
Screening and Identification of Lassa Virus Entry Inhibitors from a Fragment-Based Drug Discovery Library


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells Lines
2.2. Viruses
2.3. Optimization of High-Throughput Screening (HTS) Assay Conditions
2.4. HTS Assay of the Fragment Drug Library
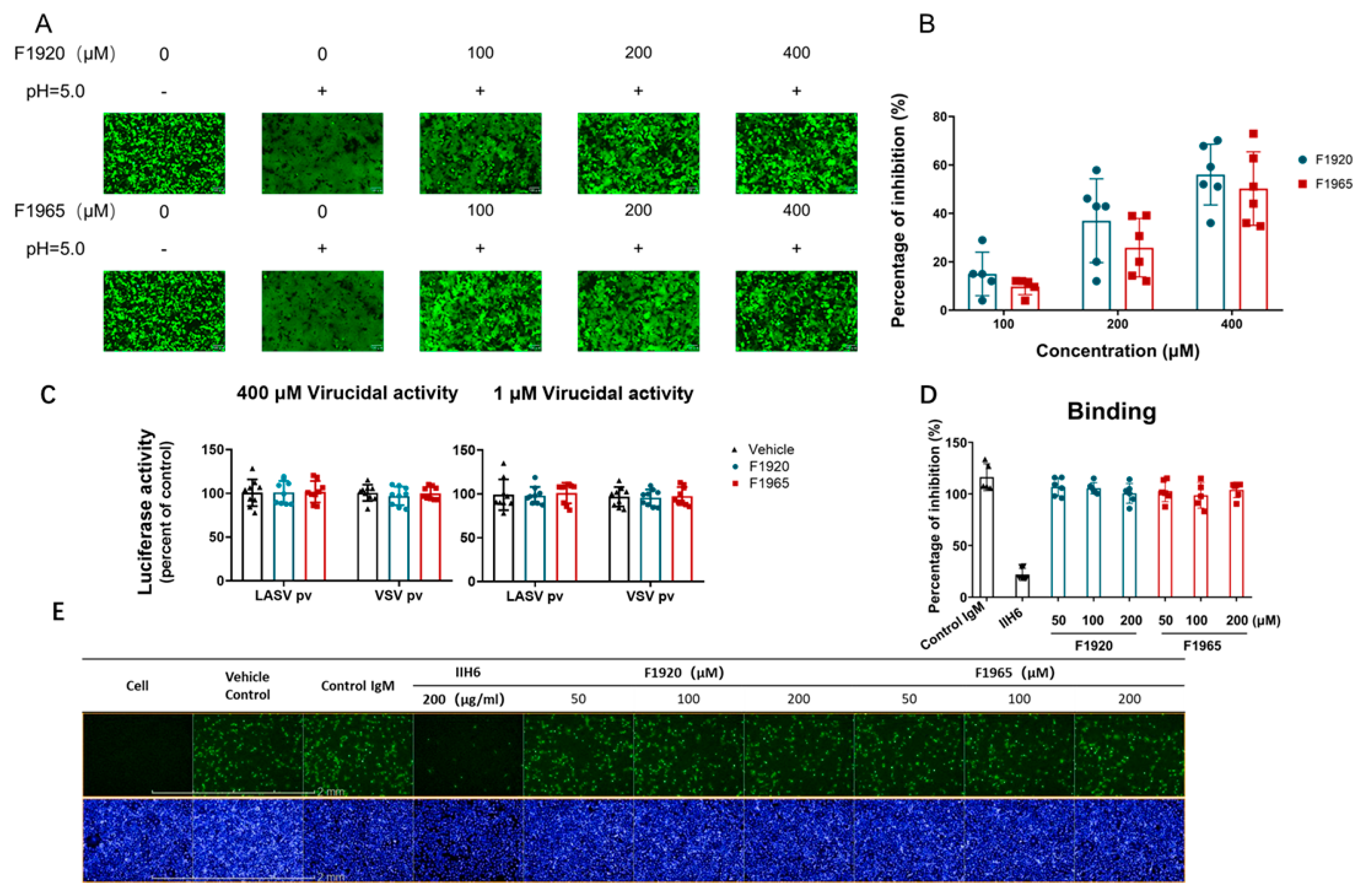
2.5. Membrane Fusion Assay
2.6. Virucidal Assay
2.7. IIH6 Inhibition Assay
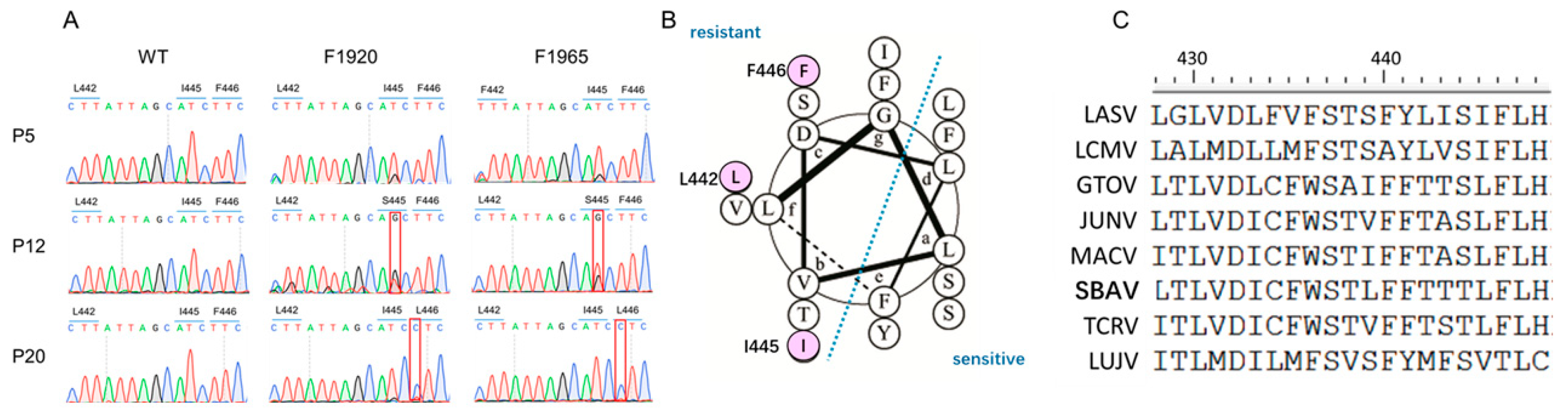
2.8. Selection of Adaptive Mutants
2.9. Inhibition of F1920 or F1965 against Authentic LCMV Infection
2.10. Inhibition of F1920 or F1965 against Recombinant Viruses Infection
3. Results
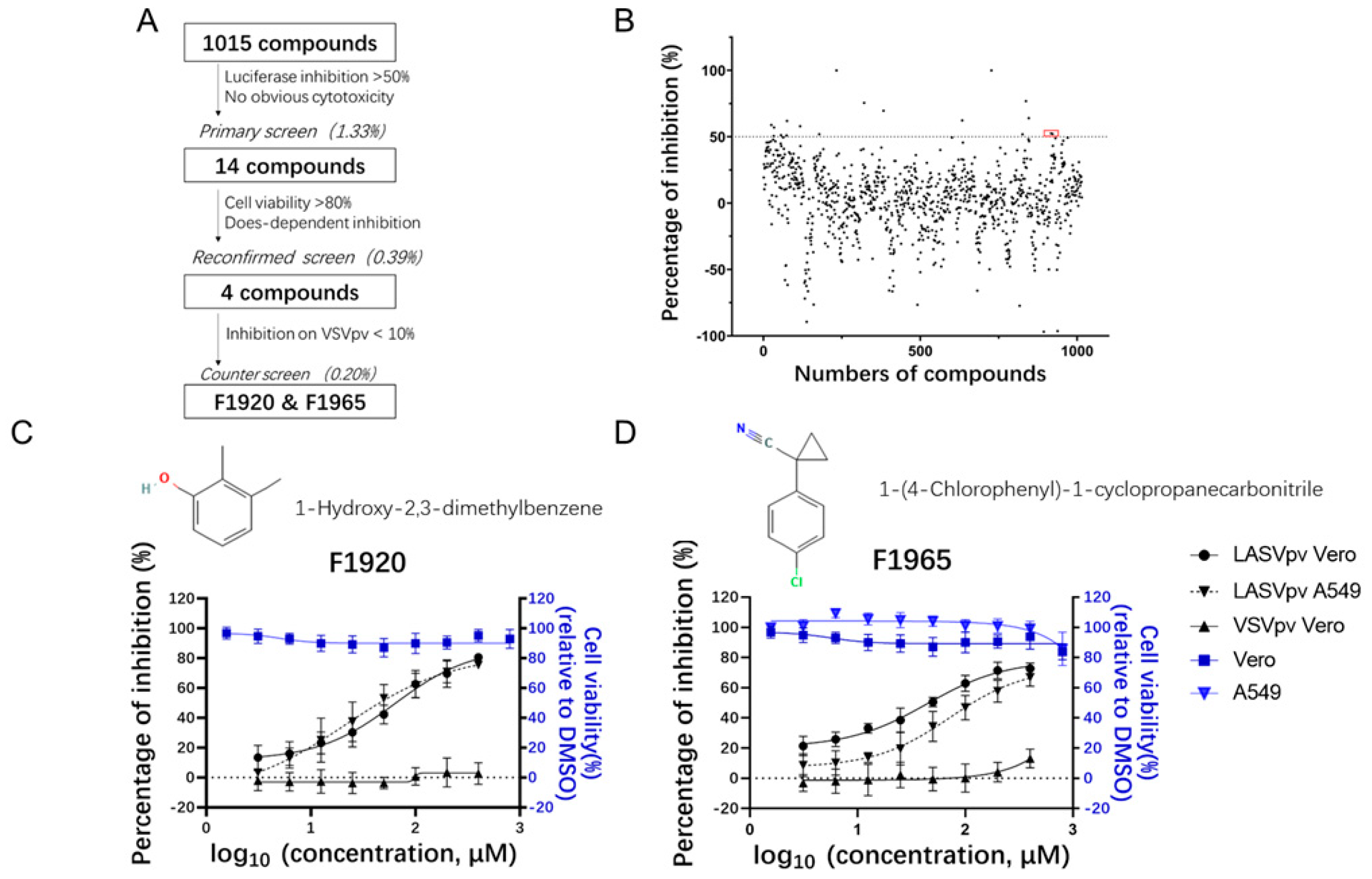
3.1. Screening of an FBDD Library for Entry Inhibitors against LASV
3.2. F1920 and F1965 Inhibit LASV GPC-Mediated Membrane Fusion
3.3. Mutations Located in GP2 Transmembrane Domain (TM) Confer Resistance to Hits
3.4. F1920 and F1965 Showed Antiviral Activity against Other Mammarena Viruses
3.5. Validation of the Antiviral Effects with Replication-Competent Recombinant Viruses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuhn, J.H.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Amarasinghe, G.K.; Anthony, S.J.; Avsic-Zupanc, T.; Ayllon, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. 2020 taxonomic update for phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2020, 165, 3023–3072. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.; Weinstein, J.; Eilon-Ashkenazy, M.; Gehring, K.; Cohen-Dvashi, H.; Elad, N.; Fleishman, S.J.; Diskin, R. Structure and receptor recognition by the Lassa virus spike complex. Nature 2022, 603, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Whitby, L.R.; Casquilho-Gray, H.E.; York, J.; Boger, D.L.; Nunberg, J.H. Small-Molecule Fusion Inhibitors Bind the pH-Sensing Stable Signal Peptide-GP2 Subunit Interface of the Lassa Virus Envelope Glycoprotein. J. Virol. 