
Mongolia Gerbils Are Broadly Susceptible to Hepatitis E Virus

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. HEV Strains
2.2. Inoculation of Mongolia Gerbils and the Sample Collection
2.3. Extraction and Detection of HEV RNA
2.4. Detection of Anti-HEV IgG Antibodies
2.5. Liver Enzyme Level
2.6. Viral Genome Sequencing
3. Results
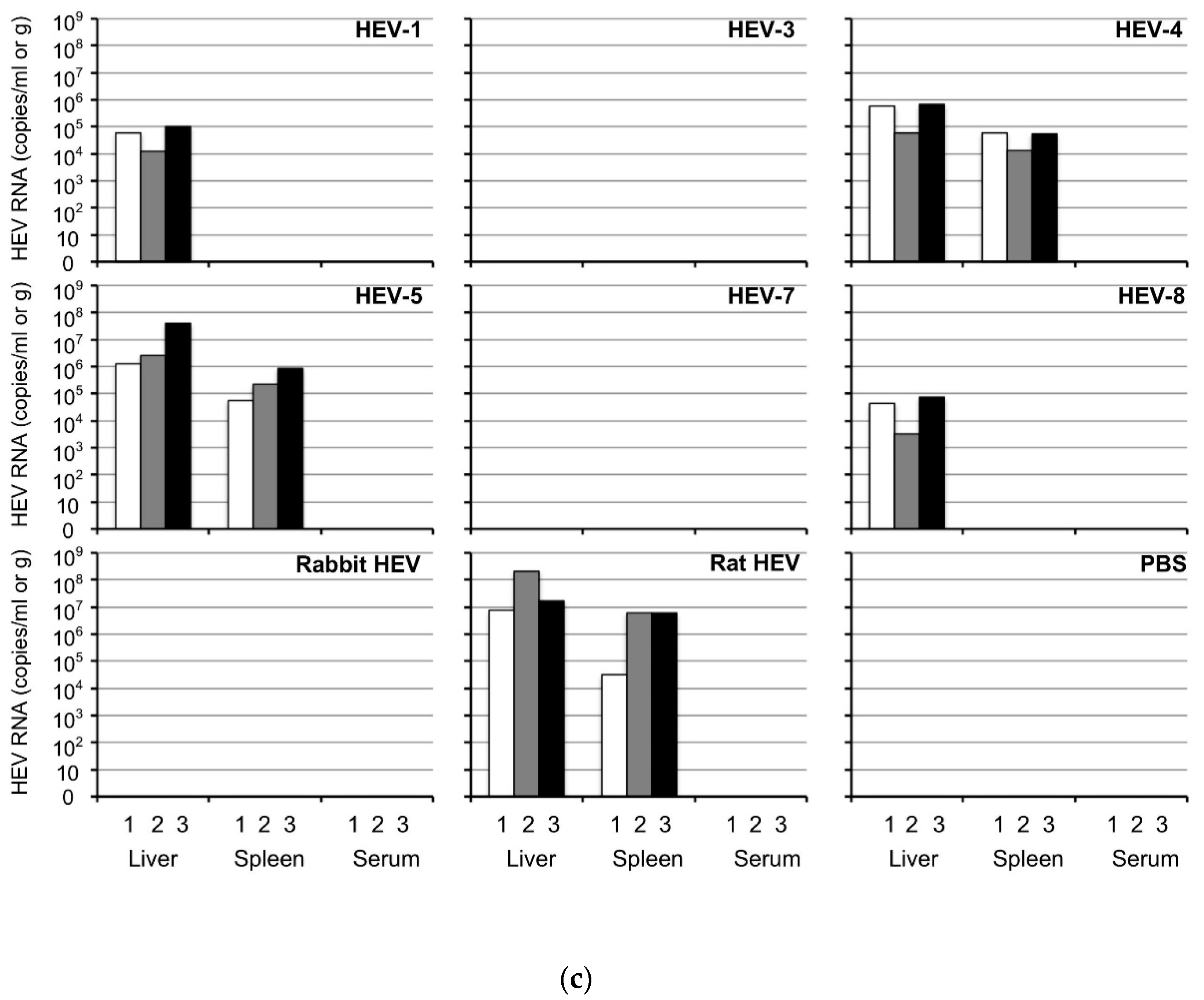
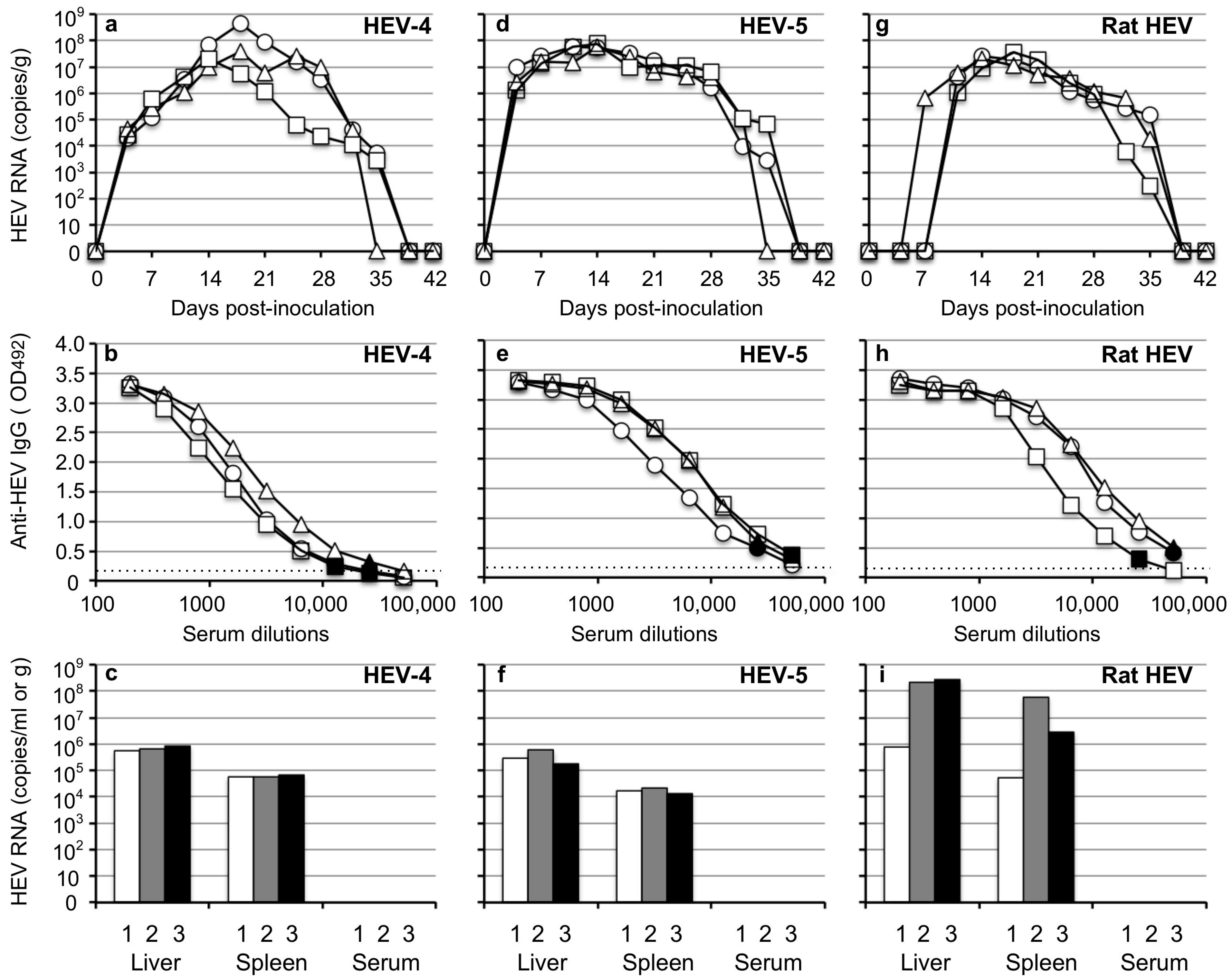
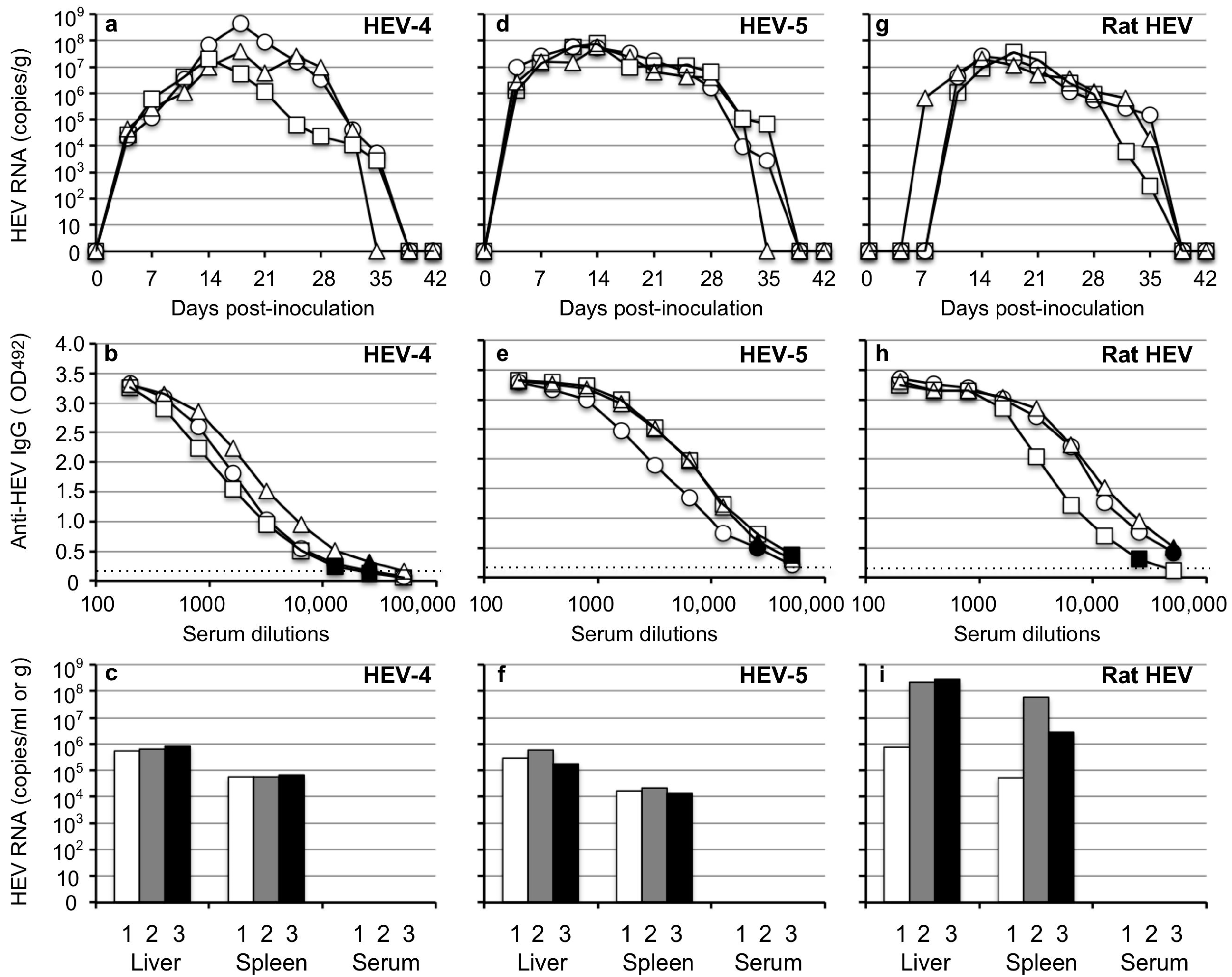
3.1. Mongolia Gerbils Had Different Susceptibilities to HEV
3.2. Analyses of the Entire Genomes of HEV-4, HEV-5, and Rat HEV
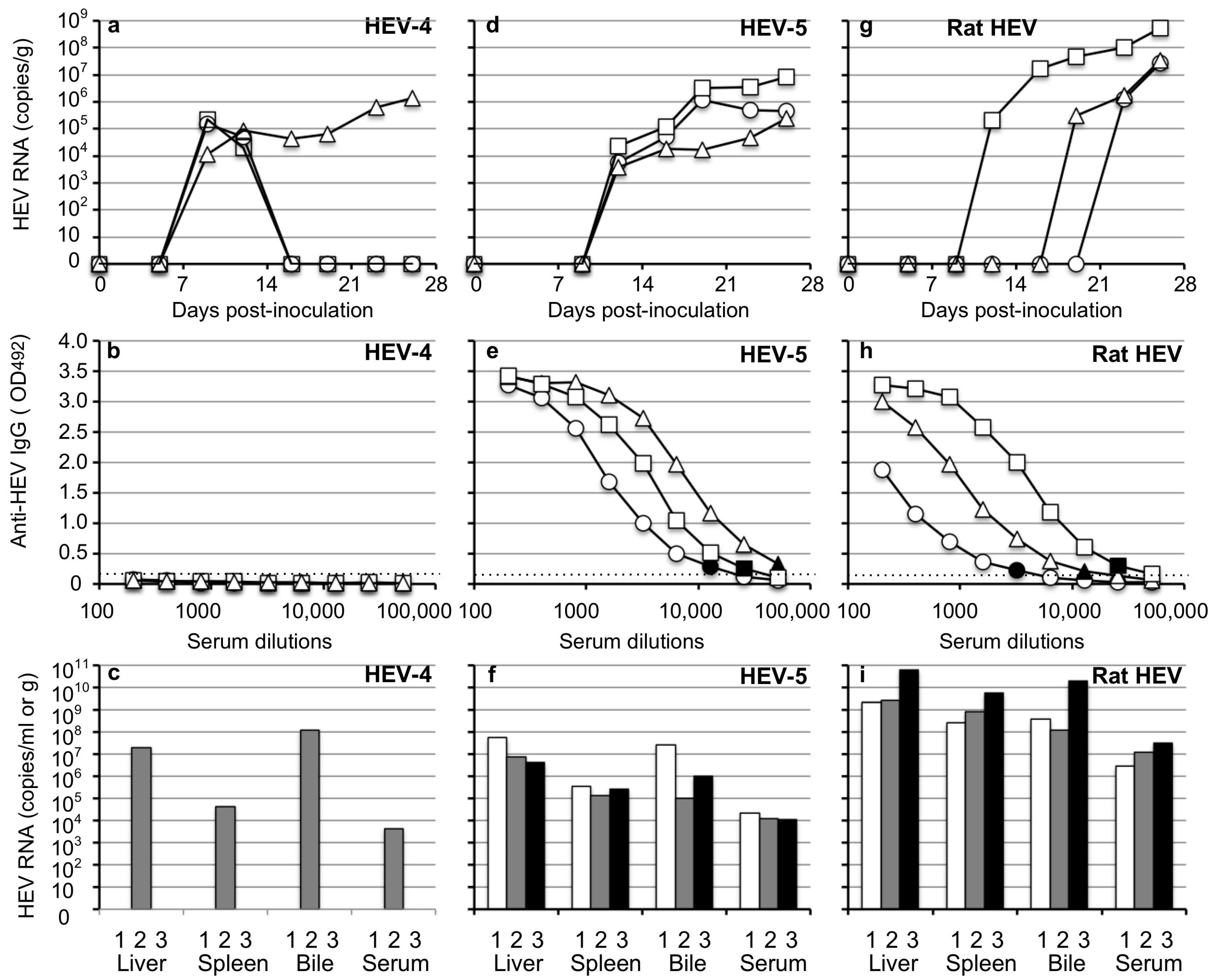
3.3. Infectivity of HEV Excreted in the Fecal Specimens
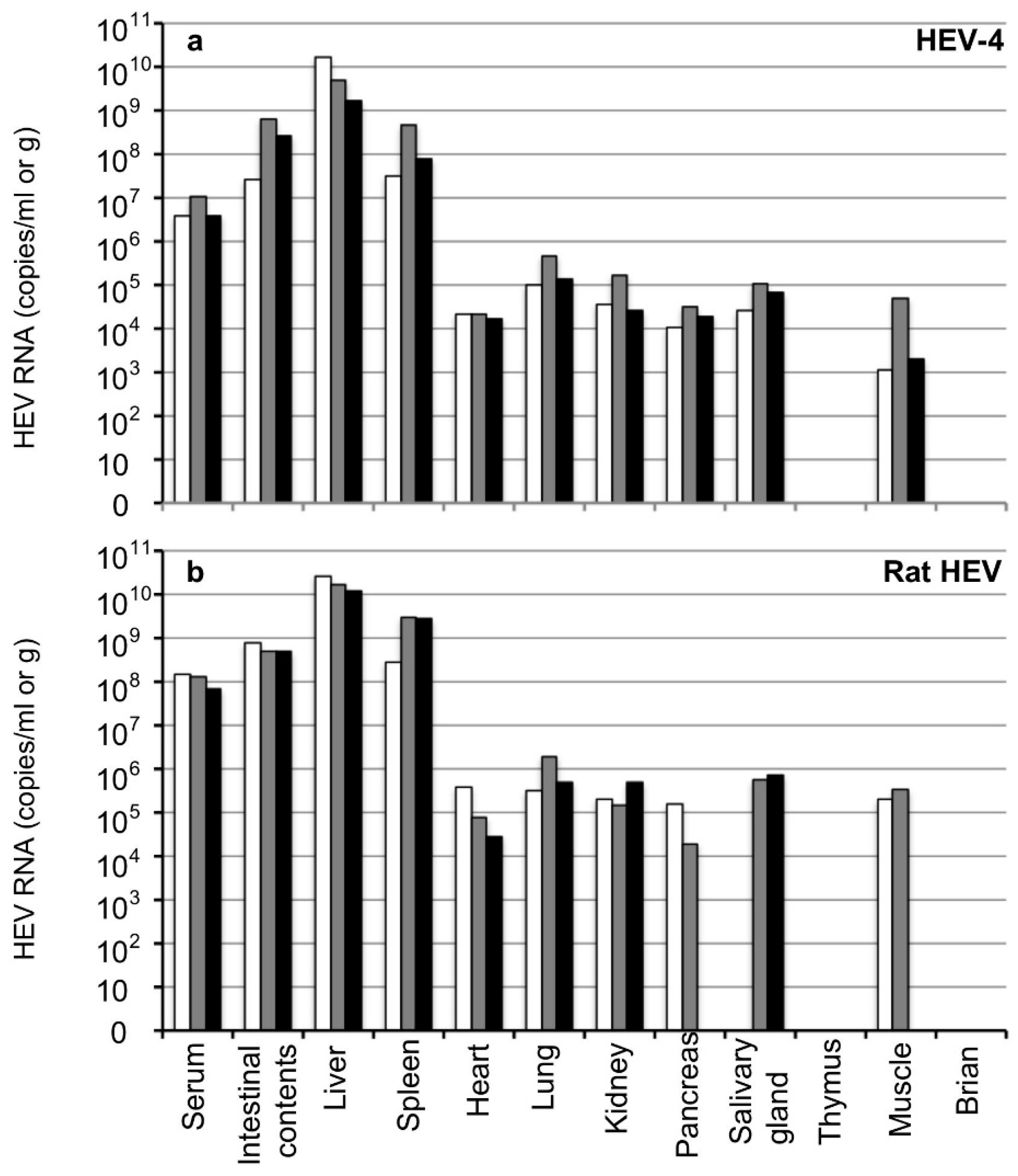
3.4. Detection of HEV in Tissues at the Early Stage of Infection
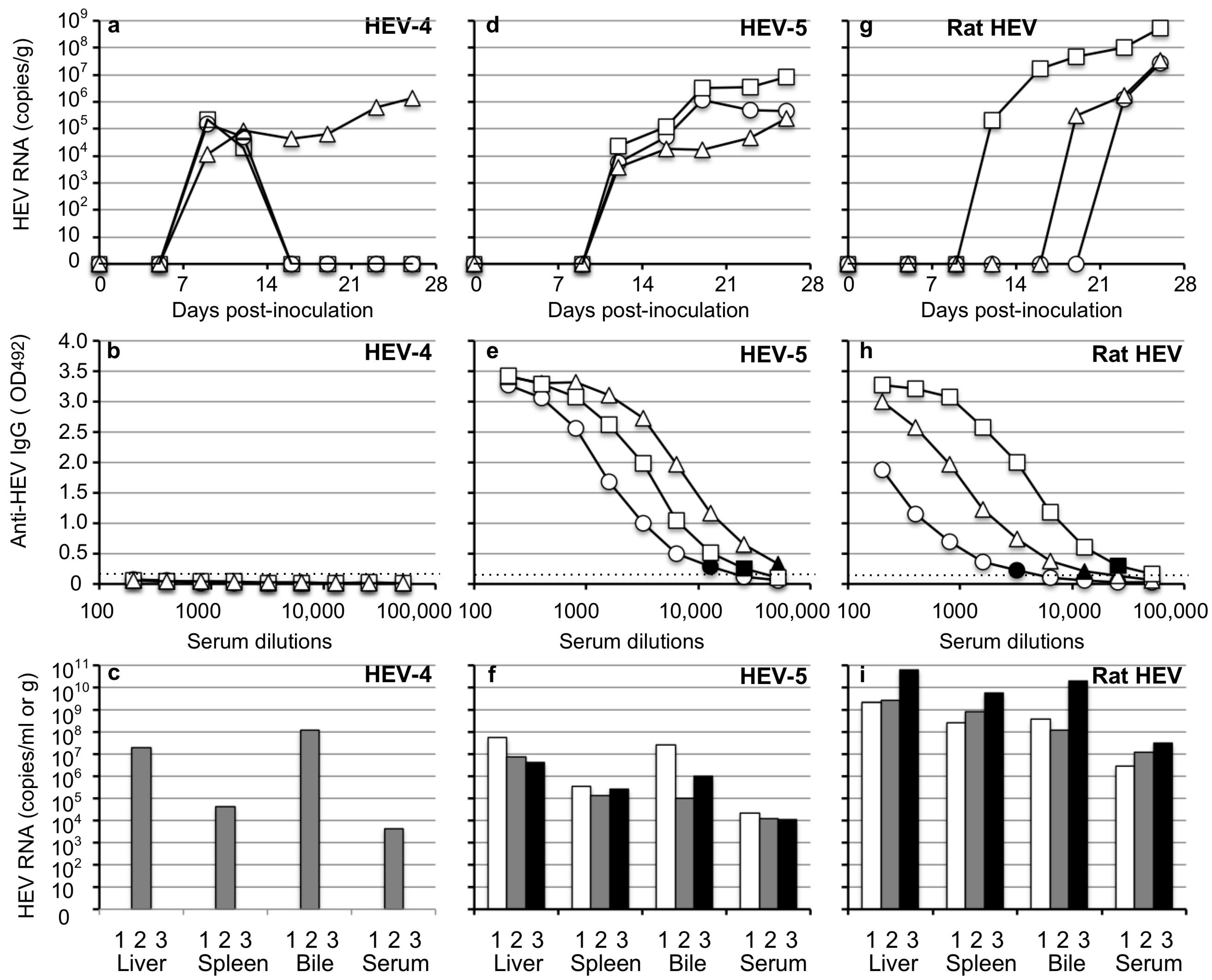
3.5. Transmission of HEV-4, HEV-5, and Rat HEV by an Oral Inoculation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meng, X.J.; Anderson, D.A.; Arankalle, V.A.; Emerson, S.U.; Harrison, T.J.; Jameel, S.; Okamoto, H. Hepeviridae. In Virus Taxonomy: Ninth Report of the ICTV; King, A.M.A., Michael, J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier/Academic Press: London, UK, 2012; pp. 1021–1028. [Google Scholar]
- Kamar, N.; Selves, J.; Mansuy, J.-M.; Ouezzani, L.; Péron, J.-M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E Virus and Chronic Hepatitis in Organ-Transplant Recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.B.; Simmonds, P.; International Committee on Taxonomy of Viruses Hepeviridae Study Group; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95 Pt 10, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Simmonds, P.; Izopet, J.; Oliveira-Filho, E.F.; Ulrich, R.G.; Johne, R.; Koenig, M.; Jameel, S.; Harrison, T.J.; Meng, X.