhibitors (PD-0332991, BYL719, and JNJ-42756493 Resp.) in HPV+ and HPV− TSCC Cell Lines
3.2.1. PD-0332991 and BYL719
3.2.2. PD-0332991 and JNJ-42756493
3.2.3. Combinational Indexes with the “Highest Single Agent” Approach
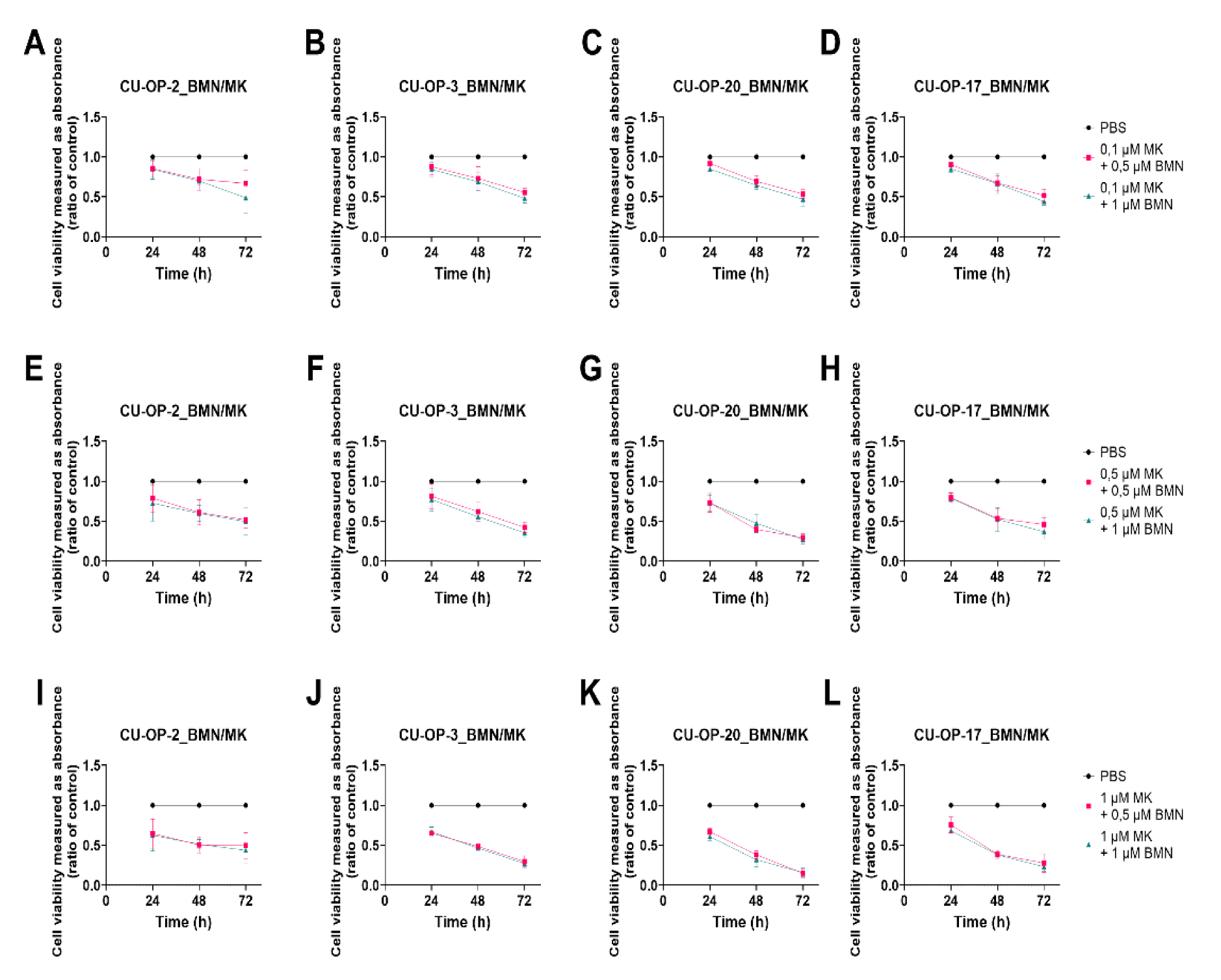
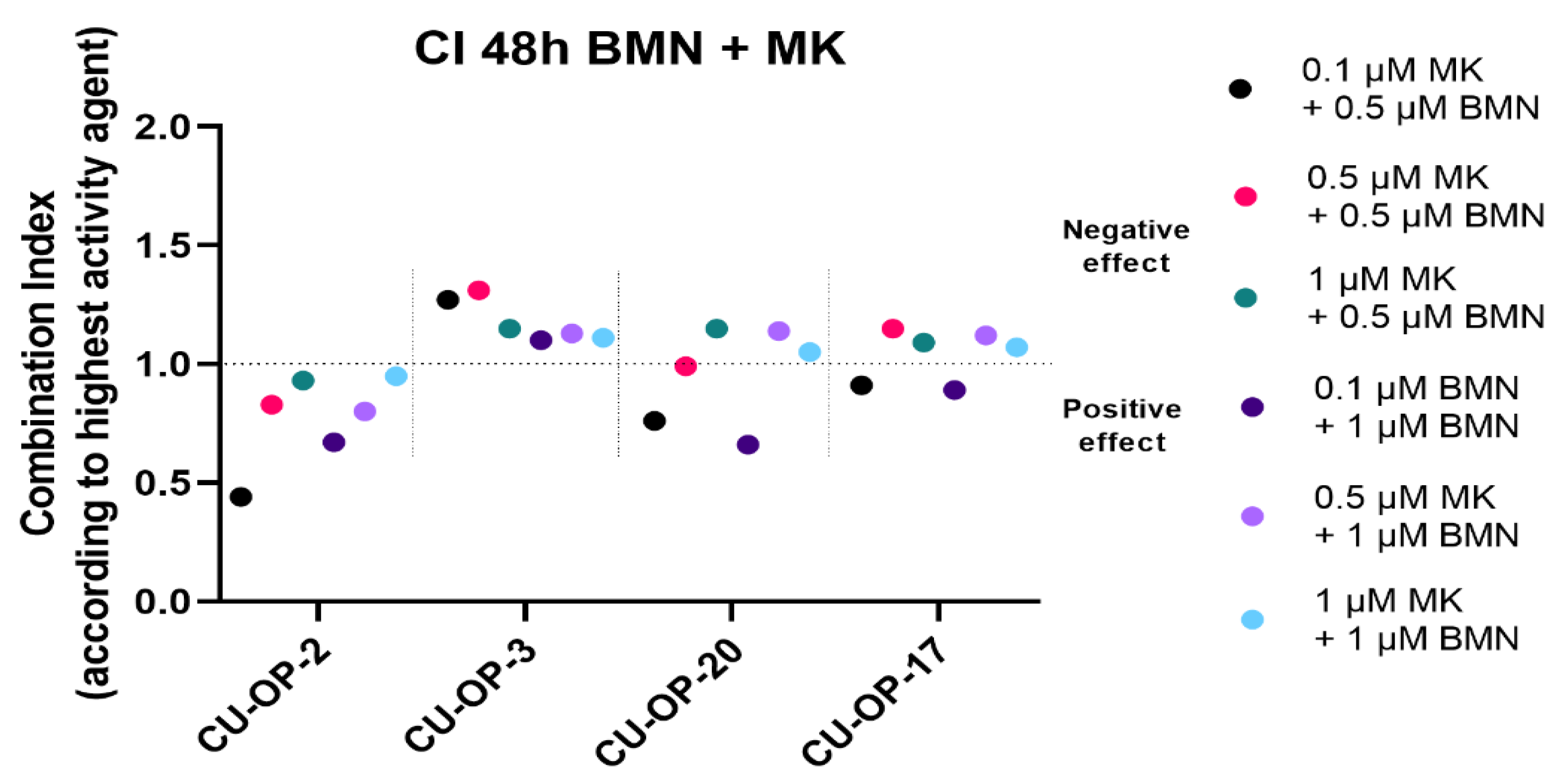
3.3. Viability after Combined Exposure of an FDA Approved PARP and a WEE1 Inhibitor (BMN-673 and MK-1775 Resp), in HPV+ and HPV− TSCC Cell Lines
3.3.1. BMN-673 and MK-1775
3.3.2. Combinational Indexes with the “Highest Single Agent” Approach
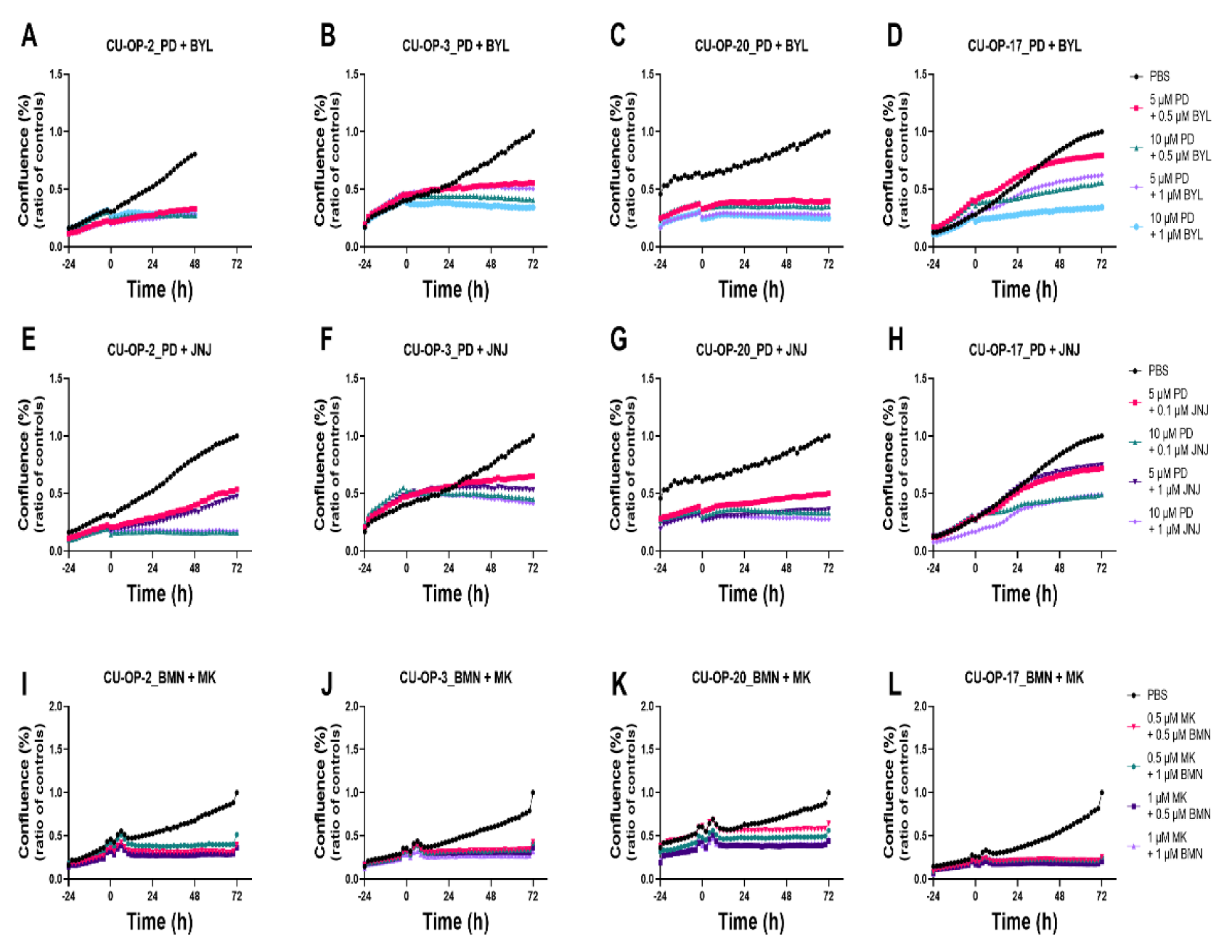
3.4. Proliferation and Cytotoxicity Responses after Single Treatments with FDA Approved CDK4/6, PI3K, FGFR, PARP, and a WEE1 Inhibitors (PD-0332991, BYL719, JNJ-42756493, BMN-673, and MK-1775 Resp.) of HPV+ and HPV− TSCC Cell Lines
3.4.1. Proliferation
3.4.2. Cytotoxicity
3.5. Proliferation and Cytotoxicity Responses after Combined Treatments with FDA Approved CDK4/6, PI3K, and FGFR Inhibitors (PD-0332991, BYL719, JNJ-42756493) and of PARP and WEE1 Inhibitors (BMN-673 and MK-1775) of HPV+ and HPV− TSCC Cell Lines
3.5.1. Proliferation
3.5.2. Cytotoxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mellin, H.; Friesland, S.; Lewensohn, R.; Dalianis, T.; Munck-Wikland, E. Human Papillomavirus (Hpv) DNA in Tonsillar Cancer: Clinical Correlates, Risk of Relapse, and Survival. Int. J. Cancer 2000, 89, 300–304. [Google Scholar] [CrossRef]
- Dahlgren, L.; Dahlstrand, H.; Lindquist, D.; Högmo, A.; Björnestål, L.; Lindholm, J.; Lundberg, B.; Dalianis, T.; Munck-Wikland, E. Human papillomavirus is more common in base of tongue than in mobile tongue cancer and is a favorable prognostic factor in base of tongue cancer patients. Int. J. Cancer 2004, 112, 1015–1019. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, D.; Romanitan, M.; Hammarstedt-Nordenvall, L.; Näsman, A.; Dahlstrand, H.; Lindholm, J.; Onelöv, L.; Ramqvist, T.; Ye, W.; Munck-Wikland, E.; et al. Human papillomavirus is a favourable prognostic factor in tonsillar cancer and its oncogenic role is supported by the expression of E6 and E7. Mol. Oncol. 2007, 1, 350–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlstrand, H.; Näsman, A.; Romanitan, M.; Lindquist, D.; Ramqvist, T.; Dalianis, T. Human papillomavirus accounts both for increased incidence and better prognosis in tonsillar cancer. Anticancer. Res. 2008, 28, 1133–1138. [Google Scholar] [PubMed]