2016, 90, 6799–6807. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, Y.; Zhang, G.; Wang, S.; Guo, J.; Cao, J.; Jia, X.; Zhang, L.; Xiao, G.; Wang, W. Screening and Identification of Lassa Virus Entry Inhibitors from an FDA-Approved Drugs Library. J. Virol. 2018, 92, e00954-18. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Zhang, X.; Guo, Y. Identification of the dietary supplement capsaicin as an inhibitor of Lassa virus entry. Acta Pharm. Sin. B 2020, 10, 789–798. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, Z.; Zhang, L.; Wang, S.; Xiao, G. Structure-function relationship of the mammarenavirus envelope glycoprotein. Virol. Sin. 2016, 31, 380–394. [Google Scholar] [CrossRef]
- Madu, I.G.; Files, M.; Gharaibeh, D.N.; Moore, A.L.; Jung, K.H.; Gowen, B.B.; Dai, D.; Jones, K.F.; Tyavanagimatt, S.R.; Burgeson, J.R.; et al. A potent Lassa virus antiviral targets an arenavirus virulence determinant. PLoS Pathog. 2018, 14, e1007439. [Google Scholar] [CrossRef]
- Dong, X.; Soong, L. Emerging and re-emerging zoonoses are major and global challenges for public health. Zoonoses 2021, 1. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, G.; Zhou, M.; Liu, Y.; Xiao, G.; Wang, W. Characterizing the Lassa Virus Envelope Glycoprotein Membrane Proximal External Region for Its Role in Fusogenicity. Virol. Sin. 2021, 36, 273–280. [Google Scholar] [CrossRef]
