
Antibody Evaluation and Mutations of Antigenic Epitopes in the Spike Protein of the Porcine Epidemic Diarrhea Virus from Pig Farms with Repeated Intentional Exposure (Feedback)

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pig Farms and Sample Collection
2.2. Cells and Virus
2.3. Serology
2.4. RNA Extraction and RT-PCR
2.5. Sequencing of S Gene and Phylogenetic Analysis
2.6. Data Analysis and Statistics
3. Results
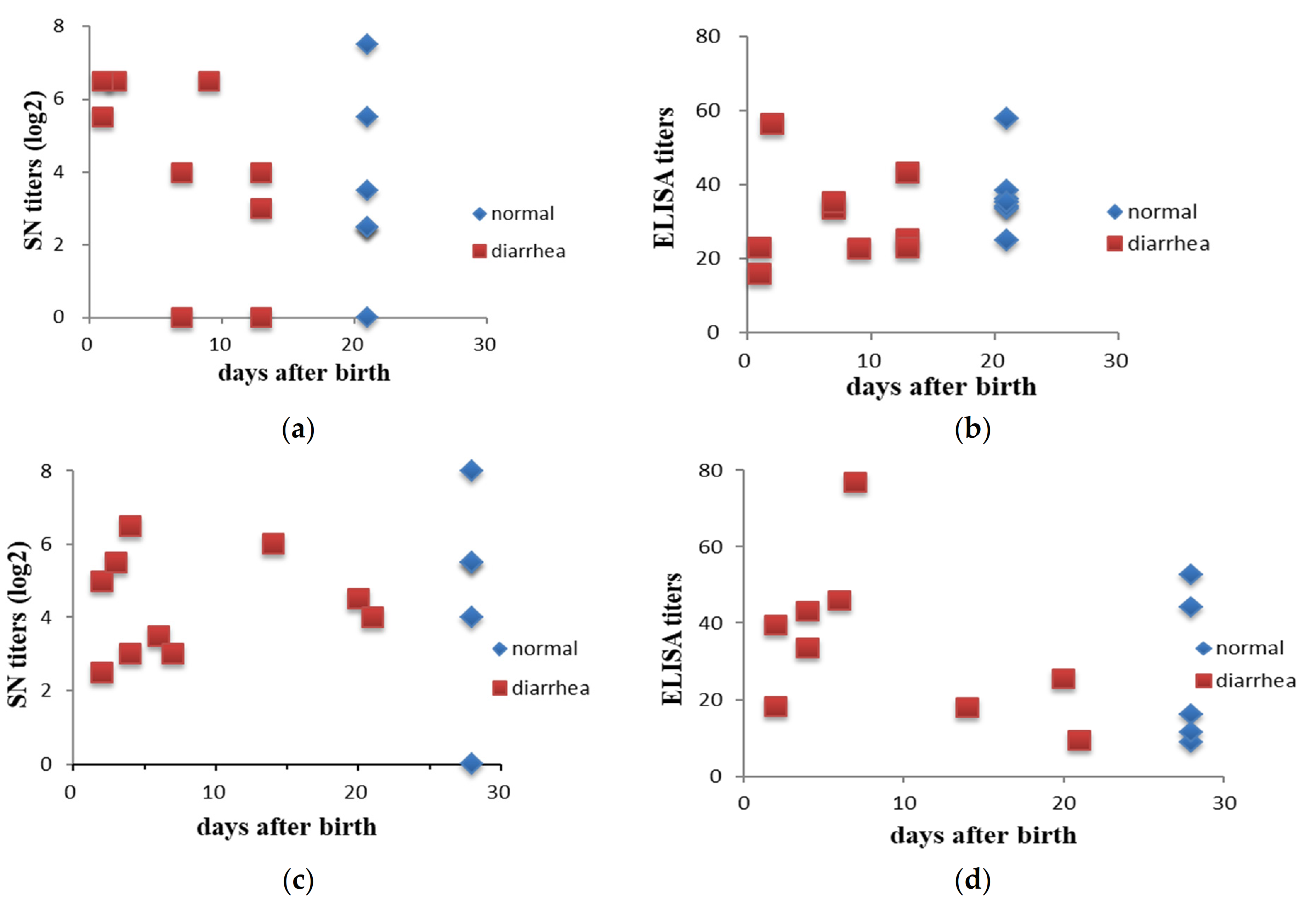
3.1. Lactogenic Immunity Increased after Intentional Exposure (Feedback)
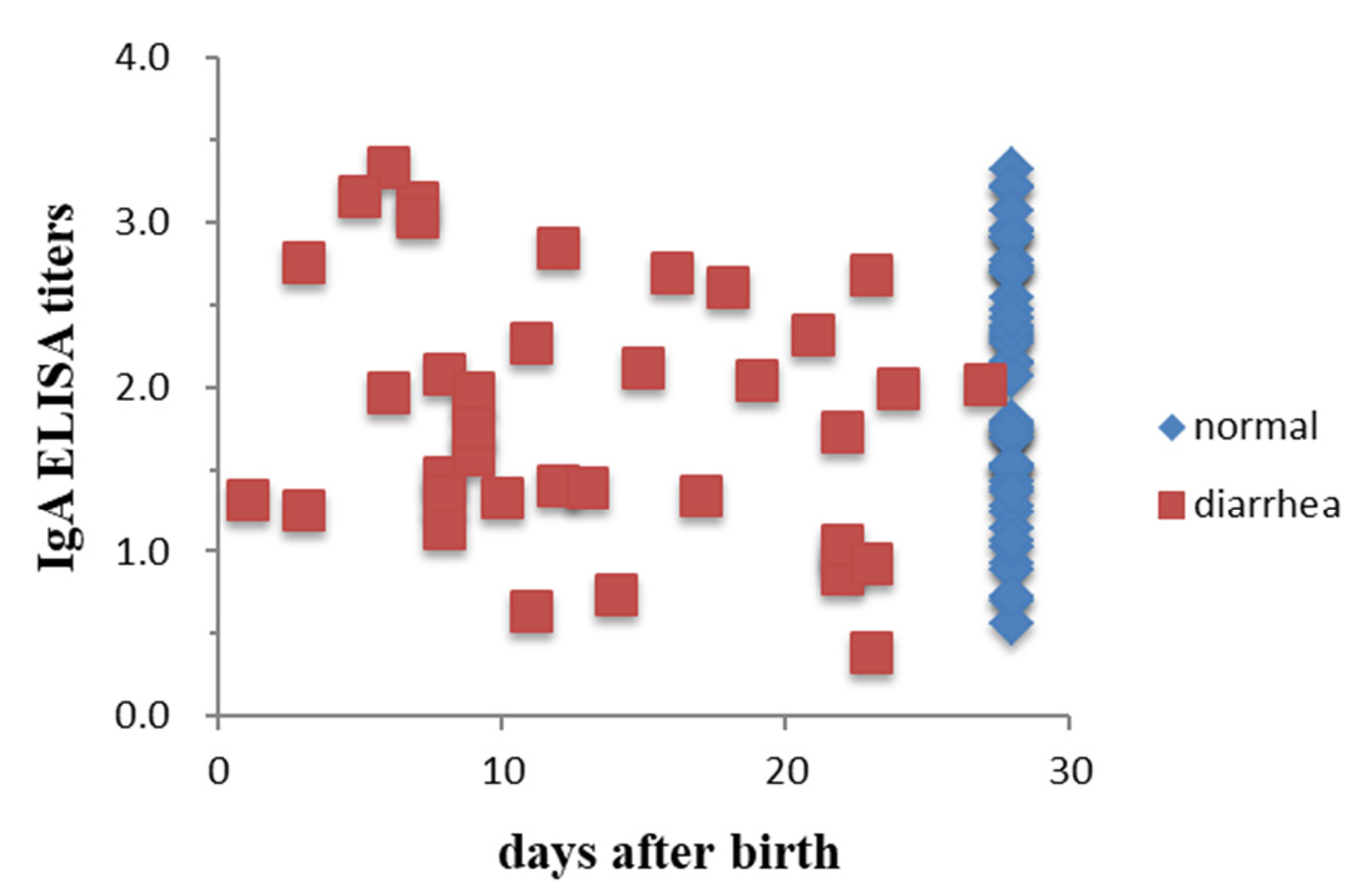
3.2. Lactogenic Immunity Showed No Association with the Protection of Piglets from PEDV in the Fields
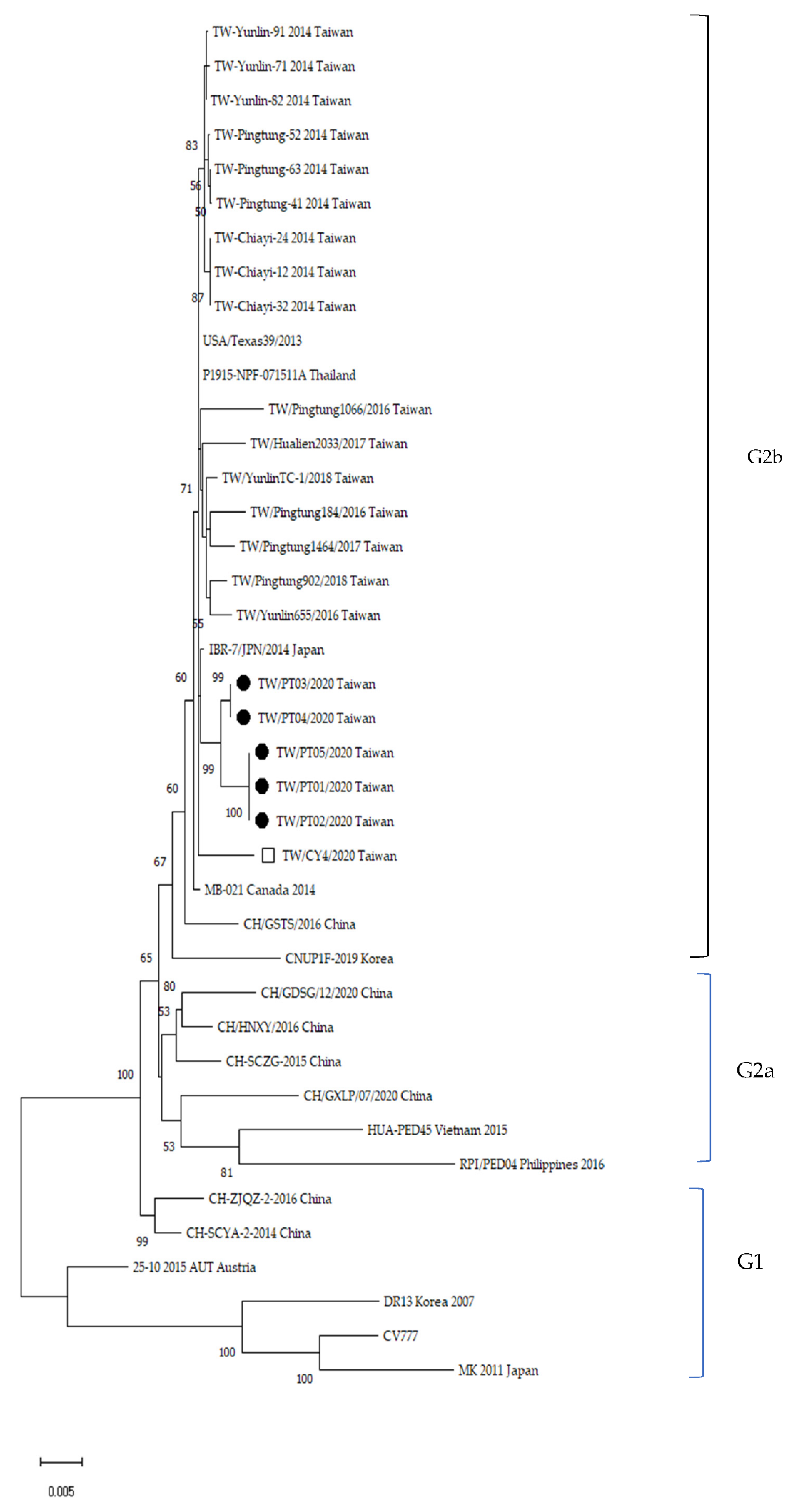
3.3. Phylogenetic and Genetic Analysis
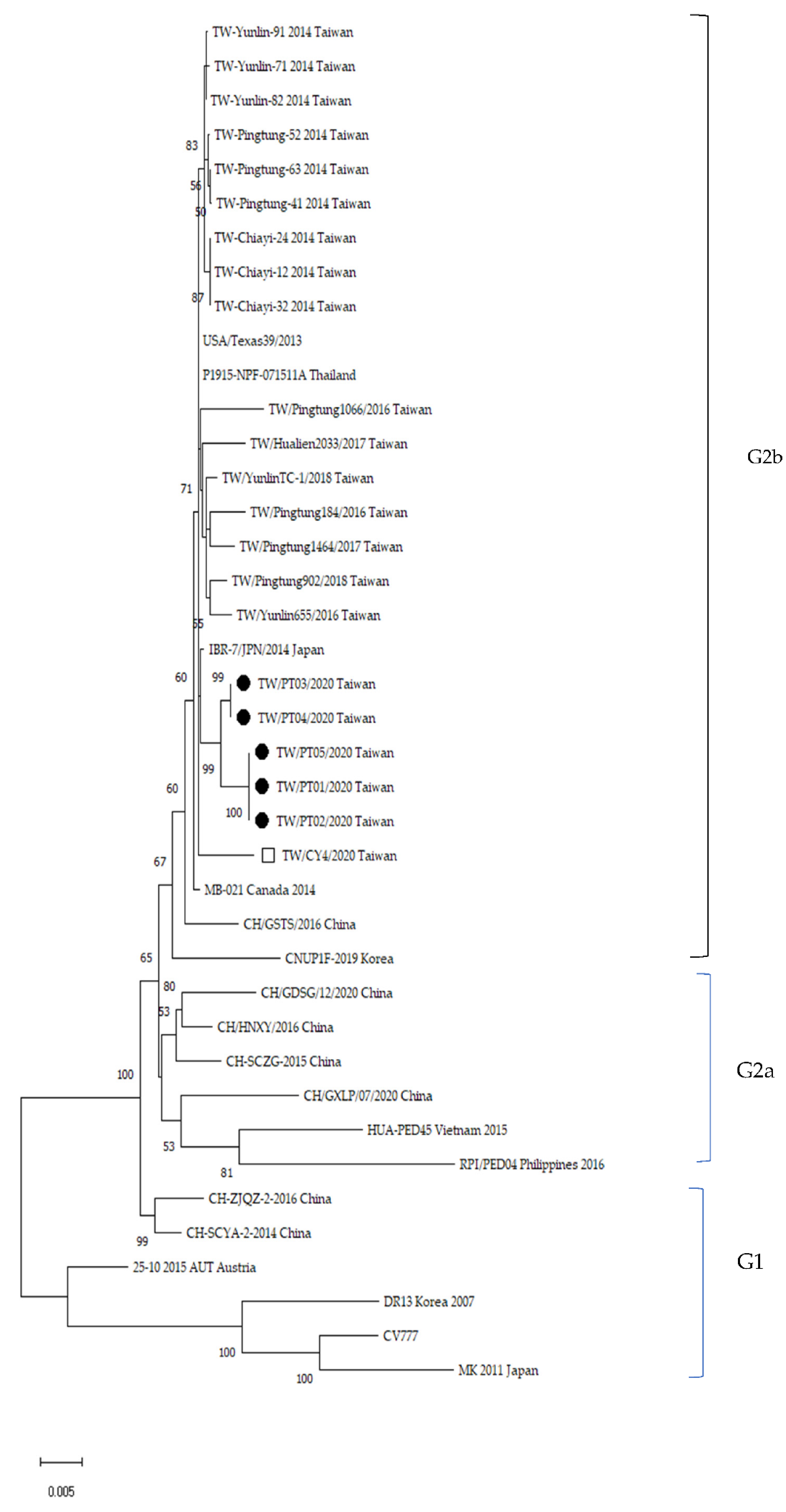
3.4. Alignment of Antigenic Epitopes of S Protein
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Number | Virus Strain Name | GenBank Accession No. | Year | Country |
|---|---|---|---|---|
| 1 | TW/PT01/2020 | OM681613 | 2020 | Taiwan |
| 2 | TW/PT02/2020 | OM863563 | 2020 | Taiwan |
| 3 | TW/PT03/2020 | OM681614 | 2020 | Taiwan |
| 4 | TW/PT04/2020 | OM863564 | 2020 | Taiwan |
| 5 | TW/PT05/2020 | OM863565 | 2020 | Taiwan |
| 6 | TW/CY4/2020 | OM863562 | 2020 | Taiwan |
| 7 | TW-Yunlin-91 | KP276248.1 | 2014 | Taiwan |
| 8 | TW-Yunlin-71 | KP276249.1 | 2014 | Taiwan |
| 9 | TW-Yunlin-82 | KP276247.1 | 2014 | Taiwan |
| 10 | TW-Pingtung-52 | KP276252.1 | 2014 | Taiwan |
| 11 | TW-Pingtung-63 | KP276250.1 | 2014 | Taiwan |
| 12 | TW-Pingtung-41 | KP276251.1 | 2014 | Taiwan |
| 13 | TW-Chiay-24 | KP276244.1 | 2014 | Taiwan |
| 14 | TW-Chiay-12 | KP276245.1 | 2014 | Taiwan |
| 15 | TW-Chiay-32 | KP276246.1 | 2014 | Taiwan |
| 16 | USA/Texas39/2013 | KJ645645.1 | 2013 | USA |
| 17 | P1915-NPF-071511A | KX981900.1 | 2015 | Thailand |
| 18 | TW/Pingtung1066/2016 | MK673518.1 | 2016 | Taiwan |
| 19 | TW/Hualien2033/2017 | MK673517.1 | 2017 | Taiwan |
| 20 | TW/YunlinTC-1/2018 | MK673544.1 | 2018 | Taiwan |
| 21 | TW/Pingtung184/2016 | MK673523.1 | 2016 | Taiwan |
| 22 | TW/Pingtung1464/2017 | MK673521.1 | 2017 | Taiwan |
| 23 | TW/Pingtung902/2018 | MK673534.1 | 2018 | Taiwan |
| 24 | TW/Yunlin655/2016 | MK673543.1 | 2016 | Taiwan |
| 25 | IBR-7/JPN/2014 | LC063832.1 | 2014 | Japan |
| 26 | MB-021 | KM196111.1 | 2014 | Canada |
| 27 | CH/GSTS/2016 | MF152598.1 | 2016 | China |
| 28 | CNUP1F-2019 | MN725088.1 | 2019 | Korea |
| 29 | CH/GDSG/12/2010 | MZ161002.1 | 2010 | China |
