
Evaluation of New Polyclonal Antibody Developed for Serological Diagnostics of Tomato Mosaic Virus

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. ToMV SL-1 Isolate and Tomato Genotypes
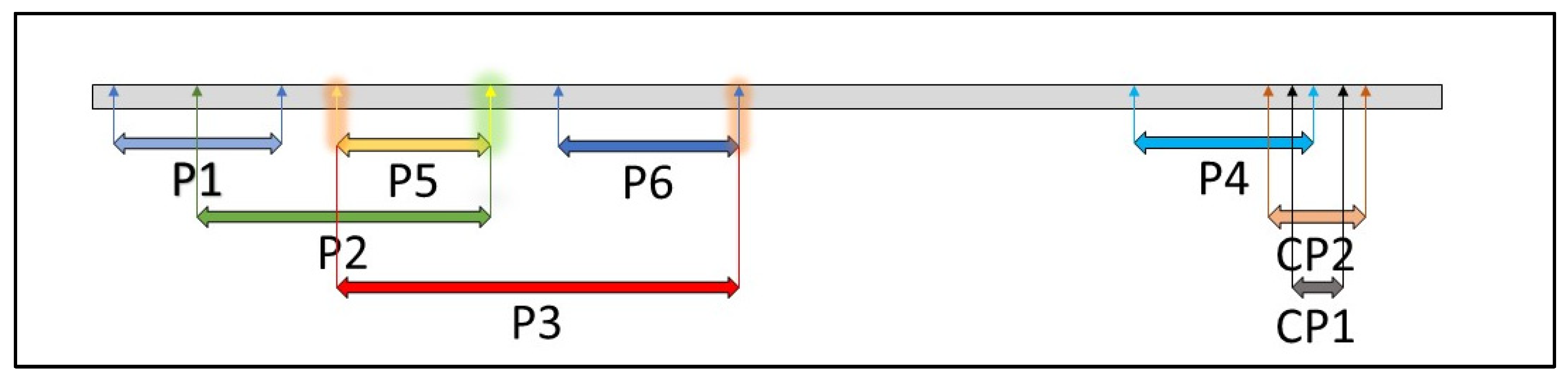
2.2. ToMV SL-1 Isolate Characterization
2.3. Development ToMV-Specific Polyclonal Antibody
2.4. Serological Testing
2.5. Statistical Evaluation
3. Results
3.1. Identity of ToMV SL-1 Isolate
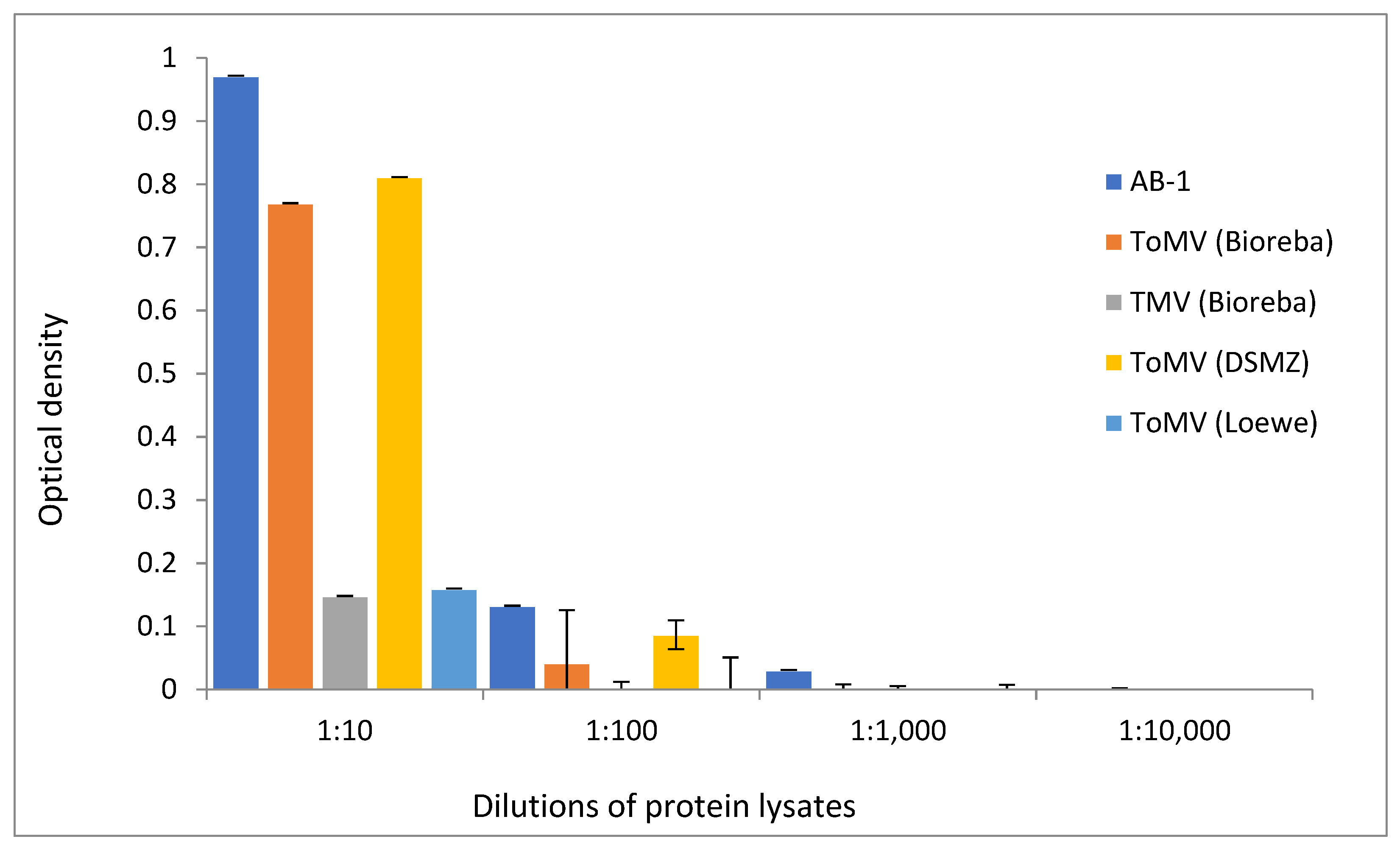
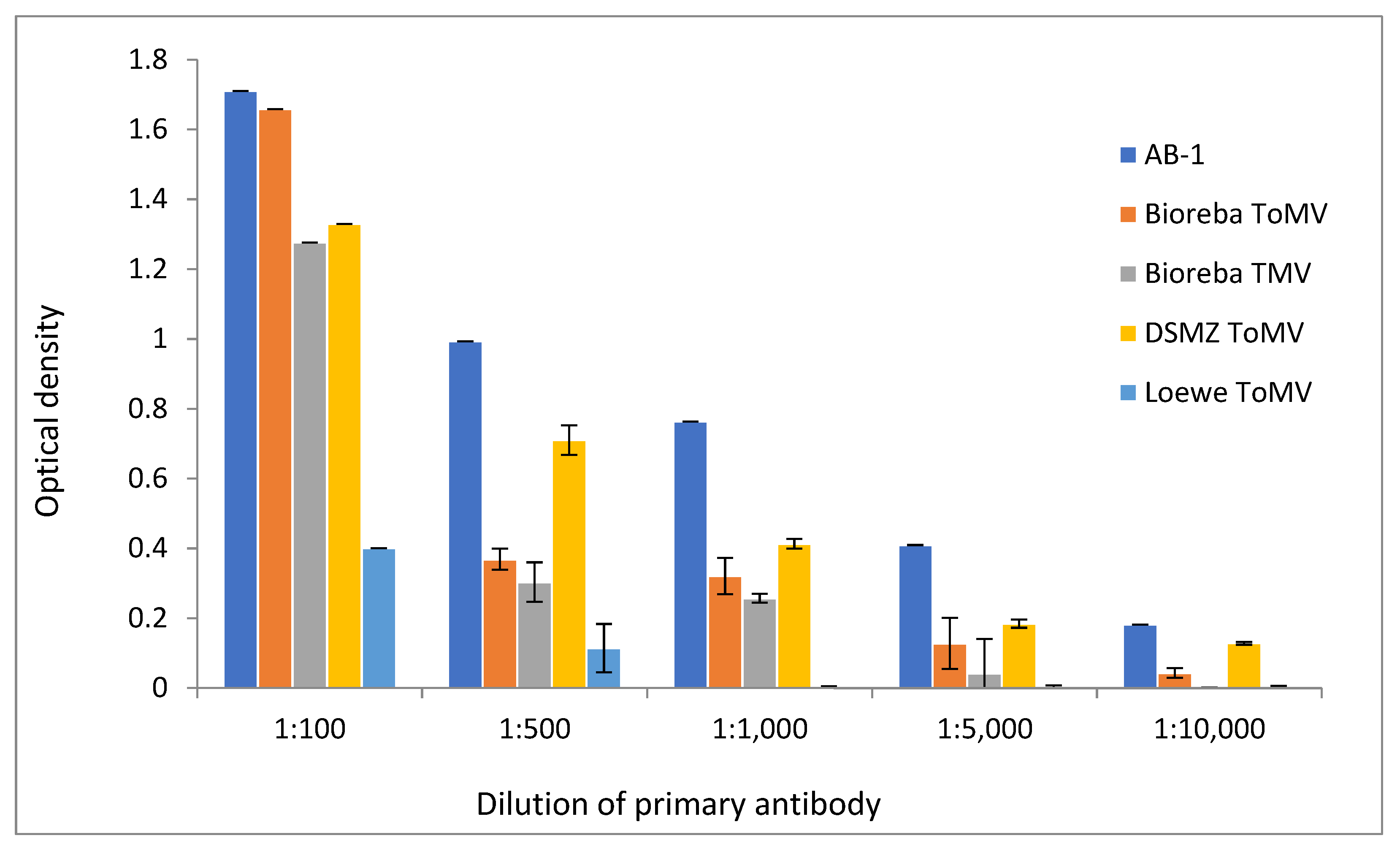
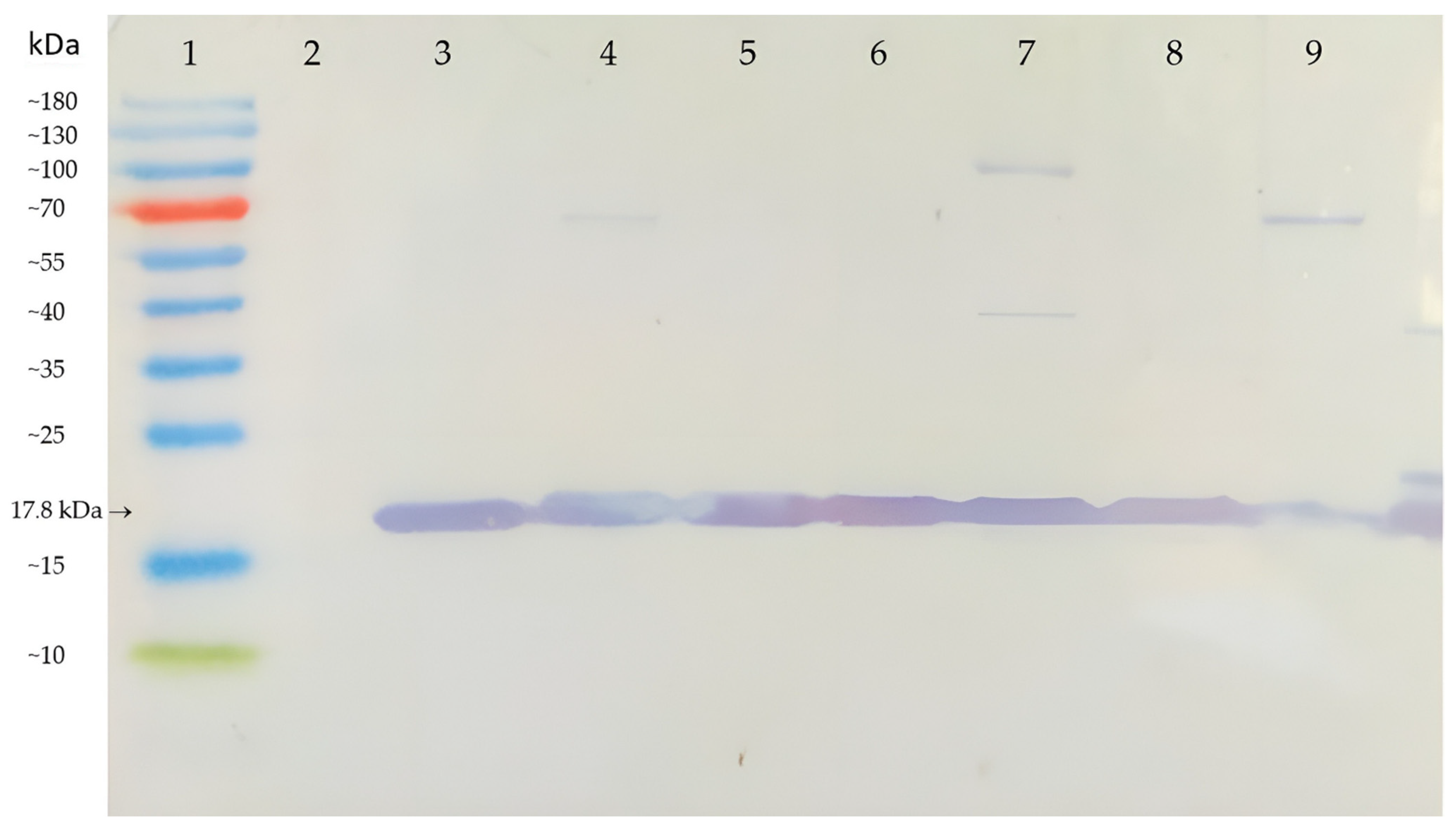
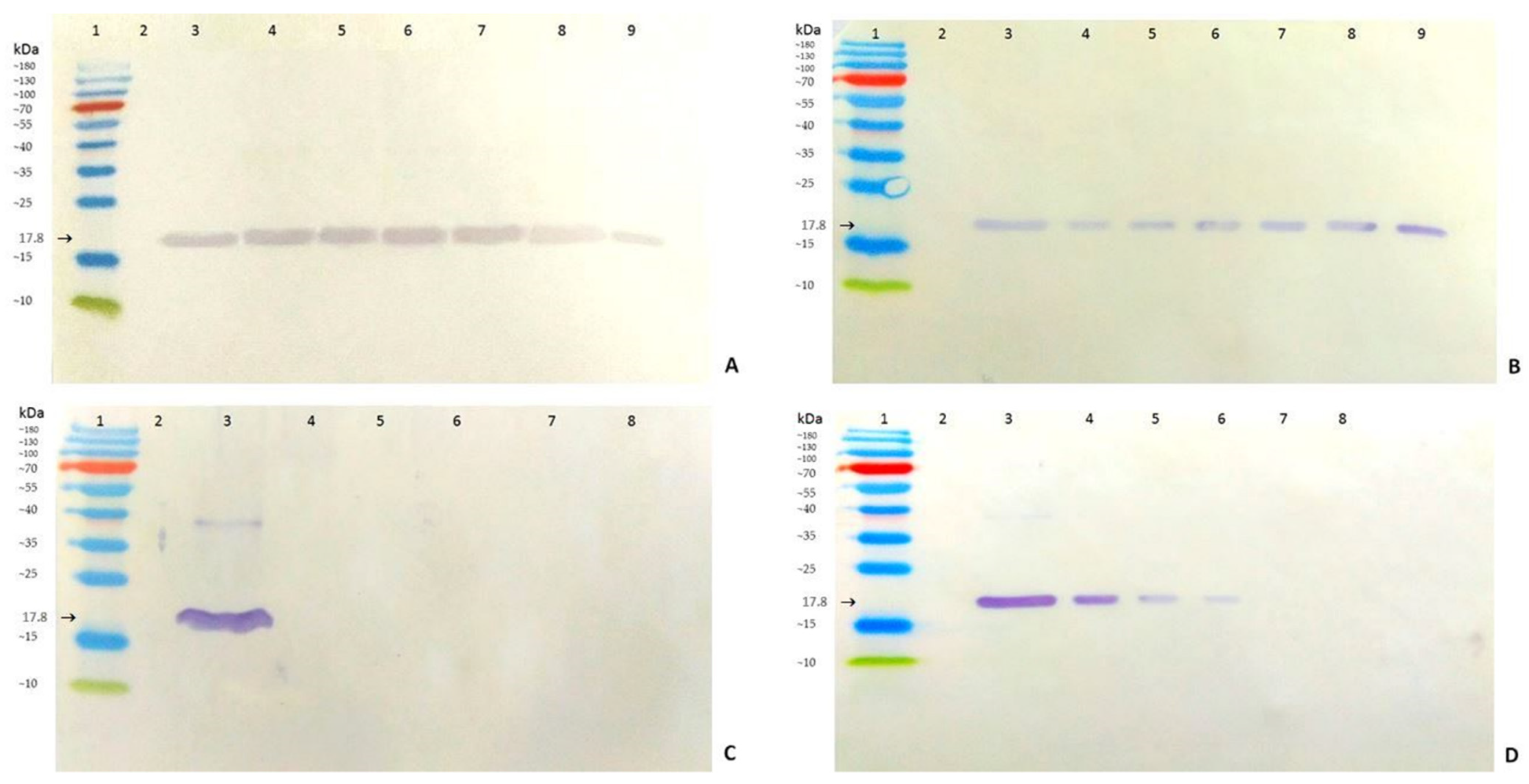
3.2. Sensitivity of AB-1 Antibody
3.3. Specificity and Cross-Reactivity of AB-1 Antibody
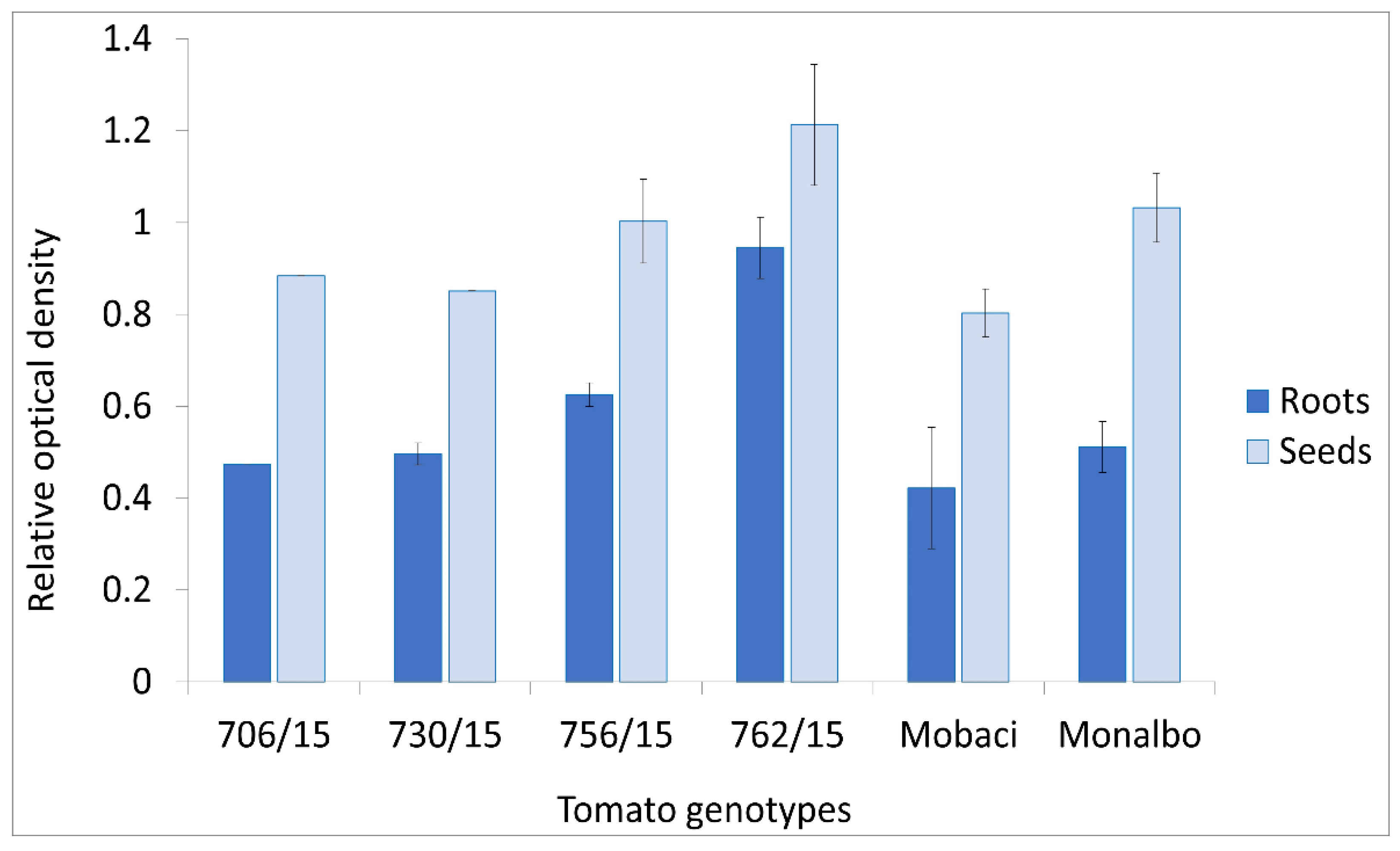
3.4. AB-1 Antibody in ToMV Analysis in Roots and Seeds
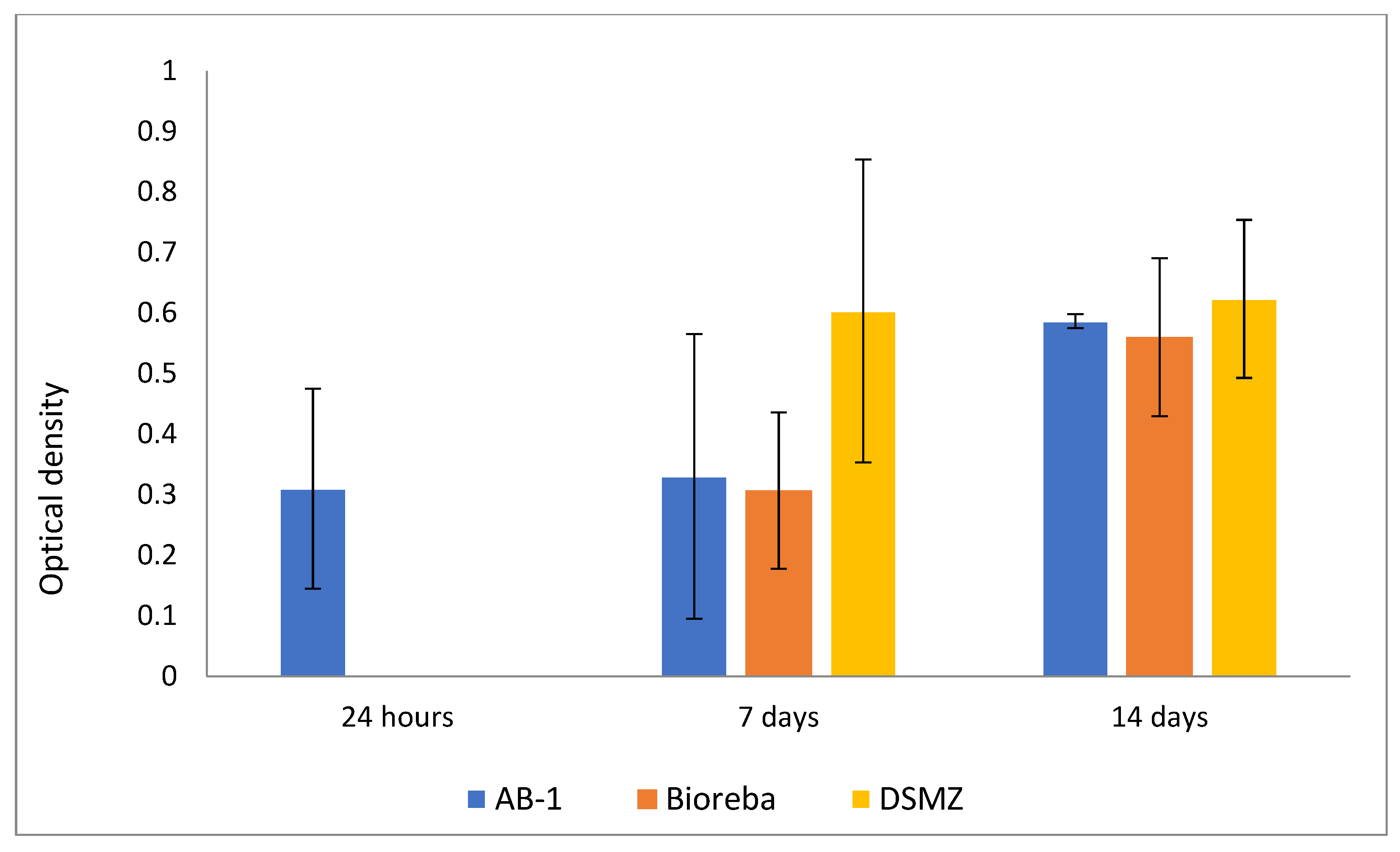
3.5. AB-1 Antibody in Analysis of ToMV Persistence and Resilience
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blancard, D. Principal Characteristics of Pathogenic Agents and Methods of Control. In Tomato Diseases, 2nd ed.; Blancard, D., Ed.; Manson Publishing Ltd.: London, UK, 2012; pp. 413–650. [Google Scholar] [CrossRef]
- Jones, J.B.; Zitter, T.A.; Momol, T.M.; Miller, S.A. Compendium of Tomato Disease and Pests, 2nd ed.; The American Phytopathological Society: St. Paul, MN, USA, 2016; p. 176. [Google Scholar]
- Tolman, J.H.; McLeod, D.G.R.; Harris, C.R. Cost of crop losses in processing tomato and cabbage in southwestern Ontario due to insects, weeds and/or diseases. Can. J. Plant Sci. 2004, 84, 915–921. [Google Scholar] [CrossRef]