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tân, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human Papillomavirus and Survival of Patients with Oropharyngeal Cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Näsman, A.; Attner, P.; Hammarstedt-Nordenvall, L.; Du, J.; Eriksson, M.; Giraud, G.; Ährlund-Richter, S.; Marklund, L.; Romanitan, M.; Lindquist, D.; et al. Incidence of human papillomavirus (HPV) positive tonsillar carcinoma in Stockholm, Sweden: An epidemic of viral-induced carcinoma? Int. J. Cancer 2009, 125, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Haeggblom, L.; Attoff, T.; Yu, J.; Holzhauser, S.; Vlastos, A.; Mirzae, L.; Msci, A.; Ahrlund-Richter, A.; Munck-Wikland, E.; Marklund, L.; et al. Changes in incidence and prevalence of human papillomavirus in tonsillar and base of tongue cancer during 2000–2016 in the Stockholm region and Sweden. Head Neck 2019, 41, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Ernster, J.A.; Sciotto, C.G.; O’Brien, M.M.; Finch, J.L.; Robinson, L.J.; Willson, T.; Mathews, M. Rising Incidence of Oropharyngeal Cancer and the Role of Oncogenic Human Papilloma Virus. Laryngoscope 2007, 117, 2115–2128. [Google Scholar] [CrossRef]
- Chaturvedi, A.K.; Engels, E.A.; Pfeiffer, R.M.; Hernandez, B.Y.; Xiao, W.; Kim, E.; Jiang, B.; Goodman, M.T.; Sibug-Saber, M.; Cozen, W.; et al. Human Papillomavirus and Rising Oropharyngeal Cancer Incidence in the United States. J. Clin. Oncol. 2011, 29, 4294–4301. [Google Scholar] [CrossRef]
- Garnaes, E.; Kiss, K.; Andersen, L.; Therkildsen, M.H.; Franzmann, M.B.; Filtenborg-Barnkob, B.; Hoegdall, E.; Krenk, L.; Josiassen, M.; Lajer, C.B.; et al. A high and increasing HPV prevalence in tonsillar cancers in Eastern Denmark, 2000-2010: The largest registry-based study to date. Int. J. Cancer 2014, 136, 2196–2203. [Google Scholar] [CrossRef]
- Pytynia, K.B.; Dahlstrom, K.R.; Sturgis, E.M. Epidemiology of HPV-associated oropharyngeal cancer. Oral Oncol. 2014, 50, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Licitra, L.; Bernier, J.; Grandi, C.; Merlano, M.; Bruzzi, P.; Lefebvre, J.-L. Cancer of the oropharynx. Crit. Rev. Oncol. 2002, 41, 107–122. [Google Scholar] [CrossRef]
- Oropharyngeal Cancer Treatment (Adult) (Pdq®)–Health Professional Version. In PDQ Cancer Information Summaries; National Cancer Institute: Bethesda, MD, USA, 2002.
- Bersani, C.; Mints, M.; Tertipis, N.; Haeggblom, L.; Sivars, L.; Ährlund-Richter, A.; Vlastos, A.; Smedberg, C.; Grün, N.; Munck-Wikland, E.; et al. A model using concomitant markers for predicting outcome in human papillomavirus positive oropharyngeal cancer. Oral Oncol. 2017, 68, 53–59. [Google Scholar] [CrossRef]