| 30 | CH/HNXY/2016 | MF152599.1 | 2016 | China |
| 31 | CH-SCZG-2015 | KU975422.1 | 2015 | China |
| 32 | CH/GXLP/07/2020 | MZ161060.1 | 2020 | China |
| 33 | HUA-PED45 | KP455313.1 | 2015 | Vietnam |
| 34 | RPI/PED04 | KT357508.1 | 2016 | Philippines |
| 35 | CH-ZJQZ-2-2016 | MG020555.1 | 2016 | China |
| 36 | CH-SCYA-2-2014 | KU975423.1 | 2014 | China |
| 37 | 25-10-2015-AUT | KT206204.1 | 2015 | Austria |
| 38 | DR 13 | DQ862099.1 | 2007 | Korea |
| 39 | CV777 | AF353511.1 | 2001 | Belgium |
| 40 | MK | AB548624.1 | 2011 | Japan |
References
- Diel, D.G.; Lawson, S.; Okda, F.; Singrey, A.; Clement, T.; Fernandes, M.H.V.; Christopher-Hennings, J.; Nelson, E.A. Porcine epidemic diarrhea virus: An overview of current virological and serological diagnostic methods. Virus Res. 2016, 226, 60–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, C.; Xiao, Y.; Li, X.; Tian, K. Porcine epidemic diarrhea virus: Current insights. Virus Adapt. Treat. 2016, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Weng, L.; Weersink, A.; Poljak, Z.; de Lange, K.; von Massow, M. An economic evaluation of intervention strategies for Porcine Epidemic Diarrhea (PED). Prev. Vet. Med. 2016, 134, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Kirchdoerfer, R. Structure and Immune Recognition of the Porcine Epidemic Diarrhea Virus Spike Protein. Microsc. Microanal. 2020, 26, 1–2. [Google Scholar] [CrossRef]
- Liu, C.; Tang, J.; Ma, Y.; Liang, X.; Yang, Y.; Peng, G.; Qi, Q.; Jiang, S.; Li, J.; Du, L.; et al. Receptor Usage and Cell Entry of Porcine Epidemic Diarrhea Coronavirus. J. Virol. 2015, 89, 6121–6125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhang, Z.; Li, J.; Gao, Y.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; Yang, H. Porcine epidemic diarrhea virus S1 protein is the critical inducer of apoptosis. Virol. J. 2018, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Ma, J.; Wang, Y.; Wang, M.; Song, W.; Zhang, W.; Lu, C.; Yao, H. Genomic and epidemiological characteristics provide new insights into the phylogeographical and spatiotemporal spread of porcine epidemic diarrhea virus in Asia. J. Clin. Microbiol. 2015, 53, 1484–1492. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; van Kuppeveld, F.J.M.; He, Q.; Rottier, P.J.M.; Bosch, B.J. Cellular entry of the porcine epidemic diarrhea virus. Virus Res. 2016, 226, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Tuanthap, S.; Vongpunsawad, S.; Phupolphan, C.; Duang-In, A.; Wattanaphansak, S.; Assavacheep, P.; Theamboonlers, A.; Luengyosluechakul, S.; Amonsin, A.; Poovorawan, Y. Analysis of the spike, ORF3, and nucleocapsid genes of porcine epidemic diarrhea virus circulating on Thai swine farms, 2011–2016. PeerJ 2019, 2019, 2011–2016. [Google Scholar] [CrossRef]