- Panno, S.; Davino, S.; Caruso, A.G.; Bertacca, S.; Crnogorac, A.; Mandić, A.; Noris, E.; Matić, S. A review of the most common and economically important diseases that undermine the cultivation of tomato crop in the Mediterranean basin. Agronomy 2021, 11, 2188. [Google Scholar] [CrossRef]
- Hanssen, I.M.; Lapidot, M.; Thomma, B.P. Emerging viral diseases of tomato crops. Mol. Plant Microbe Interact. 2010, 23, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Hančinský, R.; Mihálik, D.; Mrkvová, M.; Candresse, T.; Glasa, M. Plant viruses infecting Solanaceae family members in the cultivated and wild environments: A review. Plants 2020, 9, 667. [Google Scholar] [CrossRef]
- Ong, S.N.; Taheri, S.; Othman, R.Y.; Teo, C.H. Viral disease of tomato crops (Solanum lycopesicum L.): An overview. J. Plant Dis. Prot. 2020, 127, 725–739. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2021 Release. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 8 April 2022).
- Almeida, J.E.M.; Figueira, A.D.R.; Duarte, P.D.S.G.; Lucas, M.A.; Alencar, N.E. Procedure for detecting tobamovirus in tomato and pepper seeds decreases the cost analysis. Bragantia 2018, 77, 590–598. [Google Scholar] [CrossRef]
- Lanter, J.M.; McGuire, J.M.; Goode, M.J. Persistence of tomato mosaic virus in tomato debris and soil under field conditions. Plant Dis. 1982, 66, 552–555. [Google Scholar] [CrossRef]
- Boben, J.; Kramberger, P.; Petrovič, N.; Cankar, K.; Peterka, M.; Štrancar, A.; Ravinkar, M. Detection and quantification of Tomato mosaic virus in irrigation waters. Eur. J. Plant Pathol. 2007, 118, 59–71. [Google Scholar] [CrossRef]
- Pares, R.D.; Gunn, L.V.; Cresswell, G.C. Tomato mosaic virus infection in a recirculating nutrient solution. J. Phytopathol. 1992, 135, 192–198. [Google Scholar] [CrossRef]