- Strohl, M.P.; Wai, K.C.; Ha, P.K. De-intensification strategies in HPV-related oropharyngeal squamous cell carcinoma—a narrative review. Ann. Transl. Med. 2020, 8, 1601. [Google Scholar] [CrossRef]
- Näsman, A.; Holzhauser, S.; Kostopoulou, O.; Zupancic, M.; Ährlund-Richter, A.; Du, J.; Dalianis, T. Prognostic Markers and Driver Genes and Options for Targeted Therapy in Human-Papillomavirus-Positive Tonsillar and Base-of-Tongue Squamous Cell Carcinoma. Viruses 2021, 13, 910. [Google Scholar] [CrossRef]
- Näsman, A.; Nordfors, C.; Grün, N.; Munck-Wikland, E.; Ramqvist, T.; Marklund, L.; Lindquist, D.; Dalianis, T. Absent/weak CD 44 intensity and positive human papillomavirus (HPV) status in oropharyngeal squamous cell carcinoma indicates a very high survival. Cancer Med. 2013, 2, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Näsman, A.; Andersson, E.; Marklund, L.; Tertipis, N.; Hammarstedt-Nordenvall, L.; Attner, P.; Nyberg, T.; Masucci, G.V.; Munck-Wikland, E.; Ramqvist, T.; et al. HLA Class I and II Expression in Oropharyngeal Squamous Cell Carcinoma in Relation to Tumor HPV Status and Clinical Outcome. PLoS ONE 2013, 8, e77025. [Google Scholar] [CrossRef] [Green Version]
- Tertipis, N.; Haeggblom, L.; Nordfors, C.; Grün, N.; Näsman, A.; Vlastos, A.; Dalianis, T.; Ramqvist, T. Correlation of LMP10 Expression and Clinical Outcome in Human Papillomavirus (HPV) Positive and HPV-Negative Tonsillar and Base of Tongue Cancer. PLoS ONE 2014, 9, e95624. [Google Scholar] [CrossRef] [Green Version]
- Rietbergen, M.M.; Kemp, S.M.-D.; Bloemena, E.; Witte, B.I.; Brink, A.; de Jong, R.B.; Leemans, C.R.; Braakhuis, B.J.; Brakenhoff, R.H. Cancer stem cell enrichment marker CD98: A prognostic factor for survival in patients with human papillomavirus-positive oropharyngeal cancer. Eur. J. Cancer 2014, 50, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Nordfors, C.; Grün, N.; Tertipis, N.; Ährlund-Richter, A.; Haeggblom, L.; Sivars, L.; Du, J.; Nyberg, T.; Marklund, L.; Munck-Wikland, E.; et al. CD8+ and CD4+ tumour infiltrating lymphocytes in relation to human papillomavirus status and clinical outcome in tonsillar and base of tongue squamous cell carcinoma. Eur. J. Cancer 2013, 49, 2522–2530. [Google Scholar] [CrossRef]