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virol. J. 2015, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.M.; Saif, L.J.; Marthaler, D.; Wang, Q. Evolution, antigenicity and pathogenicity of global porcine epidemic diarrhea virus strains. Virus Res. 2016, 226, 20–39. [Google Scholar] [CrossRef] [Green Version]
- Niederwerder, M.C.; Hesse, R.A. Swine enteric coronavirus disease: A review of 4 years with porcine epidemic diarrhoea virus and porcine deltacoronavirus in the United States and Canada. Transbound. Emerg. Dis. 2018, 65, 660–675. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.N.; Chung, W.B.; Chang, S.W.; Wen, C.C.; Liu, H.; Chien, C.H.; Chiou, M.T. US-like strain of porcine epidemic diarrhea virus outbreaks in Taiwan, 2013–2014. J. Vet. Med. Sci. 2014, 76, 1297–1299. [Google Scholar] [CrossRef] [Green Version]
- Langel, S.N.; Wang, Q.; Vlasova, A.N.; Saif, L.J. Host factors affecting generation of immunity against porcine epidemic diarrhea virus in pregnant and lactating swine and passive protection of neonates. Pathogens 2020, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Hedegaard, C.J.; Heegaard, P.M.H. Passive immunisation, an old idea revisited: Basic principles and application to modern animal production systems. Vet. Immunol. Immunopathol. 2016, 174, 50–63. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, K.J.; Henry, S.; Tokach, T.; Potter, M.; Davidson, D.; Egnor, C. Infective Material, Concepts and Procedures for Intentional Sow Herd Exposure to Porcine Epidemic Diarrhea Virus; Iowa State University: Ames, IA, USA, 2013; p. 8. [Google Scholar]
- Tsai, K.J.; Deng, M.C.; Wang, F.I.; Tsai, S.H.; Chang, C.; Chang, C.Y.; Huang, Y.L. Deletion in the S1 Region of Porcine Epidemic Diarrhea Virus Reduces the Virulence and Influences the Virus-Neutralizing Activity of the Antibody Induced. Viruses 2020, 12, 1378. [Google Scholar] [CrossRef]
- Kim, Y.K.; Cho, Y.Y.; An, B.H.; Lim, S.I.; Lim, J.A.; Cho, I.S.; Le, V.P.; An, D.J. Molecular characterization of the spike and ORF3 genes of porcine epidemic diarrhea virus in the Philippines. Arch. Virol. 2016, 161, 1323–1328. [Google Scholar] [CrossRef]
