
Identification of SARS-CoV-2 Spike Palmitoylation Inhibitors That Results in Release of Attenuated Virus with Reduced Infectivity



 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids and HIV-1-Derived Pseudovirus Generation
2.2. Cell Culture
2.3. CoV S Protein Sequence Alignment
2.4. Site-Directed Mutagenesis
2.5. Transfection
2.6. SARS-CoV-2 S Protein Pseudotyped Virus Entry
2.7. Pseudovirus Egress
2.8. Immunoblotting
2.9. Co-Immunoprecipitation (Co-IP)
2.10. Acyl-PEGyl Exchange Gel-Shift (APEGS) Assay
2.11. Syncytium Formation Assay
2.12. Compound Synthesis and Characterization
2.13. Cellular Toxicity Assay
2.14. Gene Silencing
2.15. RNA Extraction and RT-qPCR
2.16. Immunofluorescence Assay (IFA) and Proximity Ligation Assay (PLA)
2.17. Surface Immunofluorescence Assay
2.18. Spike Surface Abundance by Flow Cytometry
2.19. SARS-CoV-2 Virus Stock Preparation and Titration with Plaque-Based Assays
2.20. SARS-CoV-2 Infection
2.21. Drug Treatments
2.22. Statistical Analysis and Reproducibility
3. Results
3.1. SARS-CoV-2 S Protein Is Palmitoylated on Multiple Sites
3.2. Spike Protein Palmitoylation Is Required for Infection of Cells Expressing ACE2
3.3. Effect of Palmitoylation on the Plasma Membrane Localization, Viral Egress, and ACE2 Binding of SARS-CoV-2 S Protein
3.4. Palmitoylation Is Important for S Protein-Mediated Membrane Fusion and Syncytia Formation
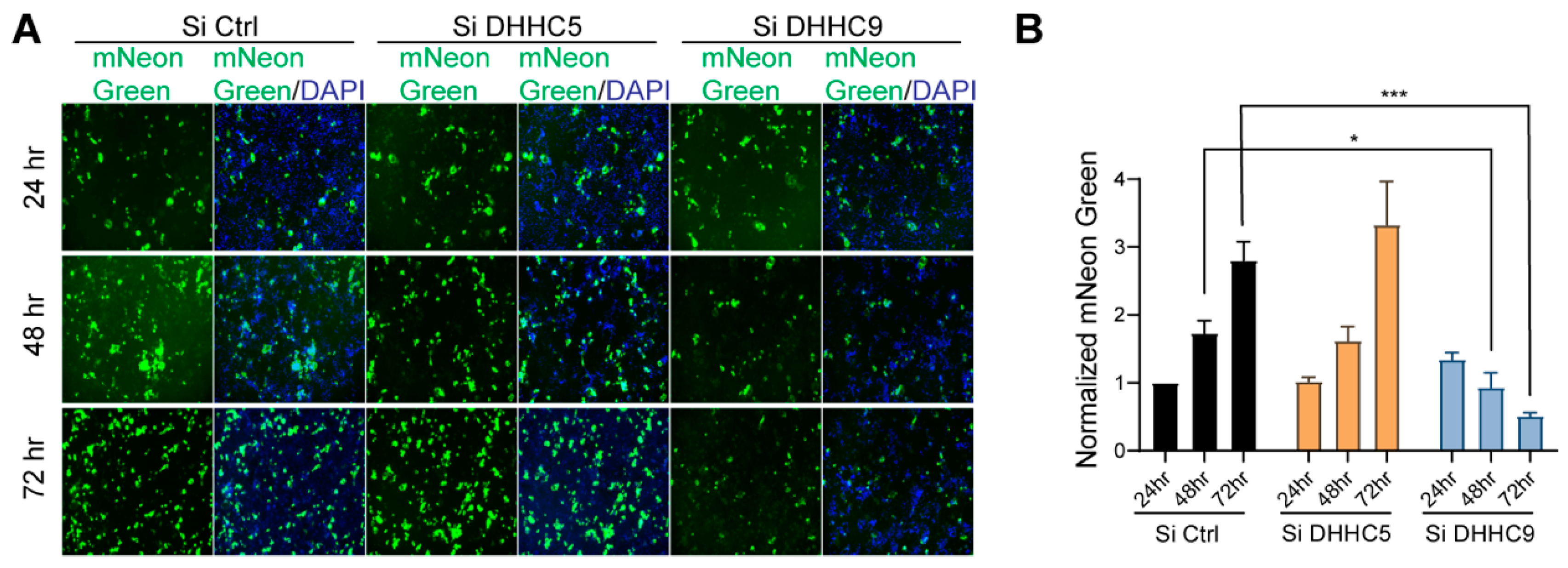
3.5. DHHC9 PAT Is Instrumental in SARS-CoV-2 S Protein Palmitoylation
3.6. DHHC9 Co-Localizes and Interacts with the SARS-CoV-2 S Protein Both in Transfected and Infected Cells
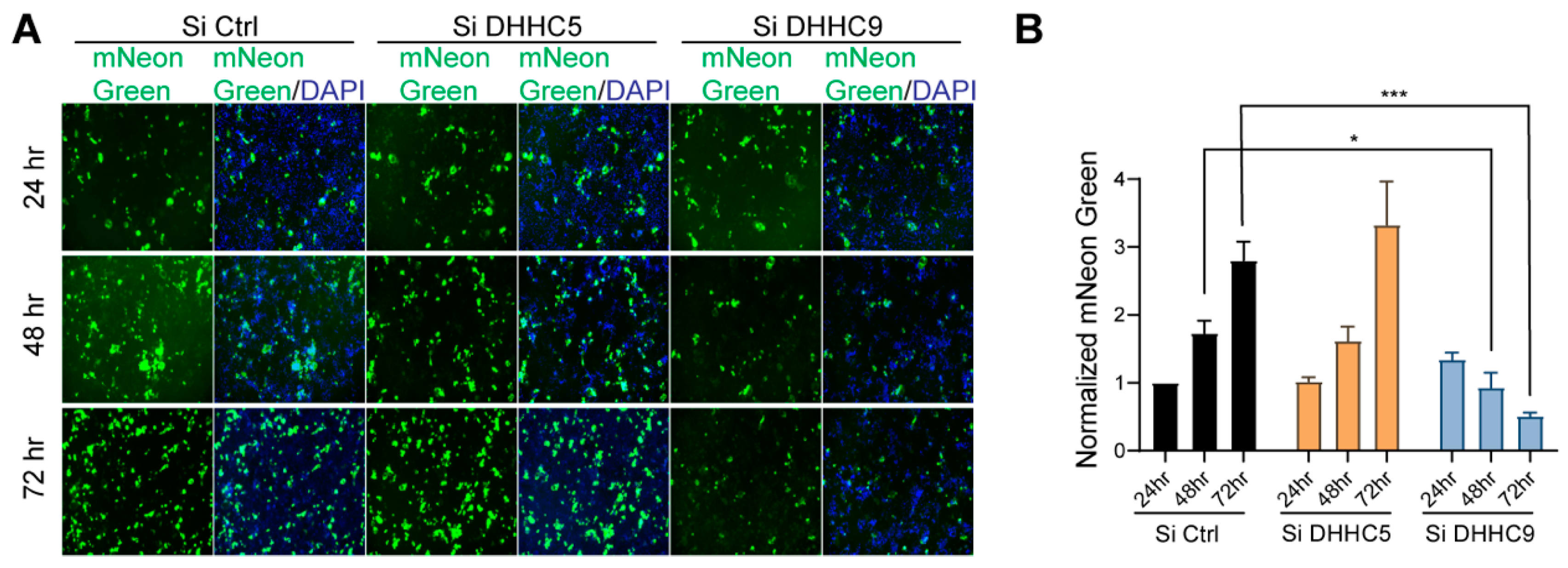
3.7. SARS-CoV-2 Infection and Syncytia Formation in Caco-2 Cells Requires DHHC9-Dependent Palmitoylation of S Protein
3.8. Novel Bis-Piperazine DHHC9 Inhibitors Inhibit SARS-CoV-2 S Palmitoylation, Fusogenicity and Infectivity
3.9. Compounds 13 and 25 Inhibit SARS-CoV-2 Infection in Cell Culture
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Zhang, S.; Wang, Q.; Anang, S.; Wang, J.; Ding, H.; Kappes, J.C.; Sodroski, J. Spike glycoprotein and host cell determinants of SARS-CoV-2 entry and cytopathic effects. J. Virol. 2020, 95, e02304-20. [Google Scholar] [CrossRef] [PubMed]
