
The Beauty of Bacteriophage T4 Research: Lindsay W. Black and the T4 Head Assembly



Abstract
1. Introduction
2. From a Dodecameric Ring to a Complex DNA-Full Prolate Icosahedron—The Molecular Assembly Line That Produces the T4 Head
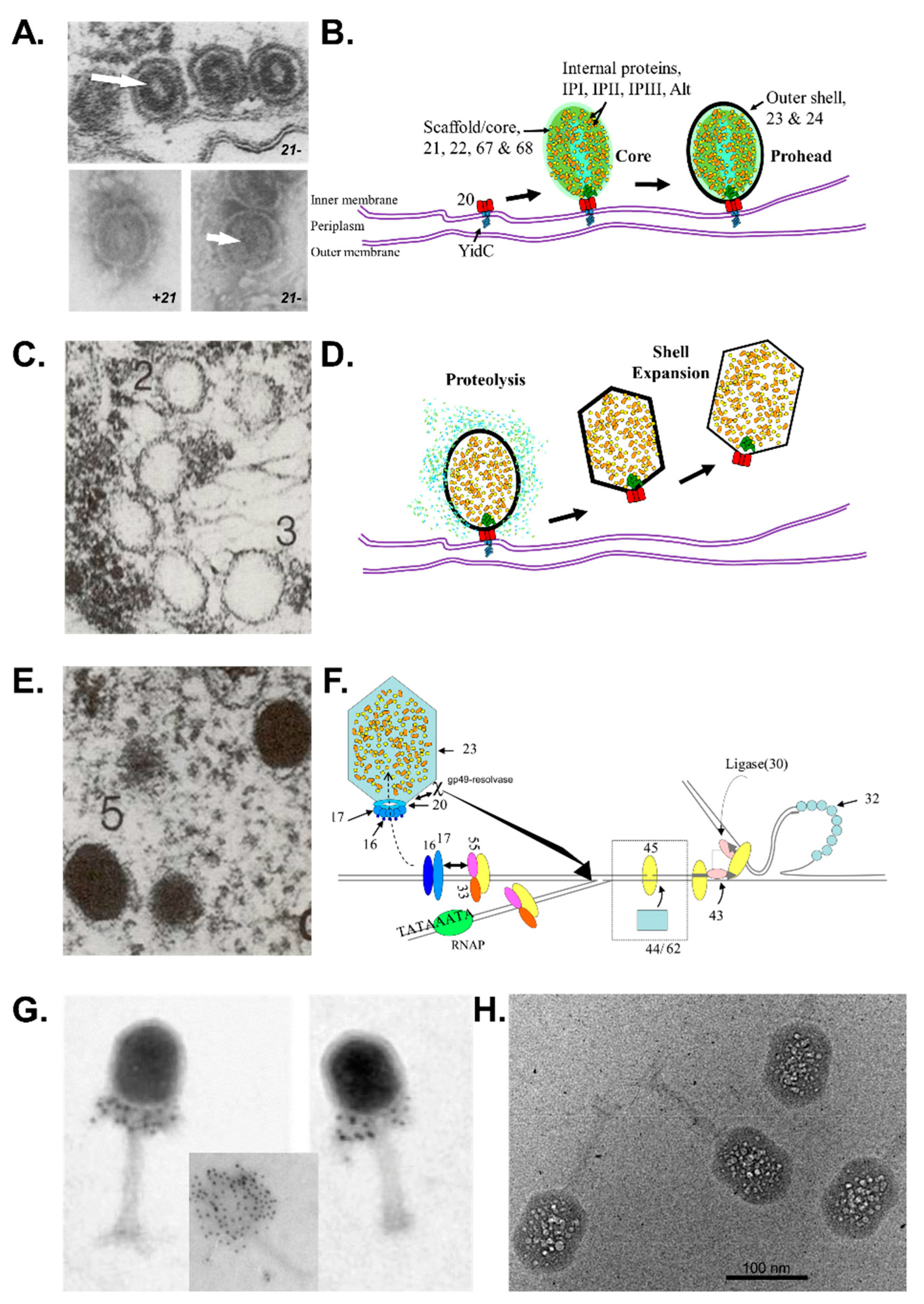
2.1. Overview of Major Steps in T4 Head Assembly
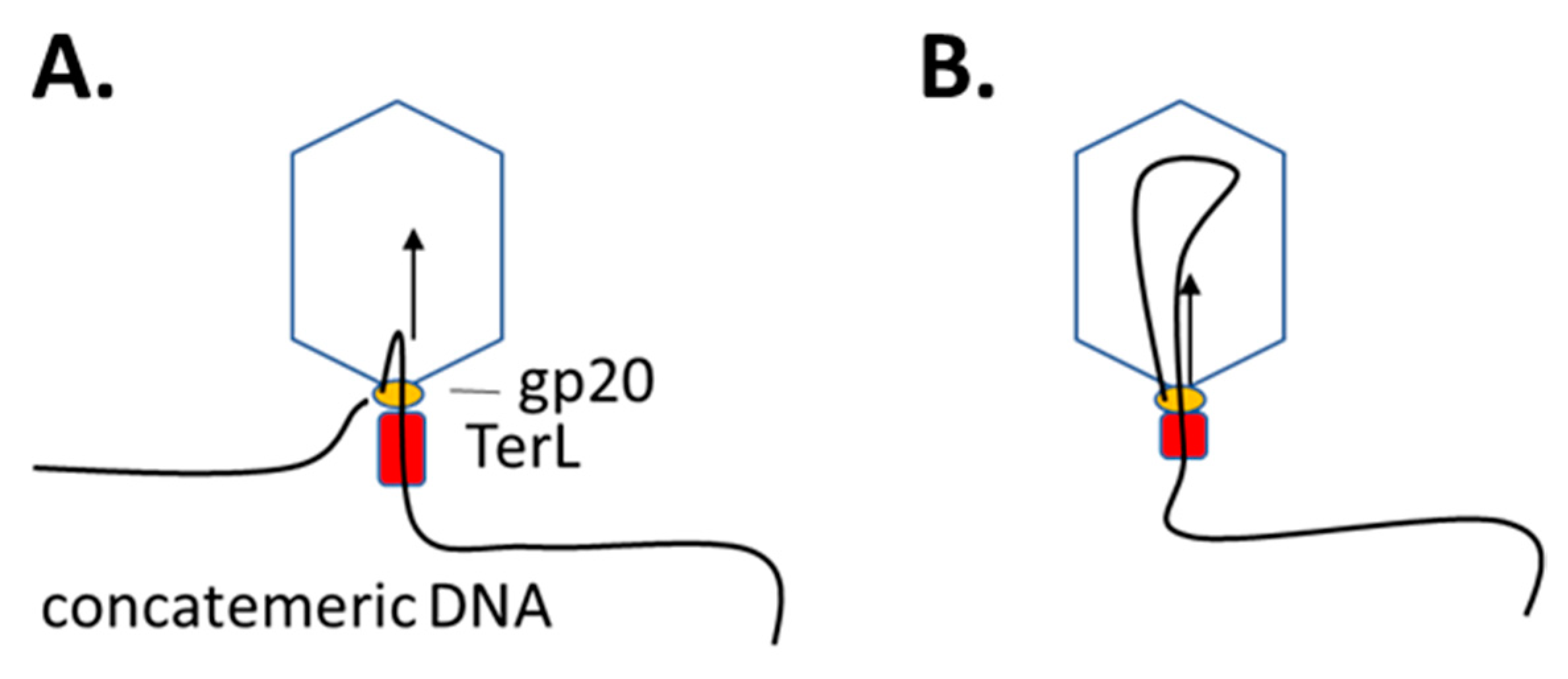
2.2. Lifting the Secrets of T4 DNA Packaging
2.3. Mysterious Small T4 Internal Head Proteins
2.4. Role of IPI in Phage–Bacterial Host Arms Race
2.5. Proteolytic Maturation of T4 Head: A Cascade of Protein Conformational Changes
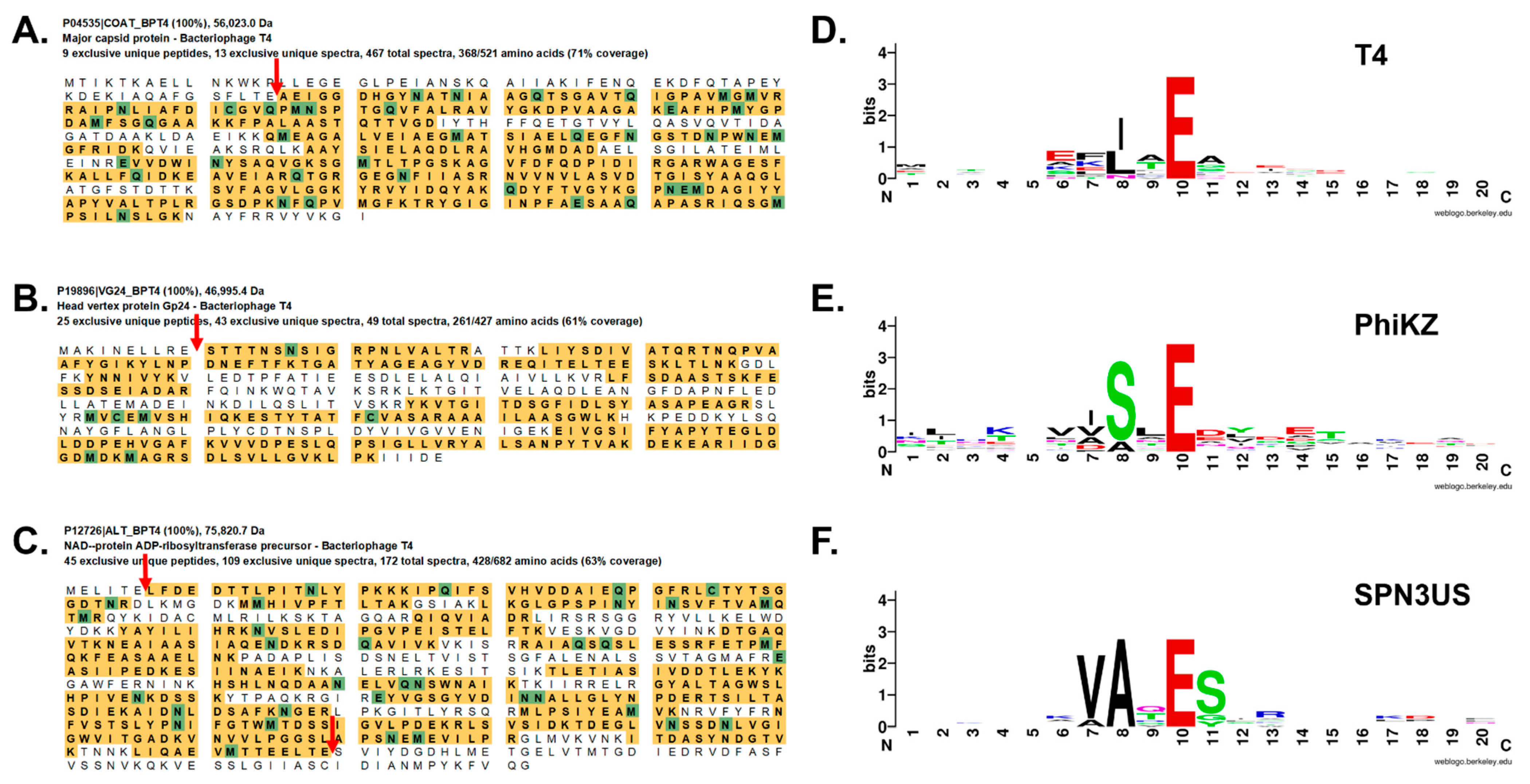
2.6. Targeting of Proteins into the T4 Phage Head
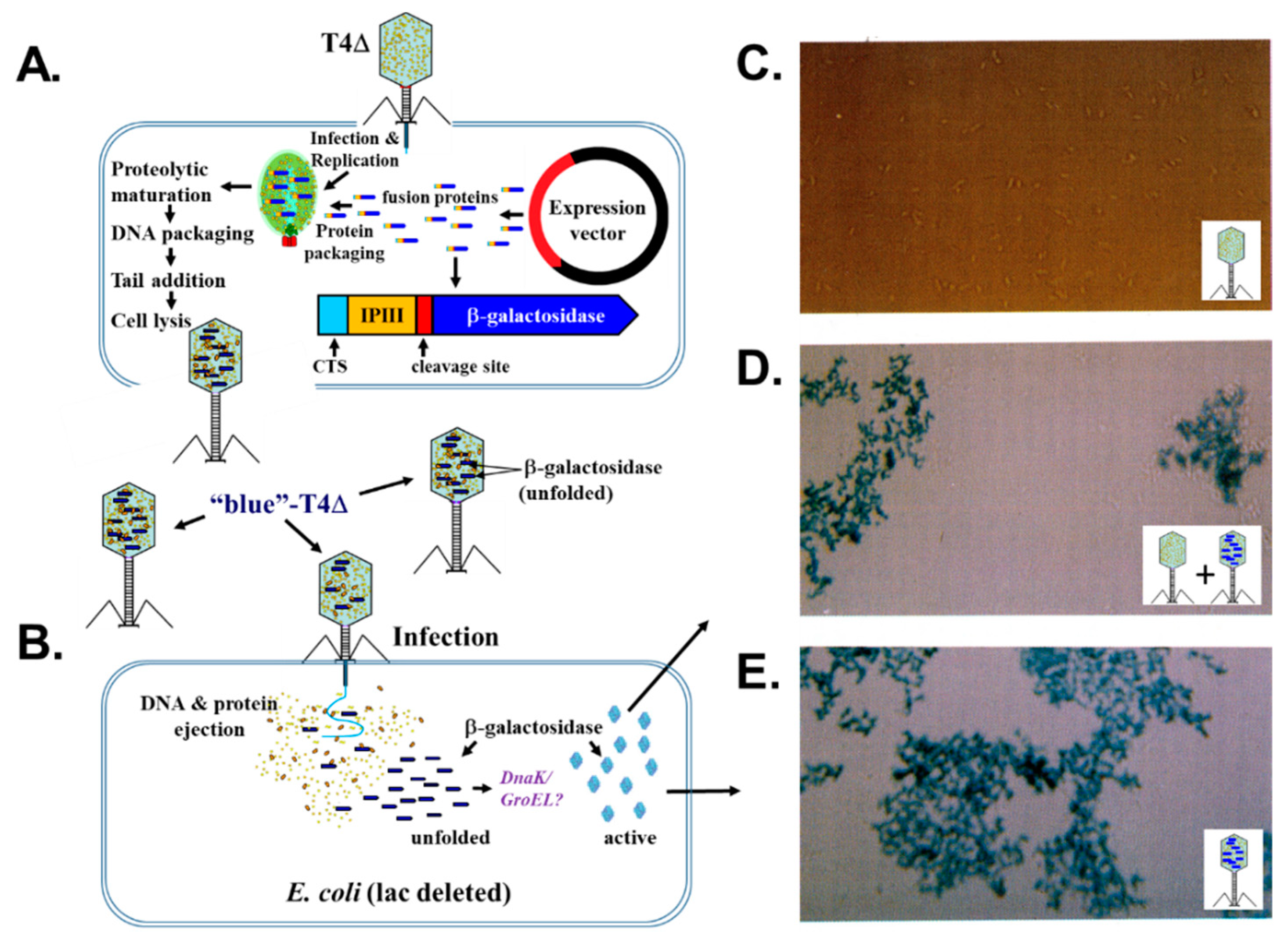
2.7. Fusion Proteins and Phage Display with Bacteriophage T4
3. The Ever-Expanding Value of T4 Phage for Elucidating Head Assembly Mechanisms in Other Phages
3.1. Billions of Head Assemblies Each Day Defined by Studies on T4
3.2. Shared Features of Head Maturation between T4 and Phages with Larger Icosahedral Capsids
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Black, L.W.; Rao, V.B. Structure, assembly, and DNA packaging of the bacteriophage T4 head. Adv. Virus Res. 2012, 82, 119–153. [Google Scholar] [CrossRef] [PubMed]
- Quinten, T.A.; Kuhn, A. Membrane Interaction of the Portal Protein gp20 of Bacteriophage T4. J. Virol. 2012, 86, 11107–11114. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, A.; Keller, B.; Maeder, M.; Traub, F. Prohead core of bacteriophage T4 can act as an intermediate in the T4 head assembly pathway. J. Virol. 1987, 61, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Black, L.W.; Showe, M.K.; Steven, A.C. Morphogenesis of the T4 head. In Molecular Biology of Bacteriophage T4; Karam, J.D., Ed.; ASM Press: Washington, DC, USA, 1994; pp. 218–258. [Google Scholar]
- Dixit, A.B.; Ray, K.; Thomas, J.A.; Black, L.W. The C-terminal domain of the bacteriophage T4 terminase docks on the prohead portal clip region during DNA packaging. Virology 2013, 446, 293–302. [Google Scholar] [CrossRef]
- van Driel, R.; Traub, F.; Showe, M.K. Probable localization of the bacteriophage T4 prehead proteinase zymogen in the center of the prehead core. J. Virol. 1980, 36, 220–223. [Google Scholar] [CrossRef]
- Black, L.W.; Thomas, J.A. Dealing With the Whole Head: Diversity and Function of Capsid Ejection Proteins in Tailed Phages. In Encyclopedia of Virology, 4th ed.; Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Oxford, UK, 2021; pp. 219–228. [Google Scholar]
- Black, L.W.; Peng, G. Mechanistic coupling of bacteriophage T4 DNA packaging to components of the replication-dependent late transcription machinery. J. Biol. Chem. 2006, 281, 25635–25643. [Google Scholar] [CrossRef]
- Baumann, R.G.; Mullaney, J.; Black, L.W. Portal fusion protein constraints on function in DNA packaging of bacteriophage T4. Mol. Microbiol. 2006, 61, 16–32. [Google Scholar] [CrossRef]