- Chiou, H.Y.; Huang, Y.L.; Deng, M.C.; Chang, C.Y.; Jeng, C.R.; Tsai, P.S.; Yang, C.; Pang, V.F.; Chang, H.W. Phylogenetic Analysis of the Spike (S) Gene of the New Variants of Porcine Epidemic Diarrhoea Virus in Taiwan. Transbound. Emerg. Dis. 2017, 64, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.H.; Bae, J.L.; Kang, T.J.; Kim, J.; Chung, G.H.; Lim, C.W.; Laude, H.; Yang, M.S.; Jang, Y.S. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol. Cells 2002, 14, 295–299. [Google Scholar]
- Okda, F.A.; Lawson, S.; Singrey, A.; Nelson, J.; Hain, K.S.; Joshi, L.R.; Christopher-Hennings, J.; Nelson, E.A.; Diel, D.G. The S2 glycoprotein subunit of porcine epidemic diarrhea virus contains immunodominant neutralizing epitopes. Virology 2017, 509, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Feng, L.; Shi, H.; Chen, J.; Cui, X.; Chen, H.; Liu, S.; Tong, Y.; Wang, Y.; Tong, G. Identification of two novel B cell epitopes on porcine epidemic diarrhea virus spike protein. Vet. Microbiol. 2008, 131, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, T.; Miyama, T.; Toyomaki, H.; Sekiguchi, S.; Sasaki, Y.; Sueyoshi, M.; Makita, K. Analysis of the effect of feedback feeding on the farm-level occurrence of porcine epidemic diarrhea in kagoshima and miyazaki prefectures, japan. J. Vet. Med. Sci. 2021, 83, 1772–1781. [Google Scholar] [CrossRef]
- Jang, G.; Park, J.; Lee, C. Successful Eradication of Porcine Epidemic Diarrhea in an Enzootically Infected Farm: A Two-Year Follow-Up Study. Pathogens 2021, 10, 830. [Google Scholar] [CrossRef] [PubMed]
- Langel, S.N.; Paim, F.C.; Lager, K.M.; Vlasova, A.N.; Saif, L.J. Lactogenic immunity and vaccines for porcine epidemic diarrhea virus (PEDV): Historical and current concepts. Virus Res. 2016, 226, 93–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Z.; Xu, Z.; Zhou, Q.; Li, W.; Wu, Y.; Du, Y.; Chen, L.; Xue, C.; Cao, Y. A heterologous ‘prime-boost’ anti-PEDV immunization for pregnant sows protects neonatal piglets through lactogenic immunity against PEDV. Lett. Appl. Microbiol. 2019, 69, 258–263. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, K.; Shyu, D.L.; Dhakal, S.; Hiremath, J.; Binjawadagi, B.; Lakshmanappa, Y.S.; Guo, R.; Ransburgh, R.; Bondra, K.M.; Gauger, P.; et al. Evaluation of humoral immune status in porcine epidemic diarrhea virus (PEDV) infected sows under field conditions. Vet. Res. 2015, 46, 140. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, T.; Song, Q.; Inskeep, M.; Stone, S.; Murtaugh, M.P. Effect of Booster Vaccination with Inactivated Porcine Epidemic Diarrhea Virus on Neutralizing Antibody Response in Mammary Secretions. Viral Immunol. 2018, 31, 62–68. [Google Scholar] [CrossRef]