- Cao, J.; Dong, S.; Liu, Y.; Zhou, M.; Guo, J.; Jia, X.; Zhang, Y.; Hou, Y.; Tian, M.; Xiao, G.; et al. Screening and Identification of Lujo Virus Entry Inhibitors From an Food and Drug Administration-Approved Drugs Library. Front. Microbiol. 2021, 12, 793519. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, J.; Cao, J.; Zhang, G.; Jia, X.; Wang, P.; Xiao, G.; Wang, W. Screening of Botanical Drugs against Lassa Virus Entry. J. Virol. 2021, 95, e02429-20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Cao, J.; Cai, Y.; Liu, Y.; Li, Y.; Wang, P.; Guo, J.; Jia, X.; Zhang, M.; Xiao, G.; et al. Structure-activity relationship optimization for lassa virus fusion inhibitors targeting the transmembrane domain of GP2. Protein Cell 2019, 10, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Whitt, M.A. Generation of VSV pseudotypes using recombinant DeltaG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J. Virol. Methods 2010, 169, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.B.; de la Torre, J.C. Rescue of the prototypic Arenavirus LCMV entirely from plasmid. Virology 2006, 350, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Zhang, Y.; Jia, X.; Dong, S.; Liu, Y.; Zhang, M.; Guo, J.; Cao, J.; Guo, Y.; Xiao, G.; et al. Screening and identification of Lassa virus endonuclease-targeting inhibitors from a fragment-based drug discovery library. Antivir. Res. 2022, 197, 105230. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.J.; Casquilho-Gray, H.E.; York, J.; DeCamp, D.L.; Dai, D.; Petrilli, E.B.; Boger, D.L.; Slayden, R.A.; Amberg, S.M.; Sprang, S.R.; et al. A specific interaction of small molecule entry inhibitors with the envelope glycoprotein complex of the Junin hemorrhagic fever arenavirus. J. Biol. Chem. 2011, 286, 6192–6200. [Google Scholar] [CrossRef]