- Qing, E.; Kicmal, T.; Kumar, B.; Hawkins, G.M.; Timm, E.; Perlman, S.; Gallagher, T. Dynamics of SARS-CoV-2 Spike Proteins in Cell Entry: Control Elements in the Amino-Terminal Domains. mBio 2021, 12, e0159021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Tsybovsky, Y.; Gorman, J.; Rapp, M.; Cerutti, G.; Chuang, G.Y.; Katsamba, P.S.; Sampson, J.M.; Schon, A.; Bimela, J.; et al. Cryo-EM Structures of SARS-CoV-2 Spike without and with ACE2 Reveal a pH-Dependent Switch to Mediate Endosomal Positioning of Receptor-Binding Domains. Cell Host Microbe 2020, 28, 867–879.e5. [Google Scholar] [CrossRef]
- Puthenveetil, R.; Lun, C.M.; Murphy, R.E.; Healy, L.B.; Vilmen, G.; Christenson, E.T.; Freed, E.O.; Banerjee, A. S-acylation of SARS-CoV-2 Spike Protein: Mechanistic Dissection, In Vitro Reconstitution and Role in Viral Infectivity. J. Biol. Chem. 2021, 297, 101112. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, Z.; Wang, X.; Zhang, J.; Ren, C.; Li, Y.; Gao, L.; Liang, X.; Wang, P.; Ma, C. Palmitoylation of SARS-CoV-2 S protein is essential for viral infectivity. Signal. Transduct Target. Ther. 2021, 6, 231. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.T.; Yu, X.T.; Cheng, W. The interactions of ZDHHC5/GOLGA7 with SARS-CoV-2 spike (S) protein and their effects on S protein’s subcellular localization, palmitoylation and pseudovirus entry. Virol. J. 2021, 18, 257. [Google Scholar] [CrossRef]
- Mesquita, F.S.; Abrami, L.; Sergeeva, O.; Turelli, P.; Qing, E.; Kunz, B.; Raclot, C.; Paz Montoya, J.; Abriata, L.A.; Gallagher, T.; et al. S-acylation controls SARS-CoV-2 membrane lipid organization and enhances infectivity. Dev. Cell 2021, 56, 2790–2807.e8. [Google Scholar] [CrossRef]
- Li, D.; Liu, Y.; Lu, Y.; Gao, S.; Zhang, L. Palmitoylation of SARS-CoV-2 S protein is critical for S-mediated syncytia formation and virus entry. J. Med. Virol. 2022, 94, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, T.; Tsutsumi, R.; Noritake, J.; Fukata, Y.; Fukata, M. Dynamic protein palmitoylation in cellular signaling. Prog. Lipid. Res. 2009, 48, 117–127. [Google Scholar] [CrossRef]
- Mitchell, D.A.; Vasudevan, A.; Linder, M.E.; Deschenes, R.J. Protein palmitoylation by a family of DHHC protein S-acyltransferases. J. Lipid Res. 2006, 47, 1118–1127. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, C.D.; Linder, M.E. Structure and function of DHHC protein S-acyltransferases. Biochem. Soc. Trans. 2017, 45, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.F.; Bracha, M.; Schlesinger, M.J. Evidence for covalent attachment of fatty acids to Sindbis virus glycoproteins. Proc. Natl Acad. Sci. USA 1979, 76, 1687–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.F.; Schlesinger, M.J. Fatty acid binding to vesicular stomatitis virus glycoprotein: A new type of post-translational modification of the viral glycoprotein. Cell 1979, 17, 813–819. [Google Scholar] [CrossRef]