-J.; et al. Proposed reference sequences for hepatitis E virus subtypes. J. Gen. Virol. 2016, 97, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Suzuki, J.; Matsuda, A.; Ishida, T.; Ami, Y.; Suzaki, Y.; Adachi, I.; Wakita, T.; Takeda, N.; Li, T.-C. Hepatitis E Virus Outbreak in Monkey Facility, Japan. Emerg. Infect. Dis. 2012, 18, 2032–2034. [Google Scholar] [CrossRef]
- Meng, X.J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Takahashi, K.; Taira, K.; Taira, M.; Ohno, A.; Sakugawa, H.; Arai, M.; Mishiro, S. Hepatitis E virus infection in wild mongooses of Okinawa, Japan: Demonstration of anti-HEV antibodies and a full-genome nucleotide sequence. Hepatol. Res. 2006, 34, 137–140. [Google Scholar] [CrossRef]
- Zhao, C.; Ma, Z.; Harrison, T.J.; Feng, R.; Zhang, C.; Qiao, Z.; Fan, J.; Ma, H.; Li, M.; Song, A.; et al. A novel genotype of hepatitis E virus prevalent among farmed rabbits in China. J. Med. Virol. 2009, 81, 1371–1379. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.L.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New Hepatitis E Virus Genotype in Camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.P.; Teng, J.L.; Cao, K.-Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.-C.; Wong, E.Y.; et al. New Hepatitis E Virus Genotype in Bactrian Camels, Xinjiang, China. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.-J.; Norder, H.; Okamoto, H.; Van Der Poel, W.H.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Lee, G.-H.; Tan, B.-H.; Teo, E.C.-Y.; Lim, S.-G.; Dan, Y.-Y.; Wee, A.; Aw, P.P.K.; Zhu, Y.; Hibberd, M.; Tan, C.