

Metagenomic Analysis of Respiratory RNA Virome of Children with and without Severe Acute Respiratory Infection from the Free State, South Africa during COVID-19 Pandemic Reveals Higher Diversity and Abundance in Summer Compared with Winter Period

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Settings and Demography
2.2. Sampling and Sample Collection
2.3. Sample Pooling
2.4. Sample Processing, Enrichment and Extraction
2.5. Positive and Negative Controls for Evaluation of Viral Recovery and Workflow Contamination
2.6. Qubit Quantification of the Extracted RNA
2.7. DNase Treatment on Extracted RNA and Purification
2.8. Depletion of Ribosomal RNA (rRNA)
2.9. Reverse Transcription and Whole Transcriptome Amplification
2.10. Library Preparation and Next-Generation Sequencing
2.11. Bioinformatic Analysis
2.12. Phylogenetic Analysis
2.13. Statistical Analysis
3. Results
3.1. General Demographics and Clinical Presentations
3.2. Sequencing Data
3.3. Respiratory Tract Virome Analyses from Children with and without SARI during Summer and Winter
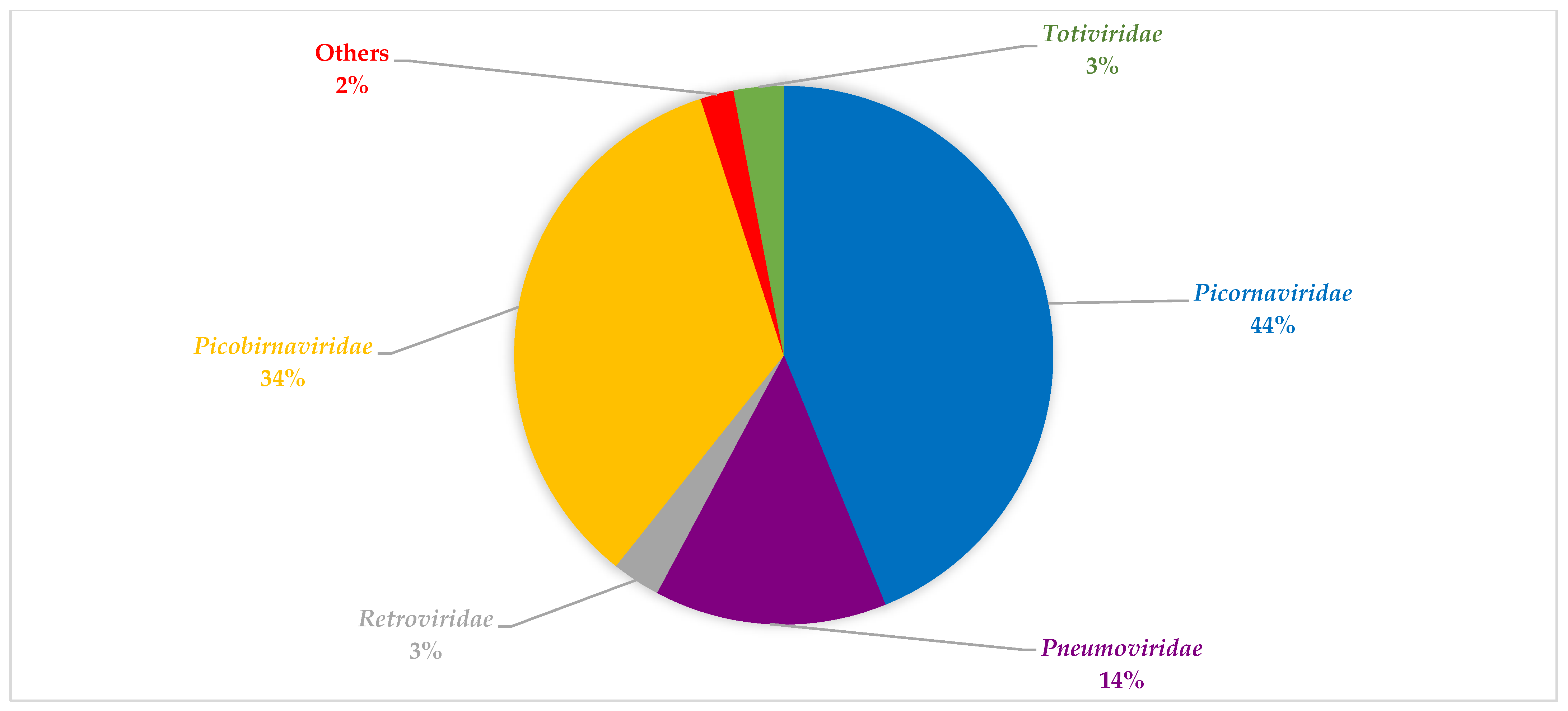
3.4. Viral Composition during the Seasons Investigated and between Both SARI and Non-SARI Group
3.5. Exclusive Viral Reads Detection in Pools
3.6. Viral Recovery and Evaluation for Contaminants in the mNGS Workflow
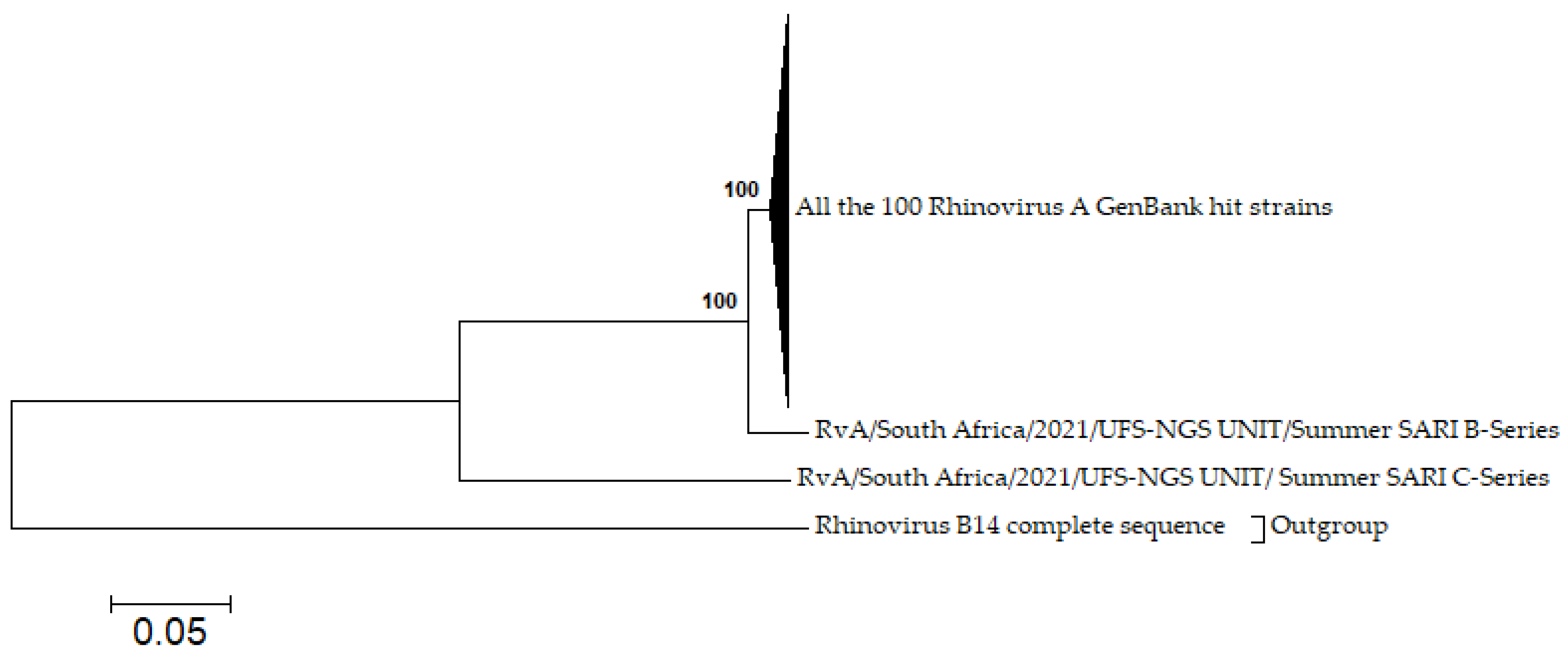
3.7. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gessner, B.D. Severe acute respiratory illness in Sub-Saharan Africa. J. Infect. Dis. 2015, 212, 843–844. [Google Scholar] [CrossRef] [PubMed]
- Ayeni, O.A.; Walaza, S.; Tempia, S.; Groome, M.; Kahn, K.; Madhi, S.A.; Cohen, A.L.; Moyes, J.; Venter, M.; Pretorius, M.; et al. Mortality in children aged <5 years with severe acute respiratory illness in a high HIV- prevalence urban and rural areas of South Africa, 2009–2013. PLoS ONE 2021, 16, e0255941. [Google Scholar] [CrossRef]
- Tang, J.W.T.; Lam, T.T.; Zaraket, H.; Lipkin, W.I.; Drews, S.J.; Hatchette, T.F.; Heraud, J.M.; Koopmans, M.P.; Abraham, A.M.; Baraket, A.; et al. Global epidemiology of non-influenza RNA respiratory viruses: Data gaps and a growing need for surveillance. Lancet Infect. Dis. 2017, 17, e320–e326. [Google Scholar] [CrossRef]
- Aman, A.T.; Wibawa, T.; Kosasih, H.; Asdie, R.H.; Safitri, I.; Intansari, U.S.; Mawarti, Y.; Sudarmono, P.; Arif, M.; Puspitasari, D.; et al. Etiologies of severe acute respiratory infection (SARI) and misdiagnosis of influenza in Indonesia, 2013–2016. Influenza Other Respir. Viruses 2021, 15, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Anders, K.L.; Nguyen, H.L.; Nguyen, N.M.; Van Thuy, N.T.; Hong Van, N.T.; Hieu, N.T.; Hong Tham, N.T.; Thanh Ha, P.T.; Lien, L.B.; Chau, N.V.V.; et al. Epidemiology and virology of acute respiratory infections during the first year of life: A birth cohort study in Vietnam. Pediatr. Infect. Dis. J. 2015, 34, 361–370. [Google Scholar] [CrossRef]