- Meshi, T.; Watanabe, Y.; Okada, Y. Molecular pathology of tobacco mosaic virus revealed by biologically active cDNAs. In Genetic Engineering with Plant Viruses; Wilson, T.M.A., Davies, J.W., Eds.; CRC Press: Boca Raton, FL, USA, 1992; pp. 149–186. [Google Scholar]
- Li, Y.; Wu, M.Y.; Song, H.H.; Hu, X.; Qiu, B.S. Identification of a tobacco protein interacting with tomato mosaic virus coat protein and facilitating long-distance movement of virus. Arch. Virol. 2005, 150, 1993–2008. [Google Scholar] [CrossRef]
- Pelham, J. Resistance in tomato to tomato mosaic virus. Euphytica 1966, 15, 258–267. [Google Scholar] [CrossRef]
- Pelham, J. Strain-genotype interaction of tobacco mosaic virus in tomato. Ann. Appl. Biol. 1972, 71, 219–288. [Google Scholar] [CrossRef]
- Weber, H.; Schultze, S.; Pfitzner, A.J. Two amino acid substitutions in the tomato mosaic virus 30-kilodalton movement protein confer the ability to overcome the Tm-2(2) resistance gene in the tomato. J. Virol. 1993, 67, 6432–6438. [Google Scholar] [CrossRef] [PubMed]
- Strasser, M.; Pfitzner, A.J. The double-resistance-breaking Tomato mosaic virus strain ToMV1-2 contains two independent single resistance-breaking domains. Arch. Virol. 2007, 152, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of plant viruses and disease management: Relevance of genetic diversity and evolution. Front. Plant Sci. 2020, 11, 1092. [Google Scholar] [CrossRef] [PubMed]
- Mehetre, G.T.; Leo, V.V.; Singh, G.; Sorokan, A.; Maksimov, I.; Yadav, M.K.; Upadhyaya, K.; Hashem, A.; Alsaleh, A.N.; Dawoud, T.M.; et al. Current developments and challenges in plant viral diagnostics: A systematic review. Viruses 2021, 13, 412. [Google Scholar] [CrossRef]