- Lobo, S.; Greentree, W.K.; Linder, M.E.; Deschenes, R.J. Identification of a Ras palmitoyltransferase in Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 41268–41273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, A.F.; Feng, Y.; Chen, L.; Davis, N.G. The yeast DHHC cysteine-rich domain protein Akr1p is a palmitoyl transferase. J. Cell. Biol. 2002, 159, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, M.; David, F.P.A.; van der Goot, F.G. SwissPalm 2: Protein S-Palmitoylation Database. Methods Mol. Biol. 2019, 2009, 203–214. [Google Scholar]
- Linder, M.E.; Deschenes, R.J. Palmitoylation: Policing protein stability and traffic. Nat. Rev. Mol. Cell Biol. 2007, 8, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, X.; Chen, X.; Aramsangtienchai, P.; Tong, Z.; Lin, H. Protein Lipidation: Occurrence, Mechanisms, Biological Functions, and Enabling Technologies. Chem. Rev. 2018, 118, 919–988. [Google Scholar] [CrossRef] [PubMed]
- Veit, M. Palmitoylation of virus proteins. Biol. Cell 2012, 104, 493–515. [Google Scholar] [CrossRef] [PubMed]
- Thorp, E.B.; Boscarino, J.A.; Logan, H.L.; Goletz, J.T.; Gallagher, T.M. Palmitoylations on murine coronavirus spike proteins are essential for virion assembly and infectivity. J. Virol. 2006, 80, 1280–1289. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lv, J.; Wang, Y.; Gao, S.; Yao, Q.; Qu, D.; Ye, R. Replication of murine coronavirus requires multiple cysteines in the endodomain of spike protein. Virology 2012, 427, 98–106. [Google Scholar] [CrossRef]
- Petit, C.M.; Chouljenko, V.N.; Iyer, A.; Colgrove, R.; Farzan, M.; Knipe, D.M.; Kousoulas, K.G. Palmitoylation of the cysteine-rich endodomain of the SARS-coronavirus spike glycoprotein is important for spike-mediated cell fusion. Virology 2007, 360, 264–274. [Google Scholar] [CrossRef] [Green Version]
- McBride, C.E.; Machamer, C.E. Palmitoylation of SARS-CoV S protein is necessary for partitioning into detergent-resistant membranes and cell-cell fusion but not interaction with M protein. Virology 2010, 405, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Levental, I.; Levental, K.R.; Heberle, F.A. Lipid rafts: Controversies resolved, mysteries remain. Trends Cell Biol. 2020, 30, 341–353. [Google Scholar] [CrossRef]