- Hulseberg, C.E.; Feneant, L.; Szymanska, K.M.; White, J.M. Lamp1 Increases the Efficiency of Lassa Virus Infection by Promoting Fusion in Less Acidic Endosomal Compartments. mBio 2018, 9, e01818-17. [Google Scholar] [CrossRef]
- Takikawa, S.; Ishii, K.; Aizaki, H.; Suzuki, T.; Asakura, H.; Matsuura, Y.; Miyamura, T. Cell fusion activity of hepatitis C virus envelope proteins. J. Virol. 2000, 74, 5066–5074. [Google Scholar] [CrossRef]
- Rathbun, J.Y.; Droniou, M.E.; Damoiseaux, R.; Haworth, K.G.; Henley, J.E.; Exline, C.M.; Choe, H.; Cannon, P.M. Novel Arenavirus Entry Inhibitors Discovered by Using a Minigenome Rescue System for High-Throughput Drug Screening. J. Virol. 2015, 89, 8428–8443. [Google Scholar] [CrossRef]
- Oppliger, J.; Torriani, G.; Herrador, A.; Kunz, S. Lassa Virus Cell Entry via Dystroglycan Involves an Unusual Pathway of Macropinocytosis. J. Virol. 2016, 90, 6412–6429. [Google Scholar] [CrossRef]
- Moraz, M.L.; Pythoud, C.; Turk, R.; Rothenberger, S.; Pasquato, A.; Campbell, K.P.; Kunz, S. Cell entry of Lassa virus induces tyrosine phosphorylation of dystroglycan. Cell. Microbiol. 2013, 15, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Bederka, L.H.; Bonhomme, C.J.; Ling, E.L.; Buchmeier, M.J. Arenavirus stable signal peptide is the keystone subunit for glycoprotein complex organization. mBio 2014, 5, e02063. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, G. DrawCoil 1.0. Available online: https://grigoryanlab.org/drawcoil/ (accessed on 17 October 2022).
- Troelsen, N.S.; Clausen, M.H. Library Design Strategies To Accelerate Fragment-Based Drug Discovery. Chemistry 2020, 26, 11391–11403. [Google Scholar] [CrossRef]