- Oguejiofor, K.; Hall, J.; Slater, C.; Betts, G.; Hall, G.; Slevin, N.; Dovedi, S.; Stern, P.L.; West, C.M.L. Stromal infiltration of CD8 T cells is associated with improved clinical outcome in HPV-positive oropharyngeal squamous carcinoma. Br. J. Cancer 2015, 113, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Oguejiofor, K.; Galletta-Williams, H.; Dovedi, S.J.; Roberts, D.L.; Stern, P.L.; West, C.M. Distinct patterns of infiltrating CD8+ T cells in HPV+ and CD68 macrophages in HPV- oropharyngeal squamous cell carcinomas are associated with better clinical outcome but PD-L1 expression is not prognostic. Oncotarget 2017, 8, 14416–14427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindquist, D.; Näsman, A.; Tarjan, M.; Henriksson, R.; Tot, T.; Dalianis, T.; Hedman, H. Expression of LRIG1 is associated with good prognosis and human papillomavirus status in oropharyngeal cancer. Br. J. Cancer 2014, 110, 1793–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lechner, M.; Frampton, G.M.; Fenton, T.; Feber, A.; Palmer, G.; Jay, A.; Pillay, N.; Forster, M.; Cronin, M.T.; Lipson, D.; et al. Targeted next-generation sequencing of head and neck squamous cell carcinoma identifies novel genetic alterations in HPV+ and HPV- tumors. Genome Med. 2013, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Tinhofer, I.; Budach, V.; Saki, M.; Konschak, R.; Niehr, F.; Jöhrens, K.; Weichert, W.; Linge, A.; Lohaus, F.; Krause, M.; et al. Targeted next-generation sequencing of locally advanced squamous cell carcinomas of the head and neck reveals druggable targets for improving adjuvant chemoradiation. Eur. J. Cancer 2016, 57, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Bersani, C.; Sivars, L.; Haeggblom, L.; DiLorenzo, S.; Mints, M.; Ährlund-Richter, A.; Tertipis, N.; Munck-Wikland, E.; Näsman, A.; Ramqvist, T.; et al. Targeted sequencing of tonsillar and base of tongue cancer and human papillomavirus positive unknown primary of the head and neck reveals prognostic effects of mutated FGFR3. Oncotarget 2017, 8, 35339–35350. [Google Scholar] [CrossRef]
- Ährlund-Richter, A.; Holzhauser, S.; Dalianis, T.; Näsman, A.; Mints, M. Whole-Exome Sequencing of HPV Positive Tonsillar and Base of Tongue Squamous Cell Carcinomas Reveals a Global Mutational Pattern along with Relapse-Specific Somatic Variants. Cancers 2021, 14, 77. [Google Scholar] [CrossRef]
- Bersani, C.; Haeggblom, L.; Ursu, R.G.; Giusca, S.E.; Marklund, L.; Ramqvist, T.; Näsman, A.; Dalianis, T. Overexpression of FGFR3 in HPV-positive Tonsillar and Base of Tongue Cancer Is Correlated to Outcome. Anticancer Res. 2018, 38, 4683–4690. [Google Scholar] [CrossRef]
