
Are Hamsters a Suitable Model for Evaluating the Immunogenicity of RBD-Based Anti-COVID-19 Subunit Vaccines?

 , , , , , , , ,
, , , , , , , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction, Recombinant Protein Expression, and Purification
2.2. Animal Immunization
2.3. IgY Preparation
2.4. SARS-CoV-2 Lysate Preparation
2.5. ELISA Assays
2.6. Viral Neutralization Test
2.7. Statistical Analysis
3. Results
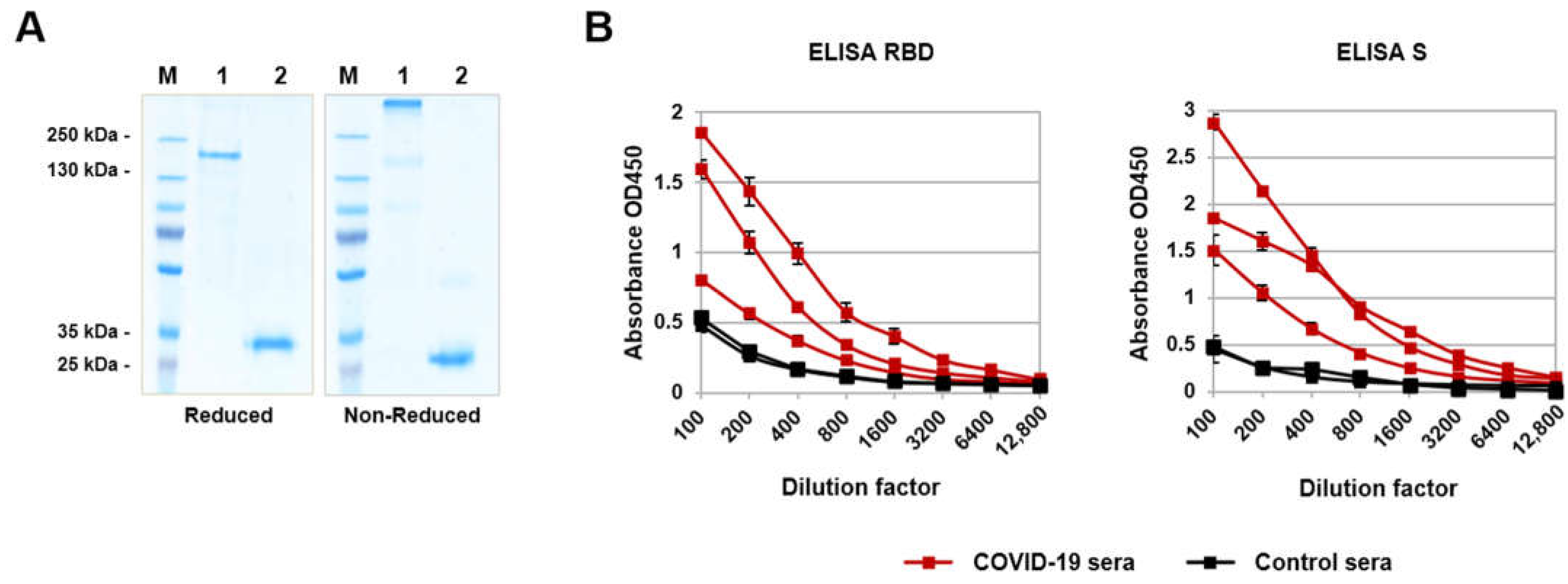
3.1. Recombinant RBD Development and Characterization
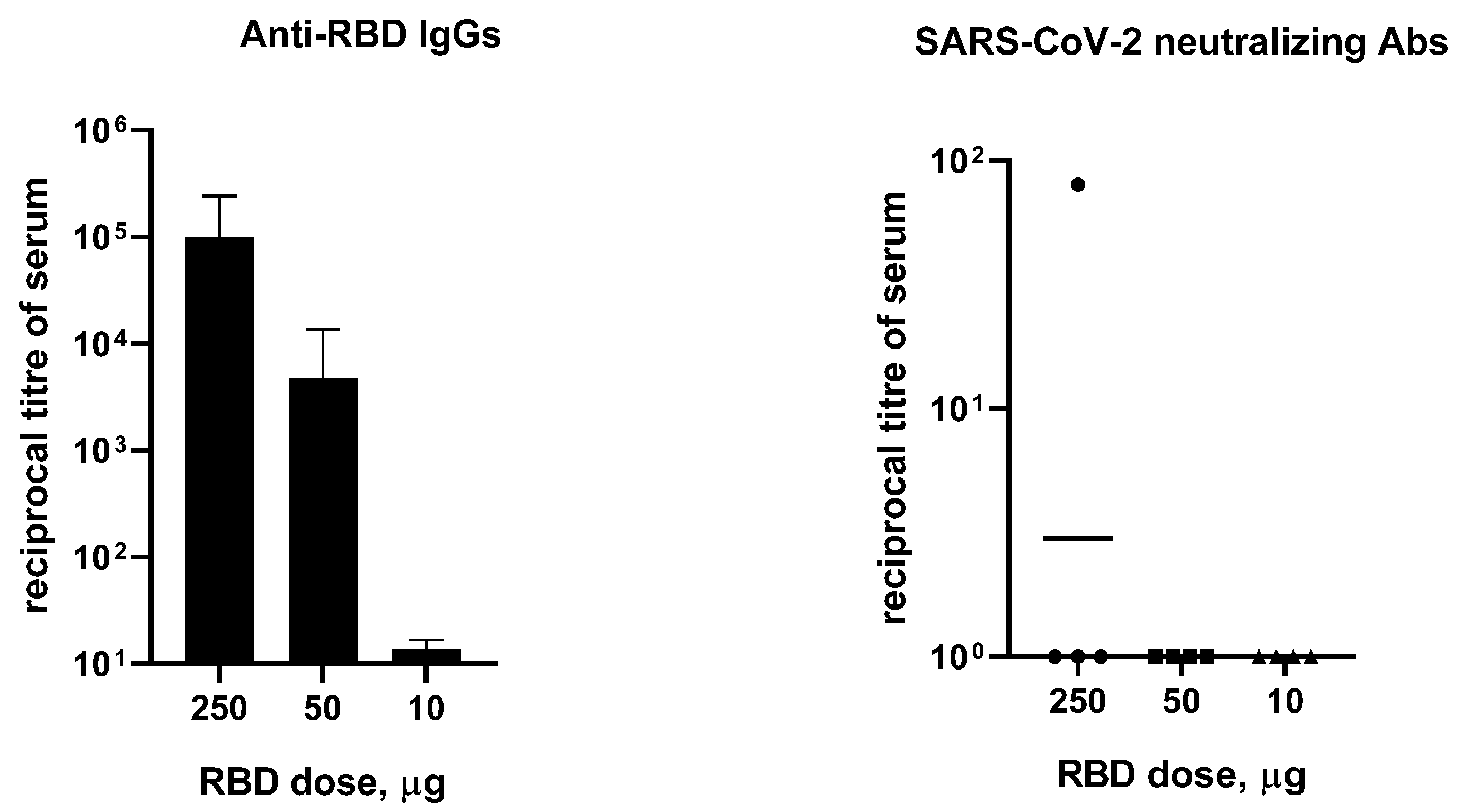
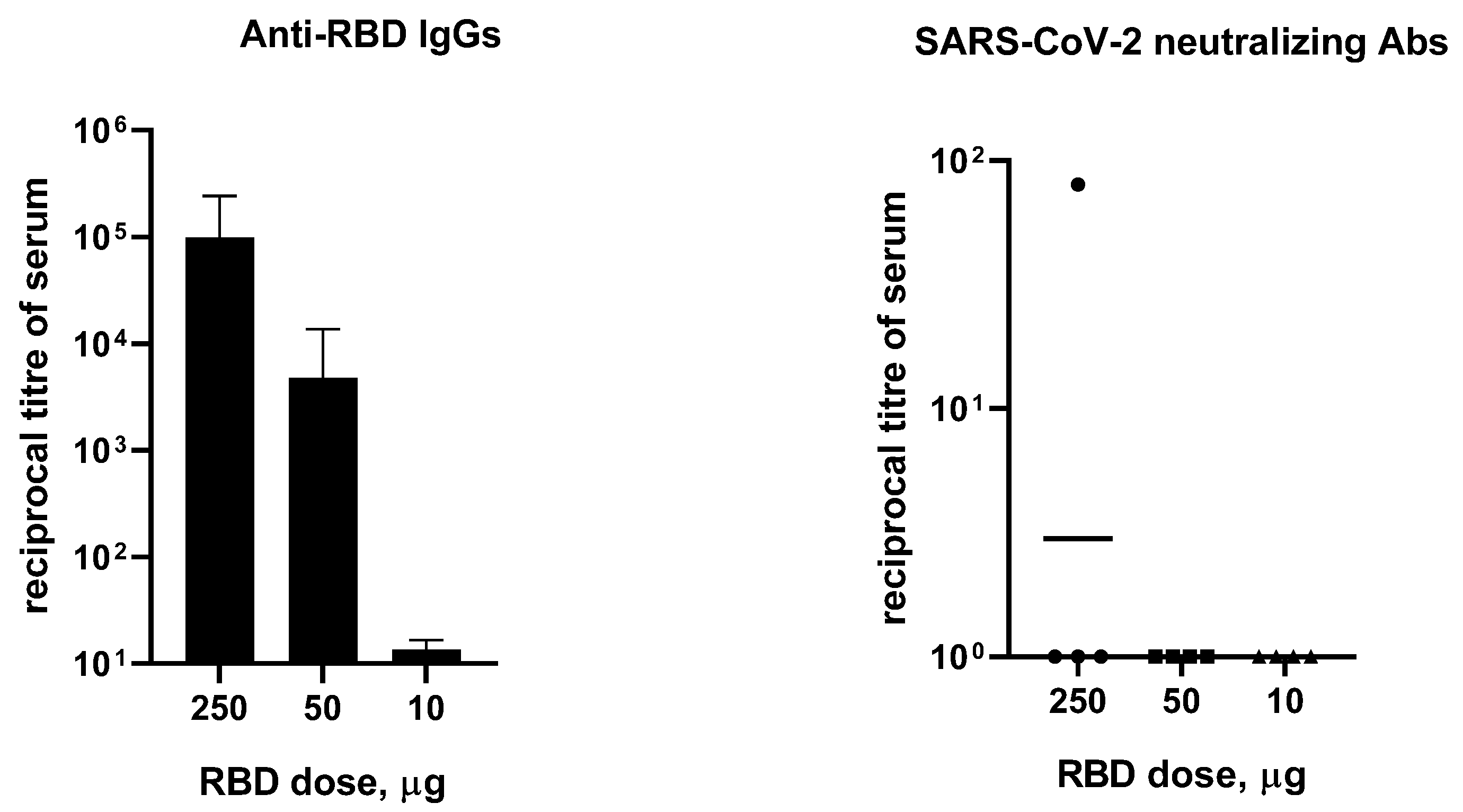
3.2. Immunogenicity of RBD in Mice, Hamsters, Ferrets, and Rabbits
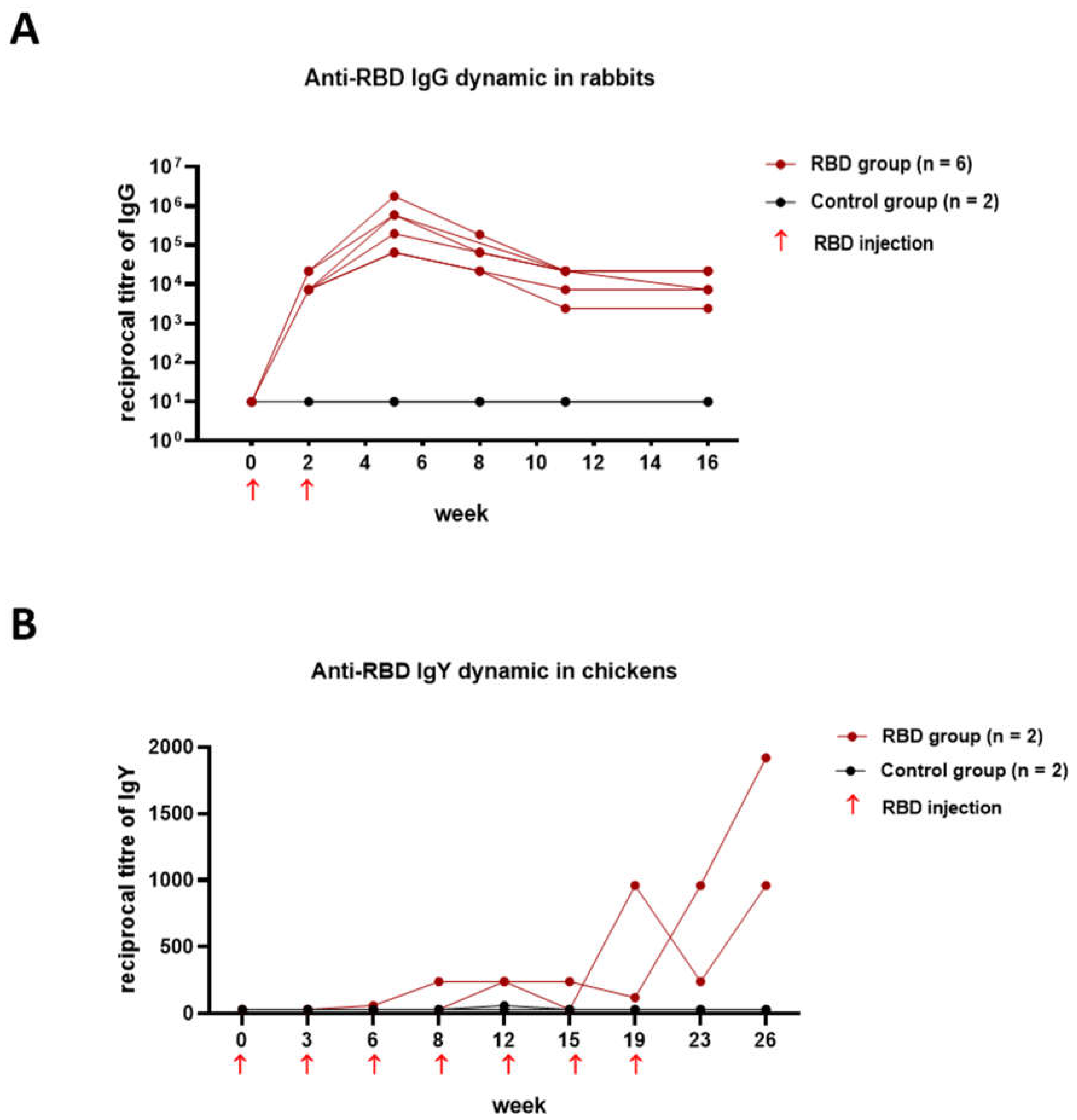
3.3. Dynamic of Humoral Immune Responses in Rabbits and Chickens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barton, M.I.; MacGowan, S.A.; Kutuzov, M.A.; Dushek, O.; Barton, G.J.; van der Merwe, P.A. Effects of common mutations in the SARS-CoV-2 Spike RBD and its ligand, the human ACE2 receptor on binding affinity and kinetics. Elife 2021, 10, e70658. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Sun, Q. Antibodies and Vaccines Target RBD of SARS-CoV-2. Front. Mol. Biosci. 2021, 8, 671633. [Google Scholar] [CrossRef] [PubMed]
- Kleanthous, H.; Silverman, J.M.; Makar, K.W.; Yoon, I.K.; Jackson, N.; Vaughn, D.W. Scientific rationale for developing potent RBD-based vaccines targeting COVID-19. NPJ Vaccines 2021, 6, 128. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef]