- Alon, D.M.; Hak, H.; Bornstein, M.; Pines, G.; Spiegelman, Z. Differential detection of the Tobamoviruses Tomato mosaic virus (ToMV) and Tomato brown rugose fruit virus (ToBRFV) using CRISPR-Cas12a. Plants 2021, 10, 1256. [Google Scholar] [CrossRef]
- Villamor, D.E.V.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E. High throughput sequencing for plant virus detection and discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Liefting, L.W.; Waite, D.W.; Thompson, J.R. Application of Oxford Nanopore technology to plant virus detection. Viruses 2021, 13, 1424. [Google Scholar] [CrossRef]
- Boonham, N.; Kreuze, J.; Winter, S.; van der Vlugt, R.; Bergervoet, J.; Tomlinson, J.; Mumford, R. Methods in virus diagnostics: From ELISA to next generation sequencing. Virus Res. 2014, 186, 20–31. [Google Scholar] [CrossRef]
- Sheveleva, A.; Ivanov, P.; Gasanova, T.; Osipov, G.; Chirkov, S. Sequence analysis of Plum pox virus strain C isolates from Russia revealed prevalence of the D96E mutation in the universal epitope and interstrain recombination events. Viruses 2018, 10, 450. [Google Scholar] [CrossRef] [PubMed]
- Souiri, A.; Zemzami, M.; Amzazi, S.; Ennaji, M.M. Polyclonal and monoclonal antibody-based methods for detection of plant viruses. Eur. J. Sci. Res. 2014, 123, 281–295. [Google Scholar]
- Sui, X.; Li, R.; Padmanabhan, C.; Ling, K.-S. Molecular, serological, and biological characterization of the emerging Tomato mottle mosaic virus on tomato. Acta Hortic. 2018, 1207, 281–286. [Google Scholar] [CrossRef]