- Buchrieser, J.; Dufloo, J.; Hubert, M.; Monel, B.; Planas, D.; Rajah, M.M.; Planchais, C.; Porrot, F.; Guivel-Benhassine, F.; Van der Werf, S.; et al. Syncytia formation by SARS-CoV-2-infected cells. EMBO J. 2020, 39, e106267. [Google Scholar] [CrossRef]
- Cattin-Ortola, J.; Welch, L.G.; Maslen, S.L.; Papa, G.; James, L.C.; Munro, S. Sequences in the cytoplasmic tail of SARS-CoV-2 Spike facilitate expression at the cell surface and syncytia formation. Nat. Commun. 2021, 12, 5333. [Google Scholar] [CrossRef]
- Lin, L.; Li, Q.; Wang, Y.; Shi, Y. Syncytia formation during SARS-CoV-2 lung infection: A disastrous unity to eliminate lymphocytes. Cell Death Differ. 2021, 28, 2019–2021. [Google Scholar] [CrossRef]
- Gadalla, M.R.; Veit, M. Toward the identification of ZDHHC enzymes required for palmitoylation of viral protein as potential drug targets. Expert Opin. Drug Discov. 2020, 15, 159–177. [Google Scholar] [CrossRef]
- Santos-Beneit, F.; Raskevicius, V.; Skeberdis, V.A.; Bordel, S. A metabolic modeling approach reveals promising therapeutic targets and antiviral drugs to combat COVID-19. Sci. Rep. 2021, 11, 11982. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Lee, M.; Sugiyama, M.; Mekhail, K.; Latreille, E.; Khosraviani, N.; Wei, K.; Lee, W.L.; Antonescu, C.; Fairn, G.D. Fatty Acid Synthase inhibition prevents palmitoylation of SARS-CoV2 Spike Protein and improves survival of mice infected with murine hepatitis virus. bioRxiv 2020. [Google Scholar] [CrossRef]
- Davda, D.; El Azzouny, M.A.; Tom, C.T.; Hernandez, J.L.; Majmudar, J.D.; Kennedy, R.T.; Martin, B.R. Profiling targets of the irreversible palmitoylation inhibitor 2-bromopalmitate. ACS Chem. Biol. 2013, 8, 1912–1917. [Google Scholar] [CrossRef] [Green Version]
- Chavda, B.; Arnott, J.A.; Planey, S.L. Targeting protein palmitoylation: Selective inhibitors and implications in disease. Expert Opin. Drug Discov. 2014, 9, 1005–1019. [Google Scholar] [CrossRef]
- Hamel, L.D.; Lenhart, B.J.; Mitchell, D.A.; Santos, R.G.; Giulianotti, M.A.; Deschenes, R.J. Identification of Protein Palmitoylation Inhibitors from a Scaffold Ranking Library. Comb. Chem. High Throughput Screen 2016, 19, 262–274. [Google Scholar] [CrossRef]
- Dull, T.; Zufferey, R.; Kelly, M.; Mandel, R.J.; Nguyen, M.; Trono, D.; Naldini, L. A third-generation lentivirus vector with a conditional packaging system. J. Virol. 1998, 72, 8463–8471. [Google Scholar] [CrossRef] [Green Version]