- Wu, W.; Cheng, N.; Black, L.W.; Dietz, H.; Steven, A.C. Biphasic Packing of DNA and Internal Proteins in Bacteriophage T4 Heads Revealed by Bubblegram Imaging. Viruses 2020, 12, 1282. [Google Scholar] [CrossRef]
- Black, L.W.; Silverman, D.J. Model for DNA packaging into bacteriophage T4 heads. J. Virol. 1978, 28, 643–655. [Google Scholar] [CrossRef]
- Jiang, X.; Jiang, H.; Li, C.; Wang, S.; Mi, Z.; An, X.; Chen, J.; Tong, Y. Sequence characteristics of T4-like bacteriophage IME08 benome termini revealed by high throughput sequencing. Virol. J. 2011, 8, 194. [Google Scholar] [CrossRef]
- Hsiao, C.L.; Black, L.W. DNA packaging and the pathway of bacteriophage T4 head assembly. Proc. Natl. Acad. Sci. USA 1977, 74, 3652–3656. [Google Scholar] [CrossRef] [PubMed]
- Ray, K.; Ma, J.X.; Oram, M.; Lakowicz, J.R.; Black, L.W. Single-Molecule and FRET Fluorescence Correlation Spectroscopy Analyses of Phage DNA Packaging: Colocalization of Packaged Phage T4 DNA Ends within the Capsid. J. Mol. Biol. 2010, 395, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Kondabagil, K.; Draper, B.; Alam, T.I.; Bowman, V.D.; Zhang, Z.; Hegde, S.; Fokine, A.; Rossmann, M.G.; Rao, V.B. The structure of the phage T4 DNA packaging motor suggests a mechanism dependent on electrostatic forces. Cell 2008, 135, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Serwer, P.; Jiang, W. Dualities in the analysis of phage DNA packaging motors. Bacteriophage 2012, 2, 239–255. [Google Scholar] [CrossRef][Green Version]
- Lin, H.; Rao, V.B.; Black, L.W. Analysis of capsid portal protein and terminase functional domains: Interaction sites required for DNA packaging in bacteriophage T4. J. Mol. Biol. 1999, 289, 249–260. [Google Scholar] [CrossRef]
- Sabanayagam, C.R.; Oram, M.; Lakowicz, J.R.; Black, L.W. Viral DNA packaging studied by fluorescence correlation spectroscopy. Biophys. J. 2007, 93, L17–L19. [Google Scholar] [CrossRef]
- Driedonks, R.A.; Engel, A.; tenHeggeler, B.; van Driel, R. Gene 20 product of bacteriophage T4 its purification and structure. J. Mol. Biol. 1981, 152, 641–662. [Google Scholar] [CrossRef]
- Hendrix, R.W. Symmetry mismatch and DNA packaging in large bacteriophages. Proc. Natl. Acad. Sci. USA 1978, 75, 4779–4783. [Google Scholar] [CrossRef]
- Ishii, T.; Yanagida, M. The two dispensable structural proteins (soc and hoc) of the T4 phage capsid; their purification and properties, isolation and characterization of the defective mutants, and their binding with the defective heads in vitro. J. Mol. Biol. 1977, 109, 487–514. [Google Scholar] [CrossRef]
- Hugel, T.; Michaelis, J.; Hetherington, C.L.; Jardine, P.J.; Grimes, S.; Walter, J.M.; Falk, W.; Anderson, D.L.; Bustamante, C. Experimental test of connector rotation during DNA packaging into bacteriophage phi29 capsids. PLoS Biol. 2007, 5, e59. [Google Scholar] [CrossRef]