- Munoz-Fontela, C.; Dowling, W.E.; Funnell, S.G.P.; Gsell, P.S.; Riveros-Balta, A.X.; Albrecht, R.A.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Güler, A.; et al. BNT162b vaccines protect rhesus macaques from SARS-CoV-2 Check for updates. Nature 2021, 592, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Oh, H.; Park, Y.W.; Kwak, H.W.; Oh, E.Y.; Park, H.J.; Kang, K.W.; Kim, G.; Koo, B.S.; Hwang, E.H.; et al. Immunization with RBD-P2 and N protects against SARS-CoV-2 in nonhuman primates. Sci. Adv. 2021, 7, eabg7156. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Gagne, M.; Wagner, D.A.; Connell, S.O.; Narpala, S.R.; Flebbe, D.R.; Andrew, S.F.; Davis, R.L.; Flynn, B.; Johnston, T.S.; et al. Protection against SARS-CoV-2 Beta variant in mRNA-1273 vaccine-boosted nonhuman primates. Science 2021, 374, 1343–1353. [Google Scholar] [CrossRef]
- Yu, J.; Tostanosk, L.H.; Peter, L.; Mercad, N.B.; McMahan, K.; Mahrokhia, S.H.; Nkolol, J.P.; Liu, J.; Li, Z.; Chandrashekar, A.; et al. DNA vaccine protection against SARS-CoV-2 in rhesus macaques. Science 2020, 369, 806–811. [Google Scholar] [CrossRef]
- Lu, Y.R.; Wang, L.N.; Jin, X.; Chen, Y.N.; Cong, C.; Yuan, Y.; Li, Y.C.; Tang, W.D.; Li, H.X.; Wu, X.T.; et al. A Preliminary Study on the Feasibility of Gene Expression Profile of Rhesus Monkey Detected With Human Microarray. Transplant. Proc. 2008, 40, 598–602. [Google Scholar] [CrossRef]
- Sutton, T.C.; Subbarao, K. Development of animal models against emerging coronaviruses: From SARS to MERS coronavirus. Virology 2015, 479–480, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host. Microbe 2020, 27, 704–709.e702. [Google Scholar] [CrossRef] [PubMed]
- Shou, S.; Liu, M.; Yang, Y.; Kang, N.; Song, Y.; Tan, D.; Liu, N.; Wang, F.; Liu, J.; Xie, Y. Animal Models for COVID-19: Hamsters, Mouse, Ferret, Mink, Tree Shrew, and Non-human Primates. Front. Microbiol. 2021, 12, 626553. [Google Scholar] [CrossRef]