-K.; et al. Chronic Infection With Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izopet, J.; Dubois, M.; Bertagnoli, S.; Lhomme, S.; Marchandeau, S.; Boucher, S.; Kamar, N.; Abravanel, F.; Guérin, J.-L. Hepatitis E Virus Strains in Rabbits and Evidence of a Closely Related Strain in Humans, France. Emerg. Infect. Dis. 2012, 18, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.; Chan, J.F.; Zhao, P.S.; Chan, W.; Poon, R.W.; Tsoi, H.; et al. Transmission of Rat Hepatitis E Virus Infection to Humans in Hong Kong: A Clinical and Epidemiological Analysis. Hepatology 2020, 73, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Harms, D.; Yang, X.-L.; Bock, C.-T. Orthohepevirus C: An Expanding Species of Emerging Hepatitis E Virus Variants. Pathogens 2020, 9, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.-C.; Yoshimatsu, K.; Yasuda, S.P.; Arikawa, J.; Koma, T.; Kataoka, M.; Ami, Y.; Suzaki, Y.; Mai, L.T.Q.; Hoa, N.T.; et al. Characterization of self-assembled virus-like particles of rat hepatitis E virus generated by recombinant baculoviruses. J. Gen. Virol. 2011, 92, 2830–2837. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-C.; Bai, H.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Takahashi, K.; Mishiro, S.; Takeda, N.; Wakita, T. Genotype 5 Hepatitis E Virus Produced by a Reverse Genetics System Has the Potential for Zoonotic Infection. Hepatol. Commun. 2018, 3, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Takeda, N.; Muramatsu, M.; Li, T.-C. Generation of a Bactrian camel hepatitis E virus by a reverse genetics system. J. Gen. Virol. 2021, 102, 001618. [Google Scholar] [CrossRef]