- Koole, K.; van Kempen, P.M.W.; Swartz, J.E.; Peeters, T.; van Diest, P.J.; Koole, R.; van Es, R.J.J.; Willems, S.M. Fibroblast growth factor receptor 3 protein is overexpressed in oral and oropharyngeal squamous cell carcinoma. Cancer Med. 2016, 5, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Beaty, B.T.; Moon, D.H.; Shen, C.J.; Amdur, R.J.; Weiss, J.; Grilley-Olson, J.; Patel, S.; Zanation, A.; Hackman, T.G.; Thorp, B.; et al. PIK3CA Mutation in HPV-Associated OPSCC Patients Receiving Deintensified Chemoradiation. JNCI J. Natl. Cancer Inst. 2020, 112, 855–858. [Google Scholar] [CrossRef]
- André, F.; Ciruelos, E.; Rubovszky, G.; Campone, M.; Loibl, S.; Rugo, H.S.; Iwata, H.; Conte, P.; Mayer, I.A.; Kaufman, B.; et al. Alpelisib for PIK3CA-Mutated, Hormone Receptor–Positive Advanced Breast Cancer. N. Engl. J. Med. 2019, 380, 1929–1940. [Google Scholar] [CrossRef]
- Bahleda, R.; Italiano, A.; Hierro, C.; Mita, A.; Cervantes, A.; Chan, N.; Awad, M.; Calvo, E.; Moreno, V.; Govindan, R.; et al. Multicenter Phase I Study of Erdafitinib (JNJ-42756493), Oral Pan-Fibroblast Growth Factor Receptor Inhibitor, in Patients with Advanced or Refractory Solid Tumors. Clin. Cancer Res. 2019, 25, 4888–4897. [Google Scholar] [CrossRef]
- Holzhauser, S.; Wild, N.; Zupancic, M.; Ursu, R.G.; Bersani, C.; Näsman, A.; Kostopoulou, O.N.; Dalianis, T. Targeted Therapy with PI3K and FGFR Inhibitors on Human Papillomavirus Positive and Negative Tonsillar and Base of Tongue Cancer Lines with and Without Corresponding Mutations. Front. Oncol. 2021, 11, 640490. [Google Scholar] [CrossRef]
- Pascual, J.; Lim, J.S.; MacPherson, I.R.J.; Armstrong, A.C.; Ring, A.; Okines, A.F.; Cutts, R.J.; Herrera-Abreu, M.T.; Gar-cia-Murillas, I.; Pearson, A.; et al. Triplet Therapy with Palbociclib, Taselisib, and Fulvestrant in PIK3CA-Mutant Breast Cancer and Doublet Palbociclib and Taselisib in Pathway-Mutant Solid Cancers. Cancer Discov. 2021, 11, 92–107. [Google Scholar] [CrossRef]
- Chen, X.; Yang, D.; Carey, J.; Karakas, C.; Albarracin, C.; Sahin, A.; Arun, B.; Durak, M.G.; Li, M.; Kohansal, M.; et al. Targeting Replicative Stress and DNA Repair by Combining PARP and Wee1 Kinase Inhibitors Is Synergistic in Triple Negative Breast Cancers with Cyclin E or BRCA1 Alteration. Cancers 2021, 13, 1656. [Google Scholar] [CrossRef]
- Seo, H.-R.; Nam, A.-R.; Bang, J.-H.; Oh, K.-S.; Kim, J.-M.; Yoon, J.; Kim, T.-Y.; Oh, D.-Y. Inhibition of WEE1 Potentiates Sensitivity to PARP Inhibitor in Biliary Tract Cancer. Cancer Res. Treat. 2022, 54, 541–553. [Google Scholar] [CrossRef]
- Pirotte, E.F.; Holzhauser, S.; Owens, D.; Quine, S.; Al-Hussaini, A.; Christian, A.D.; Giles, P.J.; Man, S.T.; Evans, M.; Powell, N.G. Sensitivity to inhibition of DNA repair by Olaparib in novel oropharyngeal cancer cell lines infected with Human Papillomavirus. PLoS ONE 2018, 13, e0207934. [Google Scholar] [CrossRef]