- Richard, M.; Kok, A.; de Meulder, D.; Bestebroer, T.M.; Lamers, M.M.; Okba, N.M.A.; Fentener van Vlissingen, M.; Rockx, B.; Haagmans, B.L.; Koopmans, M.P.G.; et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat. Commun. 2020, 11, 3496. [Google Scholar] [CrossRef]
- Li, L.; Honda-Okubo, Y.; Huang, Y.; Jang, H.; Carlock, M.A.; Baldwin, J.; Piplani, S.; Bebin-Blackwell, A.G.; Forgacs, D.; Sakamoto, K.; et al. Immunisation of ferrets and mice with recombinant SARS-CoV-2 spike protein formulated with Advax-SM adjuvant protects against COVID-19 infection. Vaccine 2021, 40, 5940–5953. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Li, K.; Rowe, D.K.; Diaz, M.C.H.; Griffin, E.F.; Beavis, A.C.; Johnson, S.K.; Padykula, I.; Jones, C.A.; Briggs, K.; et al. Protection of K18-hACE2 mice and ferrets against SARS-CoV-2 challenge by a single-dose mucosal immunization with a parainfluenza virus 5-based COVID-19 vaccine. Sci. Adv. 2021, 7, eabi5246. [Google Scholar] [CrossRef]
- Sawatzki, K.; Hill, N.J.; Puryear, W.B.; Foss, A.D.; Stone, J.J.; Runstadler, J.A. Host barriers to SARS-CoV-2 demonstrated by ferrets in a high-exposure domestic setting. Proc. Natl. Acad. Sci. USA 2021, 118, e2025601118. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Felipe, L.; Vercruysse, T.; Sharma, S.; Ma, J.; Lemmens, V.; Van Looveren, D.; Arkalagud Javarappa, M.P.; Boudewijns, R.; Malengier-Devlies, B.; Liesenborghs, L.; et al. A single-dose live-attenuated YF17D-vectored SARS-CoV-2 vaccine candidate. Nature 2021, 590, 320–325. [Google Scholar] [CrossRef]