- Black, L.W.; Yan, B.; Ray, K. The T4 TerL Prohead Packaging Motor Does Not Drive DNA Translocation by a Proposed Dehydration Mechanism. Viruses 2020, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Dixit, A.B.; Ray, K.; Black, L.W. Compression of the DNA substrate by a viral packaging motor is supported by removal of intercalating dye during translocation. Proc. Natl. Acad. Sci. USA 2012, 109, 20419–20424. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Singh, D.; Lu, S.; Kottadiel, V.I.; Vafabakhsh, R.; Mahalingam, M.; Chemla, Y.R.; Ha, T.; Rao, V.B. A viral genome packaging ring-ATPase is a flexibly coordinated pentamer. Nature Commun. 2021, 12, 6548. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.S.; Kutter, E.; Mosig, G.; Arisaka, F.; Kunisawa, T.; Ruger, W. Bacteriophage T4 Genome. Microbiol. Mol. Biol. Rev. 2003, 67, 86–156. [Google Scholar] [CrossRef]
- Depping, R.; Lohaus, C.; Meyer, H.E.; Rüger, W. The mono-ADP-ribosyltransferases Alt and ModB of bacteriophage T4: Target proteins identified. Biochem. Biophys. Res. Commun. 2005, 335, 1217–1223. [Google Scholar] [CrossRef]
- Hinton, D.M. Transcriptional control in the prereplicative phase of T4 development. Virol. J. 2010, 7, 289. [Google Scholar] [CrossRef]
- Goff, C.G.; Setzer, J. ADP ribosylation of Escherichia coli RNA polymerase is nonessential for bacteriophage T4 development. J. Virol. 1980, 33, 547–549. [Google Scholar] [CrossRef]
- Rao, V.; Black, L. Structure and assembly of bacteriophage T4 head. Virol. J. 2010, 7, 356. [Google Scholar] [CrossRef]
- Black, L.W.; Ahmad-Zadeh, C. Internal proteins of bacteriophage T4D: Their characterization and relation to head structure and assembly. J. Mol. Biol. 1971, 57, 71–92. [Google Scholar] [CrossRef]
- Black, L.W. Bacteriophage T4 internal protein mutants: Isolation and properties. Virology 1974, 60, 166–179. [Google Scholar] [CrossRef]
- Showe, M.K.; Black, L.W. Assembly core of bacteriophage T4: An intermediate in head formation. Nature 1973, 242, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Black, L.W.; Brown, D.T. Head morphologies in bacteriophage T4 head and internal protein mutant infections. J. Virol. 1976, 17, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Black, L.W.; Abremski, K. Restriction of phage T4 internal protein I mutants by a strain of Escherichia coli. Virology 1974, 60, 180–191. [Google Scholar] [CrossRef]
- Abremski, K.; Black, B.W. The function of bacteriophage T4 internal protein I in a restrictive strain of Escherichia coli. Virology 1979, 97, 439–447. [Google Scholar] [CrossRef]
- Bair, C.L.; Black, L.W. A Type IV Modification Dependent Restriction Nuclease that Targets Glucosylated Hydroxymethyl Cytosine Modified DNAs. J. Mol. Biol. 2007, 366, 768–778. [Google Scholar] [CrossRef]
- Bair, C.L.; Rifat, D.; Black, L.W. Exclusion of glucosyl-hydroxymethylcytosine DNA containing bacteriophages is overcome by the injected protein inhibitor IPI*. J. Mol. Biol. 2007, 366, 779–789. [Google Scholar] [CrossRef]