- Sawattrakool, K.; Stott, C.J.; Bandalaria-Marca, R.D.; Srijangwad, A.; Palabrica, D.J.; Nilubol, D. Field trials evaluating the efficacy of porcine epidemic diarrhea vaccine, RNA (Harrisvaccine) in the Philippines. Trop. Anim. Health Prod. 2020, 52, 2743–2747. [Google Scholar] [CrossRef]
- Subramaniam, S.; Yugo, D.M.; Heffron, C.L.; Rogers, A.J.; Sooryanarain, H.; LeRoith, T.; Overend, C.; Cao, D.; Meng, X.J. Vaccination of sows with a dendritic cell-targeted porcine epidemic diarrhea virus S1 protein-based candidate vaccine reduced viral shedding but exacerbated gross pathological lesions in suckling neonatal piglets. J. Gen. Virol. 2018, 99, 230–239. [Google Scholar] [CrossRef]
- Song, Q.; Stone, S.; Drebes, D.; Greiner, L.L.; Dvorak, C.M.T.; Murtaugh, M.P. Characterization of anti-porcine epidemic diarrhea virus neutralizing activity in mammary secretions. Virus Res. 2016, 226, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Lee, K.W.; Choi, H.W.; Lee, C. Immunogenicity and protective efficacy of recombinant S1 domain of the porcine epidemic diarrhea virus spike protein. Arch. Virol. 2014, 159, 2977–2987. [Google Scholar] [CrossRef]
- Annamalai, T.; Lin, C.M.; Gao, X.; Liu, X.; Lu, Z.; Saif, L.J.; Wang, Q. Cross protective immune responses in nursing piglets infected with a US spike-insertion deletion porcine epidemic diarrhea virus strain and challenged with an original US PEDV strain. Vet. Res. 2017, 48, 1–8. [Google Scholar] [CrossRef] [Green Version]
- De Arriba, M.L.; Carvajal, A.; Pozo, J.; Rubio, P. Mucosal and systemic isotype-specific antibody responses and protection in conventional pigs exposed to virulent or attenuated porcine epidemic diarrhoea virus. Vet. Immunol. Immunopathol. 2002, 85, 85–97. [Google Scholar] [CrossRef]
- Poonsuk, K.; Giménez-Lirola, L.G.; Zhang, J.; Arruda, P.; Chen, Q.; Da Silva Carrion, L.C.; Magtoto, R.; Pineyro, P.; Sarmento, L.; Wang, C.; et al. Does circulating antibody play a role in the protection of piglets against porcine epidemic diarrhea virus? PLoS ONE 2016, 11, e0153041. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine epidemic diarrhea virus (PEDV): An update on etiology, transmission, pathogenesis, and prevention and control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef]
- Scherba, G. Evaluation of responses to both oral and parenteral immunization modalities for porcine epidemic diarrhea virus in production units. J. Swine Heal. Prod. 2016, 24, 29–35. [Google Scholar]