- Tanaka, T.; Takahashi, M.; Kusano, E.; Okamoto, H. Development and evaluation of an efficient cell-culture system for Hepatitis E virus. J. Gen. Virol. 2007, 88 Pt 3, 903–911. [Google Scholar] [CrossRef]
- Li, T.-C.; Zhou, X.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Nakamura, T.; Takeda, N.; Wakita, T. Production of infectious dromedary camel hepatitis E virus by a reverse genetic system: Potential for zoonotic infection. J. Hepatol. 2016, 65, 1104–1111. [Google Scholar] [CrossRef]
- Zhang, W.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Jirintai, S.; Takahashi, M.; Okamoto, H.; Takeda, N.; Muramatsu, M.; Li, T.; et al. Persistent infection with a rabbit hepatitis E virus created by a reverse genetics system. Transbound. Emerg. Dis. 2020, 68, 615–625. [Google Scholar] [CrossRef]
- Guo, Y.; Yang, F.; Xu, X.; Feng, M.; Liao, Y.; He, Z.; Takeda, N.; Muramatsu, M.; Li, Q.; Li, T.-C. Immunization of human hepatitis E viruses conferred protection against challenge by a camel hepatitis E virus. Vaccine 2020, 38, 7316–7322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Takeda, N.; Muramatsu, M.; Li, T.-C. High Prevalence of Hepatitis E Virus Infection in Imported Cynomolgus Monkeys in Japan. Jpn. J. Infect. Dis. 2019, 72, 429–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Duan, S.; Guo, Y.; Li, Y.; Yoshizaki, S.; Takeda, N.; Wakita, T.; Muramatsu, M.; Zhao, Y.; He, Z.; et al. Current status of hepatitis E virus infection at a rhesus monkey farm in China. Veter. Microbiol. 2019, 230, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Sayed, I.M.; Meuleman, P. Updates in Hepatitis E virus (HEV) field; lessons learned from human liver chimeric mice. Rev. Med. Virol. 2019, 30, e2086. [Google Scholar] [CrossRef]