- Crawford, K.H.D.; Eguia, R.; Dingens, A.S.; Loes, A.N.; Malone, K.D.; Wolf, C.R.; Chu, H.Y.; Tortorici, M.A.; Veesler, D.; Murphy, M.; et al. Protocol and Reagents for Pseudotyping Lentiviral Particles with SARS-CoV-2 Spike Protein for Neutralization Assays. Viruses 2020, 12, 513. [Google Scholar] [CrossRef]
- Percher, A.; Thinon, E.; Hang, H. Mass-Tag Labeling Using Acyl-PEG Exchange for the Determination of Endogenous Protein S-Fatty Acylation. Curr Protoc. Protein Sci. 2017, 89, 14–17. [Google Scholar]
- McClafferty, H.; Shipston, M.J. siRNA Knockdown of Mammalian zDHHCs and Validation of mRNA Expression by RT-qPCR. Methods Mol. Biol. 2019, 2009, 151–168. [Google Scholar]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brunink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. 2020, 25, 2000045. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Muruato, A.; Lokugamage, K.G.; Narayanan, K.; Zhang, X.; Zou, J.; Liu, J.; Schindewolf, C.; Bopp, N.E.; Aguilar, P.V.; et al. An Infectious cDNA Clone of SARS-CoV-2. Cell Host Microbe 2020, 27, 841–848.e3. [Google Scholar] [CrossRef]
- Rodenburg, R.N.P.; Snijder, J.; van de Waterbeemd, M.; Schouten, A.; Granneman, J.; Heck, A.J.R.; Gros, P. Stochastic palmitoylation of accessible cysteines in membrane proteins revealed by native mass spectrometry. Nat. Commun. 2017, 8, 1280. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. CSS-Palm 2.0: An updated software for palmitoylation sites prediction. Protein Eng. Des. Sel. 2008, 21, 639–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Gibson, B.; Snider, J.; Jenkins, C.M.; Han, X.; Gross, R.W. Submicromolar concentrations of palmitoyl-CoA specifically thioesterify cysteine 244 in glyceraldehyde-3-phosphate dehydrogenase inhibiting enzyme activity: A novel mechanism potentially underlying fatty acid induced insulin resistance. Biochemistry 2005, 44, 11903–11912. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Scudellari, M. How the coronavirus infects cells—And why Delta is so dangerous. Nature 2021, 595, 640–644. [Google Scholar] [CrossRef]
- Swarthout, J.T.; Lobo, S.; Farh, L.; Croke, M.R.; Greentree, W.K.; Deschenes, R.J.; Linder, M.E. DHHC9 and GCP16 constitute a human protein fatty acyltransferase with specificity for H- and N-Ras. J. Biol. Chem. 2005, 280, 31141–31148. [Google Scholar] [CrossRef] [Green Version]