- International Seed Federation. Detection of Infectious Tobamoviruses in Tomato Seed; v. 6.2; International Seed federation: Nyon, Switzerland, 2019; pp. 1–8. [Google Scholar]
- Sihelská, N.; Vozárová, Z.; Predajňa, L.; Šoltys, K.; Hudcovicová, M.; Mihálik, D.; Kraic, J.; Mrkvová, M.; Kúdela, O.; Glasa, M. Experimental infection of different tomato genotypes with Tomato mosaic virus led to a low viral population heterogeneity in the capsid protein encoding region. Plant Pathol. J. 2017, 33, 508–513. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gibbs, A.J.; Wood, J.; Garcia-Arenal, F.; Ohshima, K.; Armstrong, J.S. Tobamoviruses have probably co-diverged with their eudicotyledonous hosts for at least 110 million years. Virus Evol. 2015, 1, vev019. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucl. Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef]
- Parker, J.M.; Guo, D.; Hodges, R.S. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: Correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry 1986, 25, 5425–5432. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar] [CrossRef]
- International Union for the Protection of New Varieties of Plants. Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability. Available online: https://www.upov.int/edocs/mdocs/upov/en/twv/44/tg_44_11_proj_3.pdf (accessed on 13 April 2022).
- Koh, S.H.; Li, H.; Sivasithamparam, K.; Admiraal, R.; Jones, M.G.K.; Wylie, S.J. Low root-to-root transmission of a tobamovirus, yellow tailflower mild mottle virus, and resilience of its virions. Plant Pathol. 2018, 67, 651–659. [Google Scholar] [CrossRef]
- Garcia-Calvo, E.; García-García, A.; Madrid, R.; Martin, R.; García, T. From polyclonal sera to recombinant antibodies: A review of immunological detection of gluten in foodstuff. Foods 2021, 10, 66. [Google Scholar] [CrossRef] [PubMed]