- Van Den Hoogen, B.G.; De Jong, J.C.; Groen, J.; Kuiken, T.; De Groot, R.; Fouchier, R.A.M.; Osterhaus, A.D.M.E. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat. Med. 2001, 7, 719–724. [Google Scholar] [CrossRef]
- Ksiazek, T.G.; Erdman, D.; Goldsmith, C.S.; Zaki, S.R.; Peret, T.; Emery, S.; Tong, S.; Urbani, C.; Comer, J.A.; Lim, W.; et al. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1953–1966. [Google Scholar] [CrossRef]
- Allander, T.; Tammi, M.T.; Eriksson, M.; Bjerkner, A.; Tiveljung-Lindell, A.; Andersson, B. Cloning of a human parvovirus by molecular screening of respiratory tract samples. Proc. Natl. Acad. Sci. USA 2005, 102, 12891–12896. [Google Scholar] [CrossRef]
- Zhou, Y.; Fernandez, S.; Yoon, I.K.; Simasathien, S.; Watanaveeradej, V.; Yang, Y.; Marte-Salcedo, O.A.; Shuck-Lee, D.J.; Thomas, S.J.; Hang, J.; et al. Metagenomics study of viral pathogens in undiagnosed respiratory specimens and identification of human enteroviruses at a Thailand hospital. Am. J. Trop. Med. Hyg. 2016, 95, 663–669. [Google Scholar] [CrossRef]
- Jartti, T.; Lehtinen, P.; Vuorinen, T.; Österback, R.; Van Den Hoogen, B.; Osterhaus, A.D.M.E.; Ruuskanen, O. Respiratory picornaviruses and respiratory syncytial virus as causative agents of acute expiratory wheezing in children. Emerg. Infect. Dis. 2004, 10, 1095–1101. [Google Scholar] [CrossRef]
- Young, N.S.; Brown, K.E. Parvovirus B19. N. Engl. J. Med. 2004, 350, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.; Taboada, B.; Yocupicio-Monroy, M.; Arias, C.F. Human virome. Arch. Med. Res. 2017, 48, 701–716. [Google Scholar] [CrossRef] [PubMed]
- Temerozo, J.R.; Fintelman-Rodrigues, N.; dos Santos, M.C.; Hottz, E.D.; Sacramento, C.Q.; de Paula Dias da Silva, A.; Mandacaru, S.C.; dos Santos Moraes, E.C.; Trugilho, M.R.O.; Gesto, J.S.M.; et al. Human endogenous retrovirus K in the respiratory tract is associated with COVID-19 physiopathology. Microbiome 2022, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, N.; Li, Y.; Lu, R.; Wang, H.; Liu, G.; Zou, X.; Xie, Z.; Tan, W. Metagenomic analysis of viral genetic diversity in respiratory samples from children with severe acute respiratory infection in China. Clin. Microbiol. Infect. 2016, 22, 458.e1–458.e9. [Google Scholar] [CrossRef]