- Rifat, D.; Wright, N.T.; Varney, K.M.; Weber, D.J.; Black, L.W. Restriction endonuclease inhibitor IPI* of bacteriophage T4: A novel structure for a dedicated target. J. Mol. Biol. 2008, 375, 720–734. [Google Scholar] [CrossRef]
- Isobe, T.; Black, L.W.; Tsugita, A. Protein cleavage during virus assembly: A novel specificity of assembly dependent cleavage in bacteriophage T4. Proc. Natl. Acad. Sci. USA 1976, 73, 4205–4209. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Keller, B.; Bickle, T.A. The nucleotide sequence of gene 21 of bacteriophage T4 coding for the prohead protease. Gene 1986, 49, 245–251. [Google Scholar] [CrossRef]
- Cheng, H.; Shen, N.; Pei, J.; Grishin, N.V. Double-stranded DNA bacteriophage prohead protease is homologous to herpesvirus protease. Protein Sci. 2004, 13, 2260–2269. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Mushegian, A. Displacements of prohead protease genes in the late operons of double-stranded-DNA bacteriophages. J. Bacteriol. 2004, 186, 4369–4375. [Google Scholar] [CrossRef] [PubMed]
- Fokine, A.; Rossmann, M.G. Common evolutionary origin of procapsid proteases, phage tail tubes, and tubes of bacterial Type VI secretion systems. Structure 2016, 24, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Onorato, L.; Showe, M.K. Gene 21 protein-dependent proteolysis in vitro of purified gene 22 product of bacteriophage T4. J. Mol. Biol. 1975, 92, 395–412. [Google Scholar] [CrossRef]
- Keller, B.; Sengstag, C.; Kellenberger, E.; Bickle, T.A. Gene 68, a new bacteriophage T4 gene which codes for the 17K prohead core protein is involved in head size determination. J. Mol. Biol. 1984, 179, 415–430. [Google Scholar] [CrossRef]
- Carrascosa, J.L. Head maturation pathway of bacteriophages T4 and T2. IV. In vitro transformation of T4 head-related particles produced by mutants in gene 17 to capsid-like structures. J. Virol. 1978, 26, 420–428. [Google Scholar] [CrossRef]
- Steven, A.C.; Bauer, A.C.; Bisher, M.E.; Robey, F.A.; Black, L.W. The maturation-dependent conformational change of phage T4 capsid involves the translocation of specific epitopes between the inner and the outer capsid surfaces. J. Struct. Biol. 1991, 106, 221–236. [Google Scholar] [CrossRef]
- Steven, A.C.; Couture, E.; Aebi, U.; Showe, M.K. Structure of T4 polyheads: II. A pathway of polyhead transformations as a model for T4 capsid maturation. J. Mol. Biol. 1976, 106, 187–221. [Google Scholar] [CrossRef]
- Steven, A.C.; Greenstone, H.L.; Booy, F.P.; Black, L.W.; Ross, P.D. Conformational changes of a viral capsid protein. Thermodynamic rationale for proteolytic regulation of bacteriophage T4 capsid expansion, co-operativity, and super-stabilization by soc binding. J. Mol. Biol. 1992, 228, 870–884. [Google Scholar] [CrossRef]
- Hong, Y.-R.; Black, L.W. Protein folding studies in vivo with a bacteriophage T4 expression-packaging-processing vector that delivers encapsidated fusion proteins into bacteria. Virology 1993, 194, 481–490. [Google Scholar] [CrossRef]