- Van Diep, N.; Norimine, J.; Sueyoshi, M.; Lan, N.T.; Yamaguchi, R. Novel porcine epidemic diarrhea virus (PEDV) variants with large deletions in the spike (S) ene Coexist with PEDV strains possessing an intact S gene in domestic pigs in Japan: A new disease situation. PLoS ONE 2017, 12, e0170126. [Google Scholar] [CrossRef]
- Than, V.T.; Choe, S.E.; Vu, T.T.H.; Do, T.D.; Nguyen, T.L.; Bui, T.T.N.; Mai, T.N.; Cha, R.M.; Song, D.; An, D.J.; et al. Genetic characterization of the spike gene of porcine epidemic diarrhea viruses (PEDVs) circulating in Vietnam from 2015 to 2016. Vet. Med. Sci. 2020, 6, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.H.; Li, Y.Q.; Pan, Y.Q.; Guo, Y.Y.; Guo, F.; Shi, R.Z.; Xing, L. The spike glycoprotein genes of porcine epidemic diarrhea viruses isolated in China. Vet. Res. 2021, 52, 87. [Google Scholar] [CrossRef]
- Ji, Z.; Shi, D.; Shi, H.; Wang, X.; Chen, J.; Liu, J.; Ye, D.; Jing, Z.; Liu, Q.; Fan, Q.; et al. A porcine epidemic diarrhea virus strain with distinct characteristics of four amino acid insertion in the COE region of spike protein. Vet. Microbiol. 2021, 253, 108955. [Google Scholar] [CrossRef]
- Wanitchang, A.; Saenboonrueng, J.; Kaewborisuth, C.; Srisutthisamphan, K.; Jongkaewwattana, A. A single V672F substitution in the spike protein of field-isolated PEDV Promotes Cell–Cell fusion and replication in Vero E6 cells. Viruses 2019, 11, 282. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Pan, Y.; Wang, D.; Tian, X.; Song, Y.; Cao, Y. Identification and pathogenicity of a variant porcine epidemic diarrhea virus field strain with reduced virulence. Virol. J. 2015, 12, 88. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Dong, B.; Dai, A.; Dai, A.; Li, X.; Li, X.; Yang, X.; Yang, X. The four of structural genes sequences of a porcine epidemic diarrhea virus from domestic piglet in Fujian, China. Virol. J. 2020, 17, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Xue, C.; He, L.; Wang, Y.; Cao, Y. Bioinformatics insight into the spike glycoprotein gene of field porcine epidemic diarrhea strains during 2011-2013 in Guangdong, China. Virus Genes 2014, 49, 58–67. [Google Scholar] [CrossRef] [PubMed]


| Virus Strain | Abbreviation | Pig Farm | Age of Pig (Days) | Sample | Isolated Time |
|---|---|---|---|---|---|
| TW/PT01/2020 | PT01 | SL | 4 | Intestine | August 2020 |
| TW/PT02/2020 | PT02 | SL | >7 | Intestine | October 2020 |
| TW/PT03/2020 | PT03 | SL | 5 | Intestine | September 2020 |
| TW/PT04/2020 | PT04 | SL | 6 | Intestine | December 2020 |