- Louten, J. Virus transmission and epidemiology. In Essential Human Virology; Academic Press: Cambridge, MA, USA, 2016; pp. 71–92. [Google Scholar]
- Man, W.H.; Piters, W.A.A.D.S.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef]
- Tay, C.J.X.; Ta, L.D.H.; Ow Yeong, Y.X.; Yap, G.C.; Chu, J.J.H.; Lee, B.W.; Tham, E.H. Role of upper respiratory microbiota and virome in childhood rhinitis and wheeze: Collegium Internationale Allergologicum Update 2021. Int. Arch. Allergy Immunol. 2021, 182, 265–276. [Google Scholar] [CrossRef]
- Wylie, K.M. The virome of the human respiratory tract. Clin. Chest Med. 2017, 38, 11–19. [Google Scholar] [CrossRef]
- Romero-espinoza, J.A.; Moreno-valencia, Y.; Coronel-tellez, R.H.; Castillejos-lopez, M.; Hernandez, A.; Dominguez, A.; Miliar-garcia, A.; Barbachano-guerrero, A.; Perez-padilla, R.; Alejandre-garcia, A.; et al. Virome and bacteriome characterization of children with pneumonia and asthma in Mexico City during winter seasons 2014 and 2015. PLoS ONE 2018, 13, e0192878. [Google Scholar] [CrossRef]
- Wylie, K.M.; Mihindukulasuriya, K.A.; Sodergren, E.; Weinstock, G.M.; Storch, G.A. Sequence analysis of the human virome in Febrile and Afebrile children. PLoS ONE 2012, 7, e27735. [Google Scholar] [CrossRef]
- Famoroti, T.; Sibanda, W.; Ndung, T. Prevalence and seasonality of common viral respiratory pathogens, including Cytomegalovirus in children, between 0–5 years of age in KwaZulu-Natal, an HIV endemic province in South Africa. BMC Pediatr. 2018, 18, 240. [Google Scholar] [CrossRef]
- Khor, C.S.; Sam, I.C.; Hooi, P.S.; Quek, K.F.; Chan, Y.F. Epidemiology and seasonality of respiratory viral infections in hospitalized children in Kuala Lumpur, Malaysia: A retrospective study of 27 years. BMC Pediatr. 2012, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Yang, L.; Lou, C.T.; Yang, F.; SiTou, K.I.; Hu, H.; Io, K.; Cheok, K.T.; Pan, B.; Ung, C.O.L. Viral etiology and epidemiology of pediatric patients hospitalized for acute respiratory tract infections in Macao: A retrospective study from 2014 to 2017. BMC Infect. Dis. 2021, 21, 306. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.; Chen, J. Application of next generation sequencing for the detection of human viral pathogens in clinical specimens. J. Clin. Virol. 2017, 86, 20–26. [Google Scholar] [CrossRef] [PubMed]
- van Boheemen, S.; van Rijn, A.L.; Pappas, N.; Carbo, E.C.; Vorderman, R.H.P.; Sidorov, I. Retrospective validation of a metagenomic sequencing protocol for combined detection of RNA and DNA viruses using respiratory samples from pediatric patients. J. Mol. Diagn. 2020, 22, 196–207. [Google Scholar] [CrossRef]