- Boudewijns, R.; Pérez, P.; Lázaro-Frías, A.; Van Looveren, D.; Vercruysse, T.; Thibaut, H.J.; Weynand, B.; Coelmont, L.; Neyts, J.; Astorgano, D.; et al. MVA-CoV2-S Vaccine Candidate Neutralizes Distinct Variants of Concern and Protects Against SARS-CoV-2 Infection in Hamsters. Front. Immunol. 2022, 13, 845969. [Google Scholar] [CrossRef]
- Chiba, S.; Frey, S.J.; Halfmann, P.J.; Kuroda, M.; Maemura, T.; Yang, J.E.; Wright, E.R.; Kawaoka, Y.; Kane, R.S. Multivalent nanoparticle-based vaccines protect hamsters against SARS-CoV-2 after a single immunization. Commun. Biol. 2021, 4, 597. [Google Scholar] [CrossRef] [PubMed]
- Brocato, R.L.; Kwilas, S.A.; Kim, R.K.; Zeng, X.; Principe, L.M.; Smith, J.M.; Hooper, J.W. Protective efficacy of a SARS-CoV-2 DNA vaccine in wild-type and immunosuppressed Syrian hamsters. npj Vaccines 2021, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, P.; Fan, T.; Wu, Y.; Zhang, J.; Shi, X.; Shang, W.; Fang, L.; Jiang, X.; Shi, J.; et al. Immunoglobulin fragment F(ab’)2 against RBD potently neutralizes SARS-CoV-2 in vitro. Antivir. Res. 2020, 182, 104868. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.-Y.; Lien, C.E.; Lin, Y.-J.; Lin, M.-Y.; Wu, C.-C.; Tang, W.-H.; Campbell, J.D.; Traquina, P.; Chuang, Y.-S.; Liu, L.T.-C.; et al. Protection of Hamsters Challenged with SARS-CoV-2 Delta Variant after Two Doses of Adjuvanted SARS-CoV-2 Stabilized Prefusion Spike Protein (S-2P) and a Single Dose of Beta Variant S-2P. J. Infect. Dis. 2022, jiac153. [Google Scholar] [CrossRef]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor Recognition by the Novel Coronavirus from Wuhan: An Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J. Virol. 2020, 94, e00127-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y.; et al. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef]
- Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Edwards, C.E.; Martinez, D.R.; Montgomery, S.A.; West, A.; Yount, B.L.; Hou, Y.J.; Adams, L.E.; et al. A mouse-adapted model of SARS-CoV-2 to test COVID-19 countermeasures. Nature 2020, 586, 560–566. [Google Scholar] [CrossRef]
- Jarnagin, K.; Alvarez, O.; Shresta, S.; Webb, D.R. Animal models for SARS-Cov2/Covid19 research-A commentary. Biochem. Pharmacol. 2021, 188, 114543. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, X.; Yuan, L.; Wang, S.; Zhang, Y.; Xiong, H.; Chen, R.; Ma, J.; Qi, R.; Nie, M.; et al. Sterilizing immunity against SARS-CoV-2 in hamsters conferred by a novel recombinant subunit vaccine. bioRxiv 2020. [Google Scholar] [CrossRef]
- Merkuleva, I.A.; Shcherbakov, D.N.; Borgoyakova, M.B.; Shanshin, D.V.; Rudometov, A.P.; Karpenko, L.I.; Belenkaya, S.V.; Isaeva, A.A.; Nesmeyanova, V.S.; Kazachinskaia, E.I.; et al. Comparative Immunogenicity of the Recombinant Receptor-Binding Domain of Protein S SARS-CoV-2 Obtained in Prokaryotic and Mammalian Expression Systems. Vaccines 2022, 10, 96. [Google Scholar] [CrossRef]
- Polson, A.; von Wechmar, M.B.; van Regenmortel, M.H. Isolation of viral IgY antibodies from yolks of immunized hens. Immunol. Commun. 1980, 9, 475–493. [Google Scholar] [CrossRef]
- Chepurnov, A.A.; Kazachinskaya, E.I.; Kazachkova, E.A.; Sharshov, K.A.; Kononova, Y.V.; Shelemba, A.A.; Alekseev, A.Y.; Gulyeva, M.A.; Voevoda, M.I.; Shestopalov, A.M. Development of a Purified Viral Preparation for Studies of COVID-19 (SARS-CoV-2) Biology. Bull. Exp. Biol. Med. 2021, 172, 49–52. [Google Scholar] [CrossRef]