- Ohta, E.; Misumi, Y.; Sohda, M.; Fujiwara, T.; Yano, A.; Ikehara, Y. Identification and characterization of GCP16, a novel acylated Golgi protein that interacts with GCP170. J. Biol. Chem. 2003, 278, 51957–51967. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Marsden, A.N.; Scott, C.A.; Akimzhanov, A.M.; Boehning, D. DHHC5 Mediates beta-Adrenergic Signaling in Cardiomyocytes by Targeting Galpha Proteins. Biophys. J. 2020, 118, 826–835. [Google Scholar] [CrossRef] [Green Version]
- Woodley, K.T.; Collins, M.O. S-acylated Golga7b stabilises DHHC5 at the plasma membrane to regulate cell adhesion. EMBO Rep. 2019, 20, e47472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredriksson, S.; Gullberg, M.; Jarvius, J.; Olsson, C.; Pietras, K.; Gustafsdottir, S.M.; Ostman, A.; Landegren, U. Protein detection using proximity-dependent DNA ligation assays. Nat. Biotechnol. 2002, 20, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, H.; Tousson, A.; Circolo, A.; Marchase, R.B.; Volanakis, J.E. Calnexin is associated with and induced by overexpressed human complement protein C2. Anat. Rec. 2002, 267, 7–16. [Google Scholar] [CrossRef]
- Myhill, N.; Lynes, E.M.; Nanji, J.A.; Blagoveshchenskaya, A.D.; Fei, H.; Carmine Simmen, K.; Cooper, T.J.; Thomas, G.; Simmen, T. The subcellular distribution of calnexin is mediated by PACS-2. Mol. Biol. Cell 2008, 19, 2777–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draper, J.M.; Smith, C.D. Palmitoyl acyltransferase assays and inhibitors (Review). Mol. Membr. Biol. 2009, 26, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, R.A.; Rao, P.; Fogelsong, R.J.; Bardes, E.S. 2-Bromopalmitoyl-CoA and 2-bromopalmitate: Promiscuous inhibitors of membrane-bound enzymes. Biochim. Biophys. Acta 1992, 1125, 203–209. [Google Scholar] [CrossRef]
- Chase, J.F.; Tubbs, P.K. Specific inhibition of mitochondrial fatty acid oxidation by 2-bromopalmitate and its coenzyme A and carnitine esters. Biochem. J. 1972, 129, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedro, M.P.; Vilcaes, A.A.; Tomatis, V.M.; Oliveira, R.G.; Gomez, G.A.; Daniotti, J.L. 2-Bromopalmitate reduces protein deacylation by inhibition of acyl-protein thioesterase enzymatic activities. PLoS ONE 2013, 8, e75232. [Google Scholar]