- Li, S.; He, Q.; Yan, L.; Li, M.; Liang, Z.; Shu, J.; Zhang, F.; Wang, L. Infectivity and pathogenicity of different hepatitis E virus genotypes/subtypes in rabbit model. Emerg. Microbes Infect. 2020, 9, 2697–2705. [Google Scholar] [CrossRef]
- Zhang, W.; Ami, Y.; Suzaki, Y.; Kataoka, M.; Takeda, N.; Muramatsu, M.; Li, T. A Cross-Species Transmission of a Camel-Derived Genotype 8 Hepatitis E Virus to Rabbits. Pathogens 2021, 10, 1374. [Google Scholar] [CrossRef]
- Li, T.-C.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Yasuda, S.P.; Yoshimatsu, K.; Arikawa, J.; Takeda, N.; Wakita, T. Susceptibility of laboratory rats against genotypes 1, 3, 4, and rat hepatitis E viruses. Veter. Microbiol. 2013, 163, 54–61. [Google Scholar] [CrossRef]
- Sun, Y.-S.; Li, Y.-J.; Xia, Y.; Xu, F.; Wang, W.-W.; Yang, Z.-N.; Lu, H.-J.; Chen, Z.-P.; Miao, Z.-P.; Liang, W.-F.; et al. Coxsackievirus A16 induced neurological disorders in young gerbils which could serve as a new animal model for vaccine evaluation. Sci. Rep. 2016, 6, 34299. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-X.; Zhang, B.; Feng, Y.; Xu, C.-P.; Jiang, J.-M.; Lu, Y.-Y. Establishment and characterization of an oral gerbil model for a non-mouse-adapted enterovirus 71 strain. Virus Res. 2018, 255, 117–126. [Google Scholar] [CrossRef]
- Hong, Y.; He, Z.-J.; Tao, W.; Fu, T.; Wang, Y.-K.; Chen, Y. Experimental infection of Z:ZCLA Mongolian gerbils with human hepatitis E virus. World J. Gastroenterol. 2015, 21, 862–867. [Google Scholar] [CrossRef]