- Rajagopala, S.V.; Bakhoum, N.G.; Pakala, S.B.; Shilts, M.H.; Rosas-Salazar, C.; Mai, A.; Boone, H.H.; McHenry, R.; Yooseph, S.; Halasa, N.; et al. Metatranscriptomics to characterize respiratory virome, microbiome, and host response directly from clinical samples. Cell Rep. Methods 2021, 1, 100091. [Google Scholar] [CrossRef]
- Lysholm, F.; Wetterbom, A.; Lindau, C.; Darban, H.; Bjerkner, A.; Fahlander, K.; Lindberg, A.M.; Persson, B.; Allander, T.; Andersson, B. Characterization of the viral microbiome in patients with severe lower respiratory tract infections, using metagenomic sequencing. PLoS ONE 2012, 7, e30875. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Ma, J.; Zhang, J.; Hu, Y.; Dong, W.; Wan, Z.; Li, Q.; Kuang, Y.Q.; Lan, K.; et al. Altered respiratory virome and serum cytokine profile associated with recurrent respiratory tract infections in children. Nat. Commun. 2019, 10, 228. [Google Scholar] [CrossRef]
- Fitzner, J.; Qasmieh, S.; Mounts, A.W.; Alexander, B.; Besselaar, T.; Briand, S.; Brown, C.; Clark, S.; Dueger, E.; Gross, D.; et al. Revision of clinical case definitions: Influenza-like illness and severe acute respiratory infection. Bull. World Health Organ. 2018, 96, 122–128. [Google Scholar] [CrossRef]
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.M.; et al. Genome Detective: An automated system for virus identification from high-throughput sequencing data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef]
- Cleemput, S.; Dumon, W.; Fonseca, V.; Karim, W.A.; Giovanetti, M.; Alcantara, L.C.; Deforche, K.; Oliveira, T. Genome detective coronavirus typing tool for rapid identification and characterization of novel coronavirus genomes. Bioinformatics 2020, 36, 3552–3555. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, D.W.; Schreiber, P.W.; Ruehe, B.; Zagordi, O.; Bayard, C.; Greiner, M.; Benden, C.; Geissberger, D.; Capaul, R.; Zbinden, A.; et al. Metagenomic sequencing complements routine diagnostics in identifying viral pathogens in lung transplant recipients with unknown etiology of respiratory infection. PLoS ONE 2017, 12, e0177340. [Google Scholar] [CrossRef] [PubMed]
- Moe, N.; Pedersen, B.; Nordbø, S.A.; Skanke, L.H.; Krokstad, S.; Smyrnaios, A.; Døllner, H. Respiratory virus detection and clinical diagnosis in children attending day care. PLoS ONE 2016, 11, e0159196. [Google Scholar] [CrossRef] [PubMed]