- Foucquier, J.; Guedj, M. Analysis of drug combinations: Current methodological landscape. Pharmacol. Res. Perspect. 2015, 3, e00149. [Google Scholar] [CrossRef]
- Chou, T.-C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, F.J.; Sundaresan, A.; Zhang, J.; Pedamallu, C.S.; Halle, M.K.; Srinivasasainagendra, V.; Zhang, J.; Muhammad, N.; Stanley, J.; Markovina, S.; et al. Genomic Characterization and Therapeutic Targeting of HPV Undetected Cervical Carcinomas. Cancers 2021, 13, 4551. [Google Scholar] [CrossRef]
- Agostinetto, E.; Debien, V.; Marta, G.N.; Lambertini, M.; Piccart-Gebhart, M.; de Azambuja, E. CDK4/6 and PI3K inhibitors: A new promise for patients with HER2-positive breast cancer. Eur. J. Clin. Investig. 2021, 51, e13535. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.H.; Leung, H.W.C.; Wang, S.-Y.; Huang, S.-S.; Chan, A.L.F. Efficacy and safety of CDK4/6 and PI3K/AKT/mTOR inhibitors as second-line treatment in postmenopausal patients with hormone receptor-positive, HER-2-negative metastatic breast cancer: A network meta-analysis. Expert Opin. Drug Saf. 2021, 20, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Freelander, A.; Brown, L.; Parker, A.; Segara, D.; Portman, N.; Lau, B.; Lim, E. Molecular Biomarkers for Contemporary Therapies in Hormone Receptor-Positive Breast Cancer. Genes 2021, 12, 285. [Google Scholar] [CrossRef] [PubMed]
- Holzhauser, S.; Kostopoulou, O.N.; Ohmayer, A.; Lange, B.K.; Andonova, T.; Bersani, C.; Dalianis, T. In vitro antitumor effects of FGFR and PI3K inhibitors on human papillomavirus positive and negative tonsillar and base of tongue cancer cell lines. Oncol. Lett. 2019, 18, 6249–6260. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Patel, A.A.; Wang, J.; Frederick, M.J.; Kalu, N.N.; Zhao, M.; Fitzgerald, A.L.; Xie, T.-X.; Silver, N.L.; Caulin, C.; et al. Wee-1 Kinase Inhibition Sensitizes High-Risk HPV+ HNSCC to Apoptosis Accompanied by Downregulation of MCl-1 and XIAP Antiapoptotic Proteins. Clin. Cancer Res. 2015, 21, 4831–4844. [Google Scholar] [CrossRef] [Green Version]
- Busch, C.-J.; Kröger, M.S.; Jensen, J.; Kriegs, M.; Gatzemeier, F.; Petersen, C.; Muenscher, A.; Rothkamm, K.; Rieckmann, T. G2-checkpoint targeting and radiosensitization of HPV/p16-positive HNSCC cells through the inhibition of Chk1 and Wee1. Radiother. Oncol. 2017, 122, 260–266. [Google Scholar] [CrossRef]
- Ha, D.-H.; Min, A.; Kim, S.; Jang, H.; Kim, S.H.; Kim, H.-J.; Ryu, H.S.; Ku, J.-L.; Lee, K.-H.; Im, S.-A. Antitumor effect of a WEE1 inhibitor and potentiation of olaparib sensitivity by DNA damage response modulation in triple-negative breast cancer. Sci. Rep. 2020, 10, 9930. [Google Scholar] [CrossRef]