- Li, W.; Sun, Q.; She, R.; Wang, D.; Duan, X.; Yin, J.; Ding, Y. Experimental infection of mongolian gerbils by a genotype 4 strain of swine hepatitis E virus. J. Med. Virol. 2009, 81, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-D.; Zhang, F.; Chen, C.; Peng, L.; Luo, W.-T.; Chen, R.; Xu, P.; Huang, Y.-W. Revisiting the Mongolian Gerbil Model for Hepatitis E Virus by Reverse Genetics. Microbiol. Spectr. 2022, 10, e0219321. [Google Scholar] [CrossRef]
- Li, T.-C.; Suzaki, Y.; Ami, Y.; Dhole, T.N.; Miyamura, T.; Takeda, N. Protection of cynomolgus monkeys against HEV infection by oral administration of recombinant hepatitis E virus-like particles. Vaccine 2004, 22, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-C.; Suzaki, Y.; Ami, Y.; Tsunemitsu, H.; Miyamura, T.; Takeda, N. Mice are Not Susceptible to Hepatitis E Virus Infection. J. Veter. Med. Sci. 2008, 70, 1359–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.-C.; Ami, Y.; Suzaki, Y.; Yasuda, S.P.; Yoshimatsu, K.; Arikawa, J.; Takeda, N.; Takaji, W. Characterization of Full Genome of Rat Hepatitis E Virus Strain from Vietnam. Emerg. Infect. Dis. 2013, 19, 115–118. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Jirintai, S.; Tanggis; Mulyanto; Suparyatmo, J.B.; Takahashi, M.; Kobayashi, T.; Nagashima, S.; Nishizawa, T.; Okamoto, H. Rat hepatitis E virus derived from wild rats (Rattus rattus) propagates efficiently in human hepatoma cell lines. Virus Res. 2014, 185, 92–102. [Google Scholar] [CrossRef]
- Li, T.C.; Yamakawa, Y.; Suzuki, K.; Tatsumi, M.; A Razak, M.; Uchida, T.; Takeda, N.; Miyamura, T. Expression and self-assembly of empty virus-like particles of hepatitis E virus. J. Virol. 1997, 71, 7207–7213. [Google Scholar] [CrossRef] [Green Version]