- Duarte, K.M.R.; Gomes, L.H.; Gesztesi, J.-L.; Lopes, J.D.; Tavares, F.C.A. Monoclonal antibodies to identify tomato mosaic virus (ToMV). Brasil. J. Microbiol. 2001, 32, 240–242. [Google Scholar] [CrossRef]
- Dekker, E.L.; Dore, I.; Porta, C.; Regenmortel, M.H.W. Specificity of monoclonal antibodies used in the diagnosis of Tomato mosaic virus. Arch. Virol. 1987, 94, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, C.A.; Aggeler, B. Overlooked benefits of using polyclonal antibodies. BioTechniques 2018, 63, 127–136. [Google Scholar] [CrossRef]
- Hadas, R.; Pearlsman, M.; Gefen, T.; Lachman, O.; Hadar, E.; Sharabany, G.; Antignus, Y. Indexing system for Tomato mosaic virus (ToMV) in commercial tomato seed lots. Phytoparasitica 2004, 32, 421–424. [Google Scholar] [CrossRef]
- Lima, J.A.A.; Lima, R.C.A.; Goncalves, M.F.B. Production of polyclonal antisera specific to plant viruses by rabbit oral immunization. Fitopatol. Bras. 2001, 26, 774–777. [Google Scholar] [CrossRef]
- Hjelm, B.; Forsström, B.; Löfblom, J.; Rockberg, J.; Uhlén, M. Parallel immunizations of rabbits using the same antigen yield antibodies with similar, but not identical, epitopes. PLoS ONE 2012, 7, e45817. [Google Scholar] [CrossRef]
- Pumrachat, S.; Hongprayoon, R. Epitope-based peptide antigen of pepper yellow leaf curl Thailand virus for specific antibody production. Int. J. Agric. Technol. 2021, 17, 661–672. [Google Scholar]
- Soliman, A.M.; Alhudaib, K.A.; Rezk, A.A.; El-Beltagi, H.S.; El-Ganainy, S.M. Production of polyclonal antibodies to some potato viruses using synthetic peptides. Fresenius Environ. Bull. 2019, 28, 4870–4877. [Google Scholar]