- Zhou, C.; Parsons, J.L. The radiobiology of HPV-positive and HPV-negative head and neck squamous cell carcinoma. Expert Rev. Mol. Med. 2020, 22, e3. [Google Scholar] [CrossRef]
- Morgan, M.A.; Lawrence, T.S. Molecular Pathways: Overcoming Radiation Resistance by Targeting DNA Damage Response Pathways. Clin. Cancer Res. 2015, 21, 2898–2904. [Google Scholar] [CrossRef] [Green Version]
- Holzhauser, S.; Lukoseviciute, M.; Andonova, T.; Ursu, R.G.; Dalianis, T.; Wickström, M.; Kostopoulou, O.N. Targeting Fibroblast Growth Factor Receptor (FGFR) and Phosphoinositide 3-kinase (PI3K) Signaling Pathways in Medulloblastoma Cell Lines. Anticancer Res. 2019, 40, 53–66. [Google Scholar] [CrossRef]
- Topacio, B.R.; Zatulovskiy, E.; Cristea, S.; Xie, S.; Tambo, C.S.; Rubin, S.M.; Sage, J.; Kõivomägi, M.; Skotheim, J.M. Cyclin D-Cdk4,6 Drives Cell-Cycle Progression via the Retinoblastoma Protein’s C-Terminal Helix. Mol. Cell 2019, 74, 758–770.e4. [Google Scholar] [CrossRef]
- Roberts, P.J.; Bisi, J.E.; Strum, J.C.; Combest, A.J.; Darr, D.B.; Usary, J.E.; Zamboni, W.C.; Wong, K.K.; Perou, C.; Sharpless, N. Multiple Roles of Cyclin-Dependent Kinase 4/6 Inhibitors in Cancer Therapy. JNCI J. Natl. Cancer Inst. 2012, 104, 476–487. [Google Scholar] [CrossRef]
- Corona, S.P.; Generali, D. Abemaciclib: A CDK4/6 inhibitor for the treatment of HR+/HER2– advanced breast cancer. Drug Des. Dev. Ther. 2018, 12, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Roberts, P.J.; Kumarasamy, V.; Witkiewicz, A.K.; Knudsen, E.S. Chemotherapy and CDK4/6 Inhibitors: Unexpected Bedfellows. Mol. Cancer Ther. 2020, 19, 1575–1588. [Google Scholar] [CrossRef]
- Adkins, D.; Ley, J.; Neupane, P.; Worden, F.; Sacco, A.G.; Palka, K.; Grilley-Olson, J.E.; Maggiore, R.; Salama, N.; Trinkaus, K.; et al. Palbociclib and cetuximab in platinum-resistant and in cetuximab-resistant human papillomavirus-unrelated head and neck cancer: A multicentre, multigroup, phase 2 trial. Lancet Oncol. 2019, 20, 1295–1305. [Google Scholar] [CrossRef]
- Xiong, Y.; Li, T.; Assani, G.; Ling, H.; Zhou, Q.; Zeng, Y.; Zhou, F.; Zhou, Y. Ribociclib, a selective cyclin D kinase 4/6 inhibitor, inhibits proliferation and induces apoptosis of human cervical cancer in vitro and in vivo. Biomed. Pharmacother. 2019, 112, 108602. [Google Scholar] [CrossRef]
- Van Caloen, G.; Machiels, J.-P. Potential role of cyclin-dependent kinase 4/6 inhibitors in the treatment of squamous cell carcinoma of the head and neck. Curr. Opin. Oncol. 2019, 31, 122–130. [Google Scholar] [CrossRef]