- Douros, K.; Kotzia, D.; Kottaridi, C.; Giotas, A.; Boutopoulou, B.; Bozas, E.; Matziou, V.; Priftis, K.; Papaevangelou, V. Evidence for respiratory viruses interactions in asymptomatic preschool-aged children. Allergol. Immunopathol. 2019, 47, 260–264. [Google Scholar] [CrossRef]
- Advani, S.; Sengupta, A.; Forman, M.; Valsamakis, A.; Milstone, A. Detecting respiratory viruses in asymptomatic children. Pediatr. Infect. Dis. J. 2012, 23, 1221–1226. [Google Scholar] [CrossRef]
- Walker, G.J.; Stelzer-Braid, S.; Shorter, C.; Honeywill, C.; Wynn, M.; Willenborg, C.; Barnes, P.; Kang, J.; Pierse, N.; Crane, J.; et al. Viruses associated with acute respiratory infection in a community-based cohort of healthy New Zealand children. J. Med. Virol. 2022, 94, 454–460. [Google Scholar] [CrossRef]
- Castro, I.A.; Costa, L.D.C.; Oliveira, A.C.R.; Souza, M.; das Dôres de Paula Cardoso, D.; Camargos, P.A.M.; Costa, P.S.S.; Fiaccadori, F.S. Circulation profile of respiratory viruses in symptomatic and asymptomatic children from Midwest Brazil. Brazilian J. Microbiol. 2020, 51, 1729–1735. [Google Scholar] [CrossRef]
- Sarna, M.; Lambert, S.B.; Sloots, T.P.; Whiley, D.M.; Alsaleh, A.; Mhango, L.; Bialasiewicz, S.; Wang, D.; Nissen, M.D.; Grimwood, K.; et al. Viruses causing lower respiratory symptoms in young children: Findings from the ORChID birth cohort. Thorax 2018, 73, 969–979. [Google Scholar] [CrossRef]
- Morikawa, S.; Hiroi, S.; Kase, T. Detection of respiratory viruses in gargle specimens of healthy children. J. Clin. Virol. 2015, 64, 59–63. [Google Scholar] [CrossRef]
- Twigg, H.L.; Knox, K.S.; Zhou, J.; Crothers, K.A.; Nelson, D.E.; Toh, E.; Day, R.B.; Lin, H.; Gao, X.; Dong, Q.; et al. Effect of advanced HIV infection on the respiratory microbiome. Am. J. Respir. Crit. Care Med. 2016, 194, 226–235. [Google Scholar] [CrossRef]
- Beck, J.M.; Schloss, P.D.; Venkataraman, A.; Twigg, H.; Jablonski, K.A.; Bushman, F.D.; Campbell, T.B.; Charlson, E.S.; Collman, R.G.; Crothers, K.; et al. Multicenter comparison of lung and oral microbiomes of HIV-infected and HIV-uninfected individuals. Am. J. Respir. Crit. Care Med. 2015, 192, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Monaco, C.L.; Gootenberg, D.B.; Zhao, G.; Handley, S.A.; Ghebremichael, M.S.; Lim, E.S.; Lankowski, A.; Baldridge, M.T.; Wilen, C.B.; Flagg, M.; et al. Altered virome and bacterial microbiome in human immunodeficiency virus-associated acquired immunodeficiency syndrome human immunodeficiency virus (HIV) infection is associated with increased intestinal translocation of microbial products and enteropathy as well as alterations in gut bacterial. Cell Host Microbe 2016, 19, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Moyes, D.L.; Martin, A.; Sawcer, S.; Temperton, N.; Worthington, J.; Griffiths, D.J.; Venables, P.J. The distribution of the endogenous retroviruses HERV-K113 and HERV-K115 in health and disease. Genomics 2005, 86, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Johanning, G.L.; Malouf, G.G.; Zheng, X.; Esteva, F.J.; Weinstein, J.N.; Wang-Johanning, F.; Su, X. Expression of human endogenous retrovirus-K is strongly associated with the basal-like breast cancer phenotype. Sci. Rep. 2017, 7, 41960. [Google Scholar] [CrossRef] [PubMed]
- Boller, K.; Schönfeld, K.; Lischer, S.; Fischer, N.; Hoffmann, A.; Kurth, R.; Tönjes, R.R. Human endogenous retrovirus HERV-K113 is capable of producing intact viral particles. J. Gen. Virol. 2008, 89, 567–572. [Google Scholar] [CrossRef]
- Ferravante, C.; Sanna, G.; Melone, V.; Fromentier, A.; Rocco, T.; D’Agostino, Y.; Lamberti, J.; Alexandrova, E.; Pecoraro, G.; Pagliano, P.; et al. Nasopharyngeal virome analysis of COVID-19 patients during three different waves in Campania region of Italy. J. Med. Virol. 2022, 94, 2275–2283. [Google Scholar] [CrossRef]
- Pretorius, M.A.; Tempia, S.; Treurnicht, F.K.; Walaza, S.; Cohen, A.L.; Moyes, J.; Hellferscee, O.; Variava, E.; Dawood, H.; Chhagan, M.; et al. Genetic diversity and molecular epidemiology of human rhinoviruses in South Africa. Influenza Other Respi. Viruses 2014, 8, 567–573. [Google Scholar] [CrossRef]
- Jiang, H.; Yang, T.; Yang, C.; Lu, Y.; Yi, Z.; Zhang, Q.; Wang, W. Molecular epidemiology and clinical characterization of human rhinoviruses circulating in Shanghai, 2012–2020. Arch. Virol. 2022, 167, 1111–1123. [Google Scholar] [CrossRef]
- Moriyama, M.; Hugentobler, W.J.; Iwasaki, A. Seasonality of respiratory viral infections. Annu. Rev. Virol. 2020, 7, 83–101. [Google Scholar] [CrossRef]
- Toivonen, L.; Schuez-Havupalo, L.; Karppinen, S.; Teros-Jaakkola, T.; Rulli, M.; Mertsola, J.; Waris, M.; Peltola, V. Rhinovirus infections in the first 2 years of life. Pediatrics 2016, 138, e20161309. [Google Scholar] [CrossRef]
- Oh, D.Y.; Buda, S.; Biere, B.; Reiche, J.; Schlosser, F.; Duwe, S.; Wedde, M.; von Kleist, M.; Mielke, M.; Wolff, T.; et al. Trends in respiratory virus circulation following COVID-19-targeted nonpharmaceutical interventions in Germany, January–September 2020: Analysis of national surveillance data. Lancet Reg. Health-Eur. 2021, 6, 100112. [Google Scholar] [CrossRef] [PubMed]
- Varela, F.H.; Sartor, I.T.S.; Polese-Bonatto, M.; Azevedo, T.R.; Kern, L.B.; Fazolo, T.; de David, C.N.; Zavaglia, G.O.; Fernandes, I.R.; Krauser, J.R.M.; et al. Rhinovirus as the main co-circulating virus during the COVID-19 pandemic in children. J. Pediatr. 2022, 98, 579–586. [Google Scholar] [CrossRef]