| TW/PT05/2020 | PT05 | SL | 3 | Feces | December 2020 |
| TW/CY4/2020 | CY4 | CL | 21 | Intestine | December 2020 |
| Primer Name | Nucleotide Sequence, (5′-3′) | Targets |
|---|---|---|
| Full S (F) | TAAGTTGCTAGTGCGTAATAATGAC | S full-length amplification |
| Full S (R) | CAGACTTCGAGACATCTTTG | |
| PED.sq1 | CTTTGTTAGCCATATTAGAGGT | Sequencing of the spike gene |
| PED.sq2 | ACGATCGATGGTGTTTGTAATGGA | |
| PED.sq3 | GGACACTAATTGCCCTTTCACCT | |
| PED.sq4 | ACGCCTGTTAGTGTTGATTG | |
| PED.sq5 | TTGAGAGTGTTAAAGAGGCTATT | |
| PED.sq6 | ACTCTCGACTGGACATTC | |
| PED.sq7 | AGCCATTTCTAGTTCTATTG |
| Detection Methods | No. of Sows | Antibody Titers | |
|---|---|---|---|
| Before Feedback | 2 Weeks after Feedback | ||
| Serum neutralization | 15 | 3.7 ± 0.6 a,* | 21.5 ± 5.8 b |
| ELISA | 15 | 20.1 ± 2.7 a | 27.5 ± 5.0 b |
| GenBank No. or Strain Name | Year First Identified | Substitution Sites in Antigenic Regions of S Protein * | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COE Domain (Residues 502–641) | SS6 (Residues 767–774) | ||||||||||||||||||||||||
| 509 | 520 | 524 | 526 | 531 | 532 | 537 | 541 | 554 | 561 | 566 | 573 | 581 | 594 | 603 | 604 | 606 | 608 | 612 | 615 | 624 | 626 | 636 | 769 | ||
| KP276251.1 (15) ** | 2013 | D | S | H | G | A | S | N | S | F | Y | K | P | D | L | L | F | Y | E | G | F | K | E | E | S |
| KJ434295.1 | 2013 | • | • | P | • | • | • | • | • | • | • | • | • | • | • | I | • | • | D | • | • | • | • | • | |
| KM246672.1 | 2013 | • | • | P | • | • | • | S | • | • | • | • | • | • | • | I | • | • | D | • | • | • | • | • | |
| KM246707.1 | 2013 | • | • | • | S | • | • | • | • | • | • | • | • | N | • | • | • | • | • | • | • | • | • | • | |
| KP276252.1 | 2014 | • | • | • | • | • | • | • | • | • | • | N | • | • | • | • | • | • | • | • | • | • | • | • | |
| KJ434308.1 | 2014 | • | • | • | • | • | • | • | • | • | • | • | L | • | • | • | • | • | • | • | • | • | • | • | |
| KJ434306.1 | 2014 | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | E | • | • | |
| KY929405.1 | 2015 | A | • | • | • | • | • | • | • | • | • | N | • | • | • | • | • | • | • | • | • | • | • | • | |
| KY929406.1 | 2016 | A | • | • | • | • | • | • | • | S | • | N | • | • | • | • | • | • | • | • | • | • | • | • | |
| MK673523.1 | 2016 | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | V | |
| MK673526.1 | 2017 | • | • | • | • | • | • | • | • | • | • | N | • | • | • | • | L | • | • | A | • | • | • | • | |
| MK673531.1 | 2017 | • | • | • | • | • | • | • | • | • | • | N | • | • | • | • | • | • | • | V | • | • | • | • | |