- Darsono, N.; Azizah, N.N.; Putranty, K.M.; Astuti, N.T.; Addy, H.S.; Darmanto, W.; Sugiharto, B. Production of a polyclonal antibody against the recombinant coat protein of the Sugarcane mosaic virus and its application in the immunodiagnostic of sugarcane. Agronomy 2018, 8, 93. [Google Scholar] [CrossRef]
- Phatsaman, T.; Hongprayoon, R.; Wasee, S. Monoclonal antibody-based diagnostic assays for pepper mild mottle virus. J. Plant Pathol. 2020, 102, 327–333. [Google Scholar] [CrossRef]
- Abdelmoamen, A.H.; El-Dougdoug, N.K.; El-Borollosy, A.M.; El-Dougdoug, K.A. Serological and molecular characteristics of tomato mosaic tobamovirus coat protein. Arab. Univ. J. Agric. Sci. 2018, 26, 1613–1626. [Google Scholar] [CrossRef][Green Version]
- Vermulapati, B.; Druffel, K.L.; Husebye, D.; Eigenbrode, S.D.; Pappu, H.R. Development and application of ELISA assays for the detection of two members of the family Luteoviridae infecting legumes: Pea enation mosaic virus (genus Enamovirus) and Bean leafroll virus (genus Luteovirus). Ann. Appl. Biol. 2014, 165, 130–136. [Google Scholar] [CrossRef]
- Vincent, S.J.; Coutts, B.A.; Jones, R.A.C. Effects of introduced and indigenous viruses on native plants: Exploring their disease causing potential at the agro-ecological interface. PLoS ONE 2014, 9, e91224. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer (Amplicon Length) | Forward Primer | Reverse Primer |
|---|---|---|
| CP1 (128 bp) | 5′-CAGATTTCCTGGCGATGTTT-3′ | 5′-TCGGACTCTGCTGGTTTTCT-3′ |
| CP2 (439 bp) | 5′-TGGTCATCTGTATGGGCTGA-3′ | 5′-AAGATGCAGGTGCAGAGGTC-3′ |