- Li, T.-C.; Yang, T.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Ishii, K.; Haga, K.; Nakamura, T.; Ochiai, S.; Takaji, W.; et al. Construction and characterization of an infectious cDNA clone of rat hepatitis E virus. J. Gen. Virol. 2015, 96, 1320–1327. [Google Scholar] [CrossRef]
- Yang, F.; Li, Y.; Li, Y.; Jin, W.; Duan, S.; Xu, H.; Zhao, Y.; He, Z.; Ami, Y.; Suzaki, Y.; et al. Experimental Cross-Species Transmission of Rat Hepatitis E Virus to Rhesus and Cynomolgus Monkeys. Viruses 2022, 14, 293. [Google Scholar] [CrossRef]
- Li, T.-C.; Yoshizaki, S.; Yang, T.; Kataoka, M.; Nakamura, T.; Ami, Y.; Yuriko, S.; Takeda, N.; Wakita, T. Production of infectious ferret hepatitis E virus in a human hepatocarcinoma cell line PLC/PRF/5. Virus Res. 2016, 213, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Li, W.; Guan, D.; Su, J.; Ke, C.; Ami, Y.; Suzaki, Y.; Takeda, N.; Muramatsu, M.; Li, T.-C. Characterization of a Novel Rat Hepatitis E Virus Isolated from an Asian Musk Shrew (Suncus murinus). Viruses 2020, 12, 715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Kataoka, M.; Doan, H.Y.; Wu, F.-T.; Takeda, N.; Muramatsu, M.; Li, T.-C. Isolation and Characterization of Hepatitis E Virus Subtype 4b Using a PLC/PRF/5 Cell-Derived Cell Line Resistant to Porcine Sapelovirus Infection. Jpn. J. Infect. Dis. 2021, 74, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Bendall, R.P.; Peron, J.M.; Cintas, P.; Prudhomme, L.; Mansuy, J.M.; Rostaing, L.; Keane, F.; Ijaz, S.; Izopet, J.; et al. Hepatitis E Virus and Neurologic Disorders. Emerg. Infect. Dis. 2011, 17, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Pischke, S.; Hartl, J.; Pas, S.D.; Lohse, A.W.; Jacobs, B.C.; Van der Eijk, A.A. Hepatitis E virus: Infection beyond the liver? J. Hepatol. 2017, 66, 1082–1095. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Shi, R.; Xiao, P.; Liu, T.; She, R.; Wu, Q.; An, J.; Hao, W.; Soomro, M. Hepatitis E Virus Induces Brain Injury Probably Associated With Mitochondrial Apoptosis. Front. Cell. Infect. Microbiol. 2019, 9, 433. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HEV Strain | Gerbil 1 | Gerbil 2 | Gerbil 3 |
|---|---|---|---|
| HEV-1 | 45 | 66 | 47 |
| HEV-3 | 54 | 49 | 69 |
| HEV-4 | 63 | 69 | 78 |
| HEV-5 | 59 | 58 | 78 |
| HEV-7 | 46 | 46 | 63 |
| HEV-8 | 54 | 66 | 60 |
| Rabbit HEV | 42 | 47 | 48 |
| Rat HEV | 57 | 62 | 59 |
| PBS | 53 | 57 | 60 |
| HEV Strain | Gerbil 1 | Gerbil 2 | Gerbil 3 |
|---|---|---|---|
| HEV-4 | 59 | 58 | 98 |
| Rat HEV | 92 | 70 | 61 |
| HEV Strain | Gerbil 1 | Gerbil 2 | Gerbil 3 |
|---|---|---|---|
| HEV-4 | 53 | 47 | 56 |
| HEV-5 | 56 | 70 | 61 |
| Rat HEV | 56 | 50 | 59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Ami, Y.; Suzaki, Y.; Doan, Y.H.; Muramatsu, M.; Li, T.-C. Mongolia Gerbils Are Broadly Susceptible to Hepatitis E Virus. Viruses 2022, 14, 1125. https://doi.org/10.3390/v14061125
Zhang W, Ami Y, Suzaki Y, Doan YH, Muramatsu M, Li T-C. Mongolia Gerbils Are Broadly Susceptible to Hepatitis E Virus. Viruses. 2022; 14(6):1125. https://doi.org/10.3390/v14061125
Chicago/Turabian StyleZhang, Wenjing, Yasushi Ami, Yuriko Suzaki, Yen Hai Doan, Masamichi Muramatsu, and Tian-Cheng Li. 2022. "Mongolia Gerbils Are Broadly Susceptible to Hepatitis E Virus" Viruses 14, no. 6: 1125. https://doi.org/10.3390/v14061125
APA StyleZhang, W., Ami, Y., Suzaki, Y., Doan, Y. H., Muramatsu, M., & Li, T.-C. (2022). Mongolia Gerbils Are Broadly Susceptible to Hepatitis E Virus. Viruses, 14(6), 1125. https://doi.org/10.3390/v14061125