- Mullaney, J.M.; Black, L.W. Capsid targeting sequence targets foreign proteins into bacteriophage T4 and permits proteolytic processing. J. Mol. Biol. 1996, 261, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Desmond, M.I.; Mallory, S.A.; Benitez, A.D.; Buckley, L.J.; Weintraub, S.T.; Osier, M.V.; Black, L.W.; Thomas, J.A. To Be or Not To Be T4: Evidence of a Complex Evolutionary Pathway of Head Structure and Assembly in Giant Salmonella Virus SPN3US. Front. Microbiol. 2017, 8, 2251. [Google Scholar] [CrossRef] [PubMed]
- Hintermann, E.; Kuhn, A. Bacteriophage T4 gene 21 encodes two proteins essential for phage maturation. Virology 1992, 189, 474–482. [Google Scholar] [CrossRef]
- Mullaney, J.M.; Black, L.W. Activity of foreign proteins targeted within the bacteriophage T4 head and prohead: Implications for packaged DNA structure. J. Mol. Biol. 1998, 283, 913–929. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- Ren, Z.J.; Lewis, G.K.; Wingfield, P.T.; Locke, E.G.; Steven, A.C.; Black, L.W. Phage display of intact domains at high copy number: A system based on SOC, the small outer capsid protein of bacteriophage T4. Protein Sci. 1996, 5, 1833–1843. [Google Scholar] [CrossRef]
- Malys, N.; Chang, D.Y.; Baumann, R.G.; Xie, D.; Black, L.W. A bipartite bacteriophage T4 SOC and HOC randomized peptide display library: Detection and analysis of phage T4 terminase (gp17) and late sigma factor (gp55) interaction. J. Mol. Biol. 2002, 319, 289–304. [Google Scholar] [CrossRef]
- Geiduschek, E.P.; Kassavetis, G.A. Transcription of the T4 late genes. Virol. J. 2010, 7, 288. [Google Scholar] [CrossRef]
- Russell, R.L. Speciation among the T-Even Bacteriophages; California Institute of Techonology: Pasadena, CA, USA, 1967. [Google Scholar]
- Liu, L.; Cai, L.; Zhang, R. Co-existence of freshwater and marine T4-like myoviruses in a typical subtropical estuary. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Filée, J.; Tétart, F.; Suttle, C.A.; Krisch, H.M. Marine T4-type bacteriophages, a ubiquitous component of the dark matter of the biosphere. Proc. Natl. Acad. Sci. USA 2005, 102, 12471–12476. [Google Scholar] [CrossRef]
- Bellas, C.M.; Anesio, A.M. High diversity and potential origins of T4-type bacteriophages on the surface of Arctic glaciers. Extremophiles 2013, 17, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Chibani-Chennoufi, S.; Sidoti, J.; Bruttin, A.; Dillmann, M.-L.; Kutter, E.; Qadri, F.; Sarker, S.A.; Brüssow, H. Isolation of Escherichia coli bacteriophages from the stool of pediatric diarrhea patients in Bangladesh. J. Bacteriol. 2004, 186, 8287–8294. [Google Scholar] [CrossRef] [PubMed]
- Hallewell, J.; Niu, Y.D.; Munns, K.; McAllister, T.A.; Johnson, R.P.; Ackermann, H.W.; Thomas, J.E.; Stanford, K. Differing populations of endemic bacteriophages in cattle shedding high and low numbers of Escherichia coli O157:H7 bacteria in feces. Appl. Environ. Micro. 2014, 80, 3819–3825. [Google Scholar] [CrossRef] [PubMed]