| P1 (867 bp) | 5′-CACAAACAGCCACATCGTCC-3′ | 5′-ACCTCTCTATTAGAGGCTGG-3′ |
| P2 (1361 bp) | 5′-CGAGAGGGGCAACAAACATG-3′ | 5′-GTAAGACCACTCTCGTTGCT-3′ |
| P3 (1715 bp) | 5′-AGGTCTGAGTGGGATGTCGA-3′ | 5′-ACAGGATTGATAGACGCAGC-3′ |
| P4 (742 bp) | 5′-CTTGTTGTGTCCGGTGAGTG-3′ | 5′-CCACACCTCGCTGAACTGCT-3′ |
| P5 (505 bp) | 5′-AGGTCTGAGTGGGATGTCGA-3′ | 5′-GTAAGACCACTCTCGTTGCT-3′ |
| P6 (707 bp) | 5′-ACGTCGCATTACTGGAGCAC-3′ | 5′-CACAGGATTGATAGACGCAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mrkvová, M.; Hančinský, R.; Grešíková, S.; Kaňuková, Š.; Barilla, J.; Glasa, M.; Hauptvogel, P.; Kraic, J.; Mihálik, D. Evaluation of New Polyclonal Antibody Developed for Serological Diagnostics of Tomato Mosaic Virus. Viruses 2022, 14, 1331. https://doi.org/10.3390/v14061331
Mrkvová M, Hančinský R, Grešíková S, Kaňuková Š, Barilla J, Glasa M, Hauptvogel P, Kraic J, Mihálik D. Evaluation of New Polyclonal Antibody Developed for Serological Diagnostics of Tomato Mosaic Virus. Viruses. 2022; 14(6):1331. https://doi.org/10.3390/v14061331
Chicago/Turabian StyleMrkvová, Michaela, Richard Hančinský, Simona Grešíková, Šarlota Kaňuková, Ján Barilla, Miroslav Glasa, Pavol Hauptvogel, Ján Kraic, and Daniel Mihálik. 2022. "Evaluation of New Polyclonal Antibody Developed for Serological Diagnostics of Tomato Mosaic Virus" Viruses 14, no. 6: 1331. https://doi.org/10.3390/v14061331
APA StyleMrkvová, M., Hančinský, R., Grešíková, S., Kaňuková, Š., Barilla, J., Glasa, M., Hauptvogel, P., Kraic, J., & Mihálik, D. (2022). Evaluation of New Polyclonal Antibody Developed for Serological Diagnostics of Tomato Mosaic Virus. Viruses, 14(6), 1331. https://doi.org/10.3390/v14061331