| MK673517.1 | 2017 | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | V | |
| MK673535.1 | 2017 | • | • | P | • | • | I | • | Y | • | • | • | • | • | • | • | • | H | D | • | • | • | • | • | |
| MK673545.1 | 2018 | • | • | • | • | V | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | |
| MK673528.1 | 2018 | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | D | V | • | • | • | • | |
| MK673527.1 | 2018 | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | D | V | • | • | • | • | |
| MK673520.1 | 2018 | • | • | • | • | • | • | • | • | • | • | N | • | • | • | • | • | • | • | • | • | • | • | • | |
| KM673538.1 (2) | 2018 | • | • | • | • | • | • | • | • | • | • | N | • | • | F | • | • | H | A | D | • | • | • | • | |
| KM246732.1 | 2018 | • | F | S | • | • | • | • | • | • | • | • | • | • | • | • | • | • | D | • | Y | • | D | • | |
| CY4 | 2020 | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | |
| PT01 (3) | 2020 | • | • | • | • | • | • | S | • | • | H | • | • | • | • | • | • | • | • | • | • | • | • | • | |
| PT03 (2) | 2020 | S | H | F | |||||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen Thi, T.H.; Chen, C.-C.; Chung, W.-B.; Chaung, H.-C.; Huang, Y.-L.; Cheng, L.-T.; Ke, G.-M. Antibody Evaluation and Mutations of Antigenic Epitopes in the Spike Protein of the Porcine Epidemic Diarrhea Virus from Pig Farms with Repeated Intentional Exposure (Feedback). Viruses 2022, 14, 551. https://doi.org/10.3390/v14030551
Nguyen Thi TH, Chen C-C, Chung W-B, Chaung H-C, Huang Y-L, Cheng L-T, Ke G-M. Antibody Evaluation and Mutations of Antigenic Epitopes in the Spike Protein of the Porcine Epidemic Diarrhea Virus from Pig Farms with Repeated Intentional Exposure (Feedback). Viruses. 2022; 14(3):551. https://doi.org/10.3390/v14030551
Chicago/Turabian StyleNguyen Thi, Thu Hien, Chi-Chih Chen, Wen-Bin Chung, Hso-Chi Chaung, Yen-Li Huang, Li-Ting Cheng, and Guan-Ming Ke. 2022. "Antibody Evaluation and Mutations of Antigenic Epitopes in the Spike Protein of the Porcine Epidemic Diarrhea Virus from Pig Farms with Repeated Intentional Exposure (Feedback)" Viruses 14, no. 3: 551. https://doi.org/10.3390/v14030551
APA StyleNguyen Thi, T. H., Chen, C.-C., Chung, W.-B., Chaung, H.-C., Huang, Y.-L., Cheng, L.-T., & Ke, G.-M. (2022). Antibody Evaluation and Mutations of Antigenic Epitopes in the Spike Protein of the Porcine Epidemic Diarrhea Virus from Pig Farms with Repeated Intentional Exposure (Feedback). Viruses, 14(3), 551. https://doi.org/10.3390/v14030551