- Greaves, J.; Chamberlain, L.H. Palmitoylation-dependent protein sorting. J. Cell Biol. 2007, 176, 249–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentz, J.; Mittal, A. Architecture of the influenza hemagglutinin membrane fusion site. Biochim. Biophys. Acta 2003, 1614, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shulla, A.; Gallagher, T. Role of spike protein endodomains in regulating coronavirus entry. J. Biol. Chem. 2009, 284, 32725–32734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulrahman, D.A.; Meng, X.; Veit, M. S-Acylation of Proteins of Coronavirus and Influenza Virus: Conservation of Acylation Sites in Animal Viruses and DHHC Acyltransferases in Their Animal Reservoirs. Pathogens 2021, 10, 669. [Google Scholar] [CrossRef] [PubMed]
- Zaballa, M.E.; van der Goot, F.G. The molecular era of protein S-acylation: Spotlight on structure, mechanisms, and dynamics. Crit Rev. Biochem. Mol. Biol. 2018, 53, 420–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Source |
|---|---|---|
| C1_F | TATAATGCTGAGCAGCATGACAAGCTG | This Study |
| C1_R | GTCACCATGACTATAGCG | This Study |
| C2_F | AGCAGTCTCAAAGGCTGTTGCTCTTG | This Study |
| C2_R | GCTGCTGCTTGTCATGCAGCACAG | This Study |
| C3_F | TCTAGCGGCTCTTGCTGCAAATTC | This Study |
| C3_R | GCTACTGCCTTTGAGACAGCTGCA | This Study |
| C4_F | TTGCGGCTCTAGCAGCAAATTCGATG | This Study |
| C4_R | GAGCAACAGCCTTTGAGAC | This Study |
| ΔC_F | AAAGGCAGTAGCTCTAGCGGCTCTAGCAGCAAATTCGATGAGGACGATTC | This Study |
| ΔC_R | GAGACTGCTGCTGCTGCTTGTCATGCTGCTCAGCATTATAGTCACCATG | This Study |
| Antibody | Source | Catalog Number |
|---|---|---|
| Spike S2 | GeneTex | GTX632604 |
| Spike S1 | Sino Biological | 40591-MM42 |
| ACE2 | Novus | NBP2-67692 |
| GAPDH | Santa Cruz | sc-47724 |
| Calnexin | Novus | NB300-518 |
| Calnexin | Cell Signaling | 2679P |
| FLAG | Sigma-Aldrich | F1804 |
| FLAG | Sigma-Aldrich | F7425 |
| Myc | Cell Signaling | 71D10 |
| Myc | Cell Signaling | 9B11 |
| Primer | Sequence1 | Sequence2 | Source |
|---|---|---|---|
| DHHC1 | CAAGCCCTCCAACAAGACG | CCAAAGCCGATCACAGCAAAG | [39] |
| DHHC2 | AACACTGGCGAACAAGTTGTG | AGATGGGAAGATCCTTGGCTG | [39] |
| DHHC3 | CCACTTCCGAAACATTGAGCG | CCACAGCCGTCACGGATAAA | [39] |
| DHHC4 | CCTGACTTGTGGAACCAATCC | GCACCTCACGTTCTTTGGAAAC | [39] |
| DHHC5 | CACCTGCCGCTTTTACCGT | CGGCGACCAATACAGTTATTCAC | [39] |
| DHHC6 | AGTCTGCCAAGCATACAAGGC | CCAGTGGTGCTAAAAGGAGAAAC | [39] |
| DHHC7 | CTGACCGGGTCTGGTTCATC | CATGACGAAAGTCACCACGAA | [39] |
| DHHC8 | GTATCCAGGTCCGCATGAAGT | AGCGTGGTTCAGCACGTAG | [39] |
| DHHC9 | CCCAGGCAGGAACACCTTTT | CCGAGGAATCACTCCAGGG | [39] |