- Goldin, S.; Hulata, Y.; Baran, N.; Lindell, D. Quantification of T4-Like and T7-Like Cyanophages Using the Polony Method Show They Are Significant Members of the Virioplankton in the North Pacific Subtropical Gyre. Front. Microbiol. 2020, 11, 1210. [Google Scholar] [CrossRef]
- Petrov, V.; Ratnayaka, S.; Nolan, J.; Miller, E.; Karam, J. Genomes of the T4-related bacteriophages as windows on microbial genome evolution. Virol. J. 2010, 7, 292. [Google Scholar] [CrossRef]
- Repoila, F.; Tetart, F.; Bouet, J.-Y.; Krisch, H.M. Genomic polymorpism in the T-even phages. EMBO J. 1994, 13, 4181–4192. [Google Scholar] [CrossRef]
- Lamm, N.; Wang, Y.; Mathews, C.K.; Ruger, W. Deoxycytidylate hydroxymethylase gene of bacteriophage T4. Nucleotide sequence determination and over-expression of the gene. Eur. J. Biochem. 1988, 172, 553–563. [Google Scholar] [CrossRef]
- Greenberg, G.R.; He, P.; Hilfinger, J.; Tseng, M.-J. Deoxyribonucleoside triphosphate synthesis and phage T4 DNA replication. In Molecular Biology of Bacteriophage T4; Karam, J.D., Ed.; ASM Press: Washington, DC, USA, 1994; pp. 14–27. [Google Scholar]
- Weigele, P.; Raleigh, E.A. Biosynthesis and Function of Modified Bases in Bacteria and Their Viruses. Chem. Rev. 2016, 116, 12655–12687. [Google Scholar] [CrossRef]
- Lehman, I.R.; Pratt, E.A. On the structure of the glucosylated hydroxymethylcytosine nucleotides of coliphages T2, T4, and T6. J. Biol. Chem. 1960, 235, 3254–3259. [Google Scholar] [CrossRef]
- Carlson, K.; Raleigh, E.A.; Hattman, S. Restriction and Modification. In Molecular Biology of Bacteriophage T4; Karam, J.D., Ed.; ASM Press: Washington, DC, USA, 1994; pp. 369–381. [Google Scholar]
- Thomas, J.A.; Orwenyo, J.; Wang, L.-X.; Black, L.W. The Odd “RB” Phage-Identification of Arabinosylation as a New Epigenetic Modification of DNA in T4-Like Phage RB69. Viruses 2018, 10, 313. [Google Scholar] [CrossRef]
- Krylov, V.N.; Zhazykov, I. Pseudomonas bacteriophage phiKZ—Possible model for studying the genetic control of morphogenesis. Genetika 1978, 14, 678–685. [Google Scholar] [PubMed]
- Krylov, V.; Bourkaltseva, M.; Pleteneva, E.; Shaburova, O.; Krylov, S.; Karaulov, A.; Zhavoronok, S.; Svitich, O.; Zverev, V. Phage phiKZ—The First of Giants. Viruses 2021, 13, 149. [Google Scholar] [CrossRef] [PubMed]
- Krylov, V.N.; Smirnova, T.A.; Minenkova, I.B.; Plotnilova, T.G.; Zhazikov, I.Z.; Khrenova, E.A. Pseudomonas bacteriophage PhiKZ contains an inner body in its capsid. Can. J. Microbiol. 1984, 30, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Mesyanzhinov, V.V.; Robben, J.; Grymonprez, B.; Kostyuchenko, V.A.; Bourkaltseva, M.V.; Sykilinda, N.N.; Krylov, V.N.; Volckaert, G. The genome of bacteriophage PhiKZ of Pseudomonas aeruginosa. J. Mol. Biol. 2002, 317, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Fokine, A.; Kostyuchenko, V.A.; Efimov, A.V.; Kurochkina, L.P.; Sykilinda, N.N.; Robben, J.; Volckaert, G.; Hoenger, A.; Chipman, P.R.; Battisti, A.J.; et al. A three-dimensional cryo-electron microscopy structure of the bacteriophage phiKZ head. J. Mol. Biol. 2005, 352, 117–124. [Google Scholar] [CrossRef]