- Tempia, S.; Walaza, S.; Bhiman, J.N.; McMorrow, M.L.; Moyes, J.; Mkhencele, T.; Meiring, S.; Quan, V.; Bishop, K.; McAnerney, J.M.; et al. Decline of influenza and respiratory syncytial virus detection in facility-based surveillance during the COVID-19 pandemic, South Africa, January to October 2020. Eurosurveillance 2021, 26, 2001600. [Google Scholar] [CrossRef] [PubMed]
- Groves, H.E.; Papenburg, J.; Mehta, K.; Bettinger, J.A.; Sadarangani, M.; Halperin, S.A.; Morris, S.K.; Bancej, C.; Burton, C.; Embree, J.; et al. The effect of the COVID-19 pandemic on influenza-related hospitalization, intensive care admission and mortality in children in Canada: A population-based study. Lancet Reg. Health-Am. 2022, 7, 100132. [Google Scholar] [CrossRef] [PubMed]
- Bents, S.; Viboud, C.; Grenfell, B.; Hogan, A.; Tempia, S.; von Gottberg, A.; Moyes, J.; Walaza, S.; Cohen, C.; Baker, R. The impact of COVID-19 non-pharmaceutical interventions on future respiratory syncytial virus transmission in South Africa. medRxiv 2022, 12, 22271872. [Google Scholar] [CrossRef]
- Ogunbayo, A.E.; Mogotsi, M.T.; Sondlane, H.; Nkwadipo, K.R.; Sabiu, S.; Nyaga, M.M. Pathogen profile of children hospitalised with severe acute respiratory infections during COVID-19 pandemic in the Free State Province, South Africa. Int. J. Environ. Res. Public Health 2022, 19, 10418. [Google Scholar] [CrossRef] [PubMed]
- Babiker, A.; Bradley, H.L.; Stittleburg, V.D.; Ingersoll, J.M.; Key, A.; Kraft, C.S.; Waggoner, J.J.; Piantadosi, A. Metagenomic sequencing to detect respiratory viruses in persons under investigation for COVID-19. J. Clin. Microbiol. 2021, 59, e02142-20. [Google Scholar] [CrossRef]
- Mostafa, H.H.; Fissel, J.A.; Fanelli, B.; Bergman, Y.; Gniazdowski, V.; Dadlani, M.; Carroll, K.C.; Colwell, R.R.; Simner, P.J. Metagenomic next-generation sequencing of nasopharyngeal specimens collected from confirmed and suspect covid-19 patients. mBio 2020, 11, e01969-20. [Google Scholar] [CrossRef]
- de Vries, J.J.C.; Brown, J.R.; Fischer, N.; Sidorov, I.A.; Morfopoulou, S.; Huang, J.; Munnink, B.B.O.; Sayiner, A.; Bulgurcu, A.; Rodriguez, C.; et al. Benchmark of thirteen bioinformatic pipelines for metagenomic virus diagnostics using datasets from clinical samples. J. Clin. Virol. 2021, 141, 104908. [Google Scholar] [CrossRef]