- Borgoyakova, M.B.; Karpenko, L.I.; Rudometov, A.P.; Volosnikova, E.A.; Merkuleva, I.A.; Starostina, E.V.; Zadorozhny, A.M.; Isaeva, A.A.; Nesmeyanova, V.S.; Shanshin, D.V.; et al. Self-Assembled Particles Combining SARS-CoV-2 RBD Protein and RBD DNA Vaccine Induce Synergistic Enhancement of the Humoral Response in Mice. Int. J. Mol. Sci. 2022, 23, 2188. [Google Scholar] [CrossRef] [PubMed]
- Dalvie, N.C.; Rodriguez-Aponte, S.A.; Hartwell, B.L.; Tostanoski, L.H.; Biedermann, A.M.; Crowell, L.E.; Kaur, K.; Kumru, O.; Carter, L.; Yu, J.; et al. Engineered SARS-CoV-2 receptor binding domain improves immunogenicity in mice and elicits protective immunity in hamsters. bioRxiv 2021. [Google Scholar] [CrossRef]
- Muñoz-Fontela, C.; Widerspick, L.; Albrecht, R.A.; Beer, M.; Carroll, M.W.; de Wit, E.; Diamond, M.S.; Dowling, W.E.; Funnell, S.G.P.; García-Sastre, A.; et al. Advances and gaps in SARS-CoV-2 infection models. PLoS Pathog. 2022, 18, e1010161. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.T.; Fumagalli, M.J.; Hojo-Souza, N.S.; Azevedo, P.; Salazar, N.; Rattis, B.; Ramos, S.G.; Faustino, L.; Almeida, G.G.; Oliveira, L.I.; et al. Neutralizing antibody-independent immunity to SARS-CoV-2 in hamsters and hACE-2 transgenic mice immunized with a RBD/Nucleocapsid fusion protein. bioRxiv 2021. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef] [PubMed]
- Mykytyn, A.Z.; Lamers, M.M.; Okba, N.M.A.; Breugem, T.I.; Schipper, D.; van den Doel, P.B.; van Run, P.; van Amerongen, G.; de Waal, L.; Koopmans, M.P.G.; et al. Susceptibility of rabbits to SARS-CoV-2. Emerg. Microbes Infect. 