- Lecoutere, E.; Ceyssens, P.-J.; Miroshnikov, K.A.; Mesyanzhinov, V.V.; Krylov, V.N.; Noben, J.-P.; Robben, J.; Hertveldt, K.; Volckaert, G.; Lavigne, R. Identification and comparative analysis of the structural proteomes of PhiKZ and EL, two giant Pseudomonas aeruginosa bacteriophages. Proteomics 2009, 9, 3215–3219. [Google Scholar] [CrossRef]
- Serwer, P.; Hayes, S.J.; Zaman, S.; Lieman, K.; Rolando, M.; Hardies, S.C. Improved isolation of undersampled bacteriophages: Finding of distant terminase genes. Virology 2004, 329, 412–424. [Google Scholar] [CrossRef]
- Thomas, J.A.; Rolando, M.R.; Carroll, C.A.; Shen, P.S.; Belnap, D.M.; Weintraub, S.T.; Serwer, P.; Hardies, S.C. Characterization of Pseudomonas chlororaphis myovirus 201φ2-1 via genomic sequencing, mass spectrometry, and electron microscopy. Virology 2008, 376, 330–338. [Google Scholar] [CrossRef]
- Sokolova, O.S.; Shaburova, O.V.; Pechnikova, E.V.; Shaytan, A.K.; Krylov, S.V.; Kiselev, N.A.; Krylov, V.N. Genome packaging in EL and Lin68, two giant PhiKZ-like bacteriophages of P. aeruginosa. Virology 2014, 468–470, 472–478. [Google Scholar] [CrossRef]
- Thomas, J.A.; Weintraub, S.T.; Hakala, K.; Serwer, P.; Hardies, S.C. Proteome of the large Pseudomonas myovirus 201φ2-1: Delineation of proteolytically processed virion proteins. Mol. Cell. Proteom. 2010, 9, 940–951. [Google Scholar] [CrossRef]
- Ceyssens, P.-J.; Minakhin, L.; Van den Bossche, A.; Yakunina, M.; Klimuk, E.; Blasdel, B.; De Smet, J.; Noben, J.-P.; Bläsi, U.; Severinov, K.; et al. Development of giant bacteriophage ϕKZ is independent of the host transcription apparatus. J. Virol. 2014, 88, 10501–10510. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Thomas, J.A.; Cheng, N.; Black, L.W.; Steven, A.C. Bubblegrams reveal the inner body structure of φKZ. Science 2012, 335, 182. [Google Scholar] [CrossRef] [PubMed]
- Schaerli, C.; Kellenberger. Head maturation pathway of bacteriophages T4 and T2. V. Maturable epsilon-particle accumulating an acridine-treated bacteriophage T4-infected cells. J. Virol. 1980, 33, 830–844. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A.; Weintraub, S.T.; Wu, W.; Winkler, D.C.; Cheng, N.; Steven, A.C.; Black, L.W. Extensive proteolysis of head and inner body proteins by a morphogenetic protease in the giant Pseudomonas aeruginosa phage φKZ. Mol. Microbiol. 2012, 84, 324–339. [Google Scholar] [CrossRef]
- Thomas, J.A.; Black, L.W. Mutational analysis of the Pseudomonas aeruginosa myovirus φKZ morphogenetic protease gp175. J. Virol. 2013, 87, 8713–8725. [Google Scholar] [CrossRef]
- Lee, J.-H.; Shin, H.; Kim, H.; Ryu, S. Complete genome sequence of Salmonella bacteriophage SPN3US. J. Virol. 2011, 85, 13470–13471. [Google Scholar] [CrossRef]
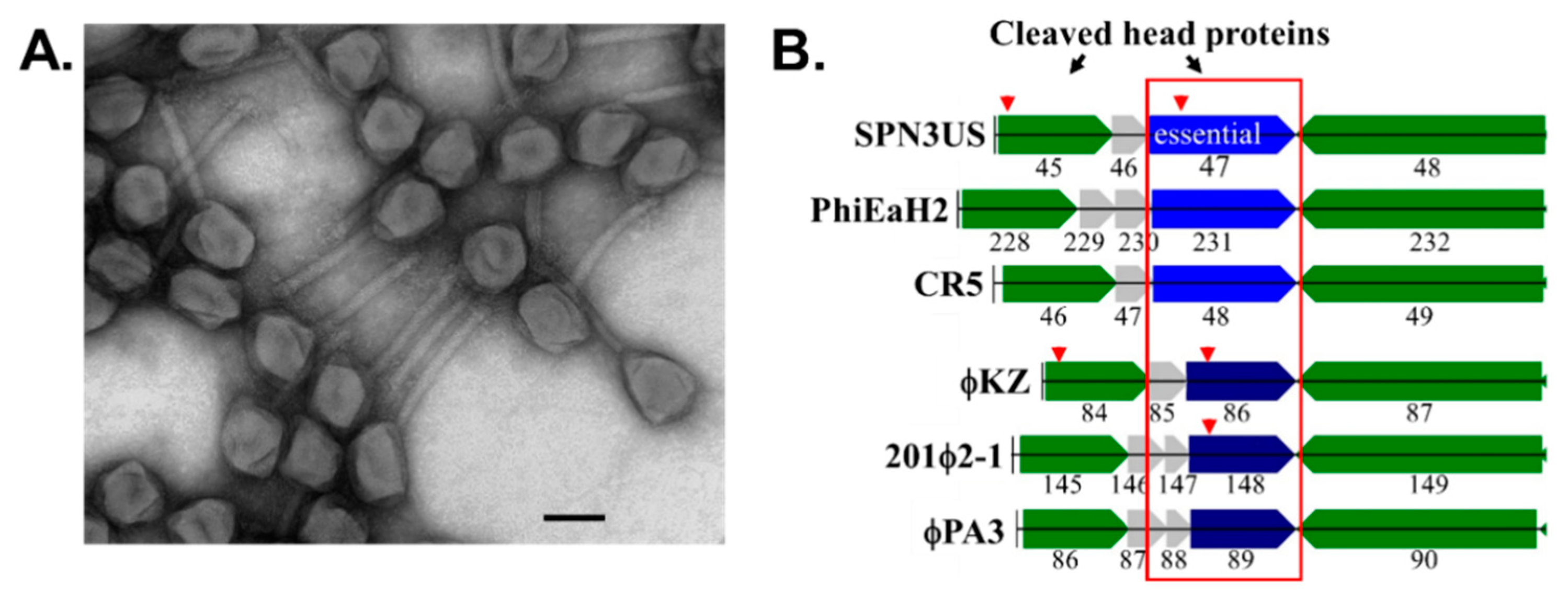