- Bolken, T.C.; Laquerre, S.; Zhang, Y.; Bailey, T.R.; Pevear, D.C.; Kickner, S.S.; Sperzel, L.E.; Jones, K.F.; Warren, T.K.; Amanda Lund, S.; et al. Identification and characterization of potent small molecule inhibitor of hemorrhagic fever New World arenaviruses. Antivir. Res. 2006, 69, 86–97. [Google Scholar] [CrossRef] [PubMed]
- York, J.; Dai, D.; Amberg, S.M.; Nunberg, J.H. pH-induced activation of arenavirus membrane fusion is antagonized by small-molecule inhibitors. J. Virol. 2008, 82, 10932–10939. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.; Liu, Y.; Jia, X.; Zhou, M.; Mao, W.; Dong, S.; Zhang, Y.; Xiao, G.; Wang, W. Screening and Identification of Lassa Virus Entry Inhibitors from a Fragment-Based Drug Discovery Library. Viruses 2022, 14, 2649. https://doi.org/10.3390/v14122649
Hou Y, Liu Y, Jia X, Zhou M, Mao W, Dong S, Zhang Y, Xiao G, Wang W. Screening and Identification of Lassa Virus Entry Inhibitors from a Fragment-Based Drug Discovery Library. Viruses. 2022; 14(12):2649. https://doi.org/10.3390/v14122649
Chicago/Turabian StyleHou, Yuxia, Yang Liu, Xiaoying Jia, Minmin Zhou, Wenting Mao, Siqi Dong, Yueli Zhang, Gengfu Xiao, and Wei Wang. 2022. "Screening and Identification of Lassa Virus Entry Inhibitors from a Fragment-Based Drug Discovery Library" Viruses 14, no. 12: 2649. https://doi.org/10.3390/v14122649
APA StyleHou, Y., Liu, Y., Jia, X., Zhou, M., Mao, W., Dong, S., Zhang, Y., Xiao, G., & Wang, W. (2022). Screening and Identification of Lassa Virus Entry Inhibitors from a Fragment-Based Drug Discovery Library. Viruses, 14(12), 2649. https://doi.org/10.3390/v14122649