| DHHC11 | GGTGCAGACCCTGATAGTCG | GCACGTATGGATCTTTCCTCAC | [39] |
| DHHC12 | GTGCTGACCTGGGGAATCAC | CTGCACATTCACGTAGCCA | [39] |
| DHHC13 | ACCCCACTCTTATTGATGGAGA | TGTCTGCCCATTTACATCTGTC | [39] |
| DHHC14 | TGTGATAACTGCGTAGAACGGT | CGTGGGTGATAACGAATGCAA | [39] |
| DHHC15 | GGTGCCAGTGCTCGTTATTGT | AAGACGTAGGCATAGTAGGACC | [39] |
| DHHC16 | ACTCCGGGGTCTAGTACAGC | CCAGCGGATCACGTTGTCT | [39] |
| DHHC17 | GGCCCGGATGAGTACGATAC | TCCAAGAGGTTCACCATATCCA | [39] |
| DHHC18 | TGACGGCCTTCATCTTCGC | CTGGACCACGAGCCTTTGAT | [39] |
| DHHC19 | TTGCTGCCTTCAATGTGGTG | CGGAGCCTTGATGTAAGATGC | [39] |
| DHHC20 | CGCACCCACGTTTTCATACG | TCTGGCATACTCATTCTGGTTTG | [39] |
| DHHC21 | TGTTGTTGACCCACATGGTTG | GAGGCCCTCACTAAGGCAA | [39] |
| DHHC22 | GAGGCACGACCATCACTGTTT | ACAGCGGAGATGTAGGCCA | [39] |
| DHHC23 | TCTGGATGAAGGGTGTGATCG | GCTCCCCTAAGCCAAGGAA | [39] |
| DHHC24 | CTGGCACAGTTTGCCTTGG | CAGGGACCCAGGTCATAGGAG | [39] |
| SARS-CoV-2 N | CACATTGGCACCCGCAATC | GAGGAACGAGAAGAGGCTTG | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramadan, A.A.; Mayilsamy, K.; McGill, A.R.; Ghosh, A.; Giulianotti, M.A.; Donow, H.M.; Mohapatra, S.S.; Mohapatra, S.; Chandran, B.; Deschenes, R.J.; et al. Identification of SARS-CoV-2 Spike Palmitoylation Inhibitors That Results in Release of Attenuated Virus with Reduced Infectivity. Viruses 2022, 14, 531. https://doi.org/10.3390/v14030531
Ramadan AA, Mayilsamy K, McGill AR, Ghosh A, Giulianotti MA, Donow HM, Mohapatra SS, Mohapatra S, Chandran B, Deschenes RJ, et al. Identification of SARS-CoV-2 Spike Palmitoylation Inhibitors That Results in Release of Attenuated Virus with Reduced Infectivity. Viruses. 2022; 14(3):531. https://doi.org/10.3390/v14030531
Chicago/Turabian StyleRamadan, Ahmed A., Karthick Mayilsamy, Andrew R. McGill, Anandita Ghosh, Marc A. Giulianotti, Haley M. Donow, Shyam S. Mohapatra, Subhra Mohapatra, Bala Chandran, Robert J. Deschenes, and et al. 2022. "Identification of SARS-CoV-2 Spike Palmitoylation Inhibitors That Results in Release of Attenuated Virus with Reduced Infectivity" Viruses 14, no. 3: 531. https://doi.org/10.3390/v14030531
APA StyleRamadan, A. A., Mayilsamy, K., McGill, A. R., Ghosh, A., Giulianotti, M. A., Donow, H. M., Mohapatra, S. S., Mohapatra, S., Chandran, B., Deschenes, R. J., & Roy, A. (2022). Identification of SARS-CoV-2 Spike Palmitoylation Inhibitors That Results in Release of Attenuated Virus with Reduced Infectivity. Viruses, 14(3), 531. https://doi.org/10.3390/v14030531