- Holzhauser, S.; Pirotte, E.; Jones, J.; Owens, D.; Al-Hussaini, A.; Giles, P.; Evans, M.; Man, S.; Powell, N. Sensitivity of human papillomavirus-positive and -negative oropharyngeal cancer cell lines to ionizing irradiation. Oncol. Rep. 2020, 44, 1717–1726. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µM) b | ||||
|---|---|---|---|---|
| Drugs | Cell Lines | 24 h | 48 h | 72 h |
| CDK4/6 (PD-0332991) | CU-OP-2 | 25.1 | 26.8 | 35.9 |
| CU-OP-3 | 13.4 | 15.2 | 12.3 | |
| CU-OP-20 | 10.4 | 9.8 | 10.1 | |
| CU-OP-17 | 21.0 | 16.6 | 14.4 | |
| PARP (BMN-673) | CU-OP-2 | 291.4 a | 127.2 a | 14.9 |
| CU-OP-3 | 3219 a | 81.4 a | 13.9 | |
| CU-OP-20 | 229.5 a | 30.9 | 6.0 | |
| CU-OP-17 | 93.8 a | 12.8 | 4.8 | |
| WEE1 (MK-1775) | CU-OP-2 | 3.4 | 2.4 | 1.7 |
| CU-OP-3 | 9.6 | 0.6 | 0.2 | |
| CU-OP-20 | 2.9 | 0.4 | 0.2 | |
| CU-OP-17 | 6.5 | 0.4 | 0.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostopoulou, O.N.; Zupancic, M.; Pont, M.; Papin, E.; Lukoseviciute, M.; Mikelarena, B.A.; Holzhauser, S.; Dalianis, T. Targeted Therapy of HPV Positive and Negative Tonsillar Squamous Cell Carcinoma Cell Lines Reveals Synergy between CDK4/6, PI3K and Sometimes FGFR Inhibitors, but Rarely between PARP and WEE1 Inhibitors. Viruses 2022, 14, 1372. https://doi.org/10.3390/v14071372
Kostopoulou ON, Zupancic M, Pont M, Papin E, Lukoseviciute M, Mikelarena BA, Holzhauser S, Dalianis T. Targeted Therapy of HPV Positive and Negative Tonsillar Squamous Cell Carcinoma Cell Lines Reveals Synergy between CDK4/6, PI3K and Sometimes FGFR Inhibitors, but Rarely between PARP and WEE1 Inhibitors. Viruses. 2022; 14(7):1372. https://doi.org/10.3390/v14071372
Chicago/Turabian StyleKostopoulou, Ourania N., Mark Zupancic, Mariona Pont, Emma Papin, Monika Lukoseviciute, Borja Agirre Mikelarena, Stefan Holzhauser, and Tina Dalianis. 2022. "Targeted Therapy of HPV Positive and Negative Tonsillar Squamous Cell Carcinoma Cell Lines Reveals Synergy between CDK4/6, PI3K and Sometimes FGFR Inhibitors, but Rarely between PARP and WEE1 Inhibitors" Viruses 14, no. 7: 1372. https://doi.org/10.3390/v14071372
APA StyleKostopoulou, O. N., Zupancic, M., Pont, M., Papin, E., Lukoseviciute, M., Mikelarena, B. A., Holzhauser, S., & Dalianis, T. (2022). Targeted Therapy of HPV Positive and Negative Tonsillar Squamous Cell Carcinoma Cell Lines Reveals Synergy between CDK4/6, PI3K and Sometimes FGFR Inhibitors, but Rarely between PARP and WEE1 Inhibitors. Viruses, 14(7), 1372. https://doi.org/10.3390/v14071372