- Reilly, E.R.; Abajorga, M.K.; Kiser, C.; Mohd Redzuan, N.H.; Haidar, Z.; Adams, L.E.; Diaz, R.; Pinzon, J.A.; Hudson, A.O.; Black, L.W.; et al. A Cut above the Rest: Characterization of the Assembly of a Large Viral Icosahedral Capsid. Viruses 2020, 12, 725. [Google Scholar] [CrossRef]
- Heymann, J.B.; Wang, B.; Newcomb, W.W.; Wu, W.; Winkler, D.C.; Cheng, N.; Reilly, E.R.; Hsia, R.C.; Thomas, J.A.; Steven, A.C. The Mottled Capsid of the Salmonella Giant Phage SPN3US, a Likely Maturation Intermediate with a Novel Internal Shell. Viruses 2020, 12, 910. [Google Scholar] [CrossRef]
- Tessman, I. Mutagenic treatment of double- and single-stranded DNA phages T4 and S13 with hydroxylamine. Virology 1968, 35, 330–333. [Google Scholar] [CrossRef]
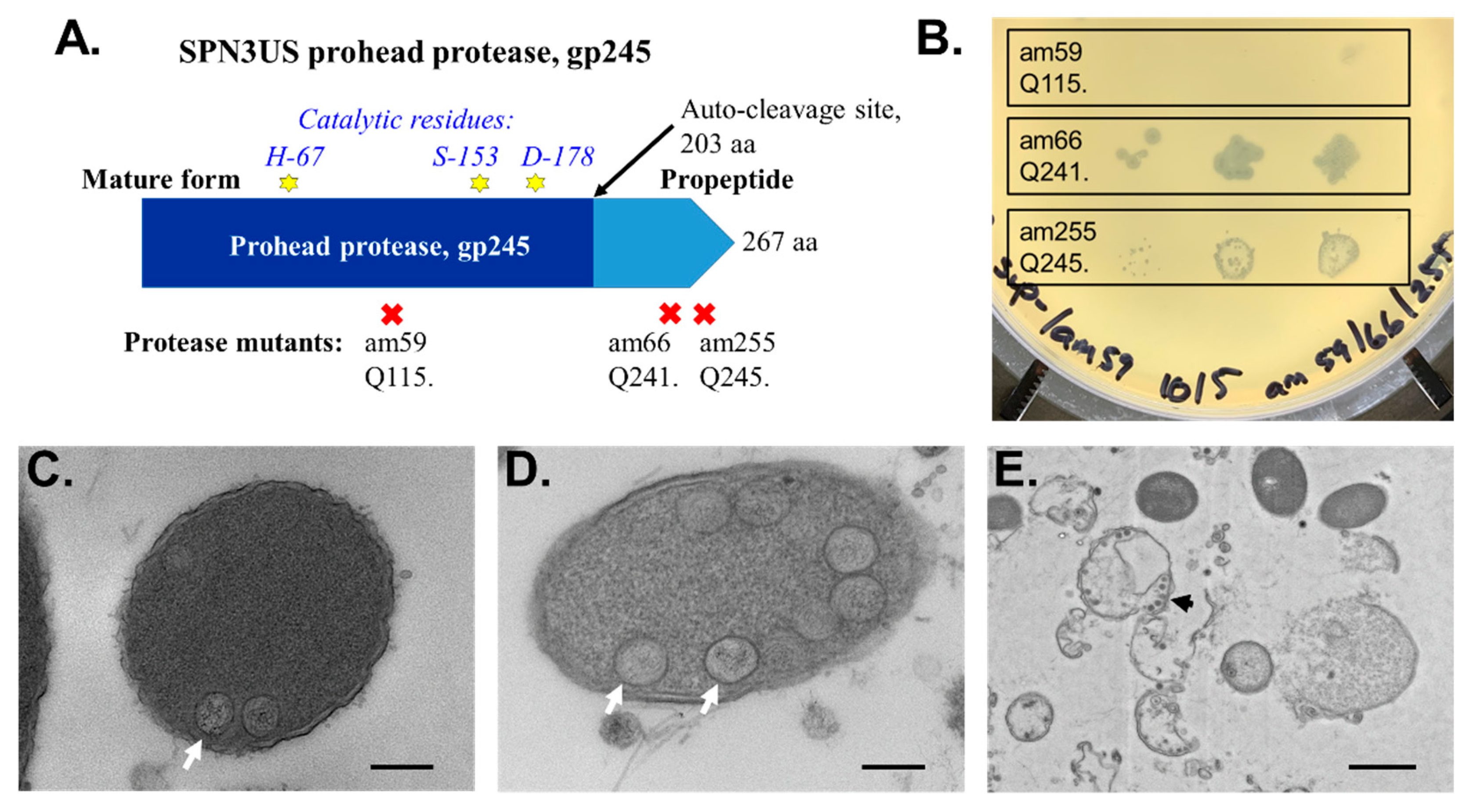
- Thomas, J.A.; Benítez Quintana, A.D.; Bosch, M.A.; Coll De Peña, A.; Aguilera, E.; Coulibaly, A.; Wu, W.; Osier, M.V.; Hudson, A.O.; Weintraub, S.T.; et al. Identification of essential genes in the Salmonella phage SPN3US reveals novel insights into giant phage head structure and assembly. J. Virol. 2016, 90, 10284–10298. [Google Scholar] [CrossRef]
- Weintraub, S.T.; Redzuan, N.H.M.; Barton, M.K.; Md Amin, N.A.; Desmond, M.I.; Adams, L.E.; Ali, B.; Pardo, S.; Molleur, D.; Wu, W.; et al. Global proteomic profiling of Salmonella infection by a giant phage. J. Virol. 2019, 93, e01833-18. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| gp | Function | Prohead, Copy Number | Mature Head, Copy Number | Cleaved? | Essential? |
|---|---|---|---|---|---|
| Outer shell | |||||
| 20 | portal | 12 | 12 | - | + |
| 23 | major capsid | 930 | 930 | + | + |
| 24 | head vertex | 55 | 55 | + | + |
| Hoc | decoration protein | 0 | 155 | - | - |
| Soc | decoration protein | 0 | 810 | - | - |
| Internal | |||||
| 21 | prohead protease | 72 | 3 | + | + |
| 22 | scaffold | 576 | 115 (2.5 kDa fragment) | + | + |
| 67 | core component | 341 | 139 (3.9 kDa fragment) | + | + |
| 68 | core component | 240 | 0 | - | + |
| Alt | ADP-ribosyltransferase | 40 | 40 | + | - |
| IPI | internal protein I, Type IV restriction enzyme inhibitor | 360 | 360 | + | semi |
| IPII | internal protein II | 360 | 360 | + | - |
| IPIII | internal protein III | 370 | 370 | + | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuhn, A.; Thomas, J.A. The Beauty of Bacteriophage T4 Research: Lindsay W. Black and the T4 Head Assembly. Viruses 2022, 14, 700. https://doi.org/10.3390/v14040700
Kuhn A, Thomas JA. The Beauty of Bacteriophage T4 Research: Lindsay W. Black and the T4 Head Assembly. Viruses. 2022; 14(4):700. https://doi.org/10.3390/v14040700
Chicago/Turabian StyleKuhn, Andreas, and Julie A. Thomas. 2022. "The Beauty of Bacteriophage T4 Research: Lindsay W. Black and the T4 Head Assembly" Viruses 14, no. 4: 700. https://doi.org/10.3390/v14040700
APA StyleKuhn, A., & Thomas, J. A. (2022). The Beauty of Bacteriophage T4 Research: Lindsay W. Black and the T4 Head Assembly. Viruses, 14(4), 700. https://doi.org/10.3390/v14040700