2021, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.; de Riols de Fonclare, D.; Garcia, D.; Beurlet, S.; Becquart, P.; Rosolen, S.G.; Briend-Marchal, A.; Leroy, E.M. First Evidence of Natural SARS-CoV-2 Infection in Domestic Rabbits. Vet. Sci. 2022, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E.; Suarez, D.L.; Spackman, E.; Tumpey, T.M.; Beck, J.R.; Erdman, D.; Rollin, P.E.; Ksiazek, T.G. Domestic Poultry and SARS Coronavirus, Southern China. Emerg. Infect. Dis. 2004, 10, 914–916. [Google Scholar] [CrossRef]
- Shen, H.; Cai, Y.; Zhang, H.; Wu, J.; Ye, L.; Yang, P.; Lin, X.; Jiang, S.; Liao, M. Anti-SARS-CoV-2 IgY Isolated from Egg Yolks of Hens Immunized with Inactivated SARS-CoV-2 for Immunoprophylaxis of COVID-19. Virol. Sin. 2021, 36, 1080–1082. [Google Scholar] [CrossRef]
- Wongso, H.; Mahendra, I.; Arnafia, W.; Idar, I.; Yusuf, M.; Achmad, A.; Holik, H.A.; Kurniawan, A.; Halimah, I.; Sriyani, M.E.; et al. Preclinical Evaluation of Chicken Egg Yolk Antibody (IgY) Anti-RBD Spike SARS-CoV-2—A Candidate for Passive Immunization against COVID-19. Vaccines 2022, 10, 128. [Google Scholar] [CrossRef]
- Bao, L.; Zhang, C.; Lyu, J.; Yi, P.; Shen, X.; Tang, B.; Zhao, H.; Ren, B.; Kuang, Y.; Zhou, L.; et al. Egg yolk immunoglobulin (IgY) targeting SARS-CoV-2 S1 as potential virus entry blocker. J. Appl. Microbiol. 2022, 132, 2421–2430. [Google Scholar] [CrossRef]
- Frumkin, L.R.; Lucas, M.; Scribner, C.L.; Ortega-Heinly, N.; Rogers, J.; Yin, G.; Hallam, T.J.; Yam, A.; Bedard, K.; Begley, R.; et al. Egg-derived anti-SARS-CoV-2 immunoglobulin Y (IgY) with broad variant activity as intranasal prophylaxis against COVID-19: Preclinical studies and randomized controlled phase 1 clinical trial. medRxiv 2022. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merkuleva, I.A.; Shcherbakov, D.N.; Borgoyakova, M.B.; Isaeva, A.A.; Nesmeyanova, V.S.; Volkova, N.V.; Aripov, V.S.; Shanshin, D.V.; Karpenko, L.I.; Belenkaya, S.V.; et al. Are Hamsters a Suitable Model for Evaluating the Immunogenicity of RBD-Based Anti-COVID-19 Subunit Vaccines? Viruses 2022, 14, 1060. https://doi.org/10.3390/v14051060
Merkuleva IA, Shcherbakov DN, Borgoyakova MB, Isaeva AA, Nesmeyanova VS, Volkova NV, Aripov VS, Shanshin DV, Karpenko LI, Belenkaya SV, et al. Are Hamsters a Suitable Model for Evaluating the Immunogenicity of RBD-Based Anti-COVID-19 Subunit Vaccines? Viruses. 2022; 14(5):1060. https://doi.org/10.3390/v14051060
Chicago/Turabian StyleMerkuleva, Iuliia A., Dmitry N. Shcherbakov, Mariya B. Borgoyakova, Anastasiya A. Isaeva, Valentina S. Nesmeyanova, Natalia V. Volkova, Vazirbek S. Aripov, Daniil V. Shanshin, Larisa I. Karpenko, Svetlana V. Belenkaya, and et al. 2022. "Are Hamsters a Suitable Model for Evaluating the Immunogenicity of RBD-Based Anti-COVID-19 Subunit Vaccines?" Viruses 14, no. 5: 1060. https://doi.org/10.3390/v14051060
APA StyleMerkuleva, I. A., Shcherbakov, D. N., Borgoyakova, M. B., Isaeva, A. A., Nesmeyanova, V. S., Volkova, N. V., Aripov, V. S., Shanshin, D. V., Karpenko, L. I., Belenkaya, S. V., Kazachinskaia, E. I., Volosnikova, E. A., Esina, T. I., Sergeev, A. A., Titova, K. A., Konyakhina, Y. V., Zaykovskaya, A. V., Pyankov, O. V., Kolosova, E. A., ... Ilyichev, A. A. (2022). Are Hamsters a Suitable Model for Evaluating the Immunogenicity of RBD-Based Anti-COVID-19 Subunit Vaccines? Viruses, 14(5), 1060. https://doi.org/10.3390/v14051060