- Cloete, J.; Kruger, A.; Masha, M.; du Plesis, N.M.; Mawela, D.; Tshukudu, M.; Manyane, T.; Komane, L.; Venter, M.; Jassat, W.; et al. Paediatric hospitalisations due to COVID-19 during the first SARS-CoV-2 omicron (B.1.1.529) variant wave in South Africa: A multicentre observational study. Lancet Child Adolesc. Health 2022, 6, 294–302. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pooled Samples | Number of Samples | Reads after Quality Filtering | % of Viral Reads |
|---|---|---|---|
| SARI Summer | |||
| SARI Pool A1 | 3 | 1,079,760 | 2 |
| SARI Pool B1 | 8 | 1,703,176 | 3 |
| SARI Pool C1 | 23 | 8,500,586 | 4 |
| SARI Pool D1 | 55 | 1,341,384 | 5 |
| SARI Pool E1 | 16 | 862,754 | 11 |
| Non-SARI Summer | |||
| Non-SARI Pool A2 | 6 | 140,444 | 5 |
| Non-SARI Pool B2 | 16 | 312,496 | 12 |
| Non-SARI Pool C2 | 38 | 366,146 | 4 |
| Non-SARI Pool D2 | 20 | 1,250,024 | 6 |
| SARI Winter | |||
| SARI Pool E1 | 15 | 293,394 | 2 |
| SARI Pool F1 | 14 | 166,726 | 2 |
| SARI Pool G1 | 15 | 272,596 | 3 |
| Non-SARI Winter | |||
| Non-SARI Pool E2 | 20 | 15,434 | 5 |
| Non-SARI Pool F2 | 20 | 13,308 | 6 |
| Non-SARI Pool G2 | 19 | 37,010 | 9 |
| Family | Viral Species/Read Number | Genome Structure | A1 | B1 | C1 | D1 | E1 |
|---|---|---|---|---|---|---|---|
| Picornaviridae | HRV-A (21,328) | ssRNA | 342/16.2 | 15,441/99.8 | 5495/99.2 | 22/13.6 | 28/3.4 |
| HRV-C (9972) | ssRNA | 1122/53.9 | 2315/25.2 | 31/8.2 | 264/25.8 | 6240/89.5 | |
| Parechovirus (236) | ssRNA | 176/27.8 | 60/8 | ||||
| Enterovirus J (60) | ssRNA | 60/7.9 | |||||
| HRV-B (261) | ssRNA | 159/28.3 | 102/4.1 | ||||
| Enterovirus A (44) | ssRNA | 44/8.4 | |||||
| Enterovirus B (2523) | ssRNA | 2523/82.8 | |||||
| Enterovirus C (13) | ssRNA | ||||||
| Enterovirus A114 (101) | ssRNA | 24/3.4 | 77/30.1 | 13/3 | |||
| Partitiviridae | Fig cryptic virus (420) | dsRNA | 420/78.1 | ||||
| Pneumoviridae | RSV-B (2333) | ssRNA | 234/23 | 1840/89.9 | 493/48 | ||
| RSV-A (572) | ssRNA | 124/19 | 522/40.5 | 50/7.6 | |||
| Totiviridae | Scheffersomyces segobiensis virus L (7892) | dsRNA | 152/49.4 | 271/45.1 | 1395/46.3 | 3110/84.4 | 2964/80.3 |
| Diatom colony-associated dsRNA virus 3 (27) | dsRNA | 27/5.2 | |||||
| Diatom colony-associated dsRNA virus 10 (21) | dsRNA | 17/7.3 | 4/4.5 | ||||
| Red clover powdery mildew-associated totivirus 2 (218) | dsRNA | 198/41.5 | 20/3.5 | ||||
| Xanthophyllomyces dendrorhous virus L1B (82) | dsRNA | 82/16.9 | |||||
| Saccharomyces cerevisiae virus (52) | dsRNA | 52/19.2 | |||||
| Maize-associated totivirus 2 (1346) | dsRNA | 1346/11.8 | |||||
| Red clover powdery mildew-associated totivirus 6 (3) | dsRNA | 3/3.5 | |||||
| Picobirnaviridae | Otarine Picobirnavirus (segment 2) (49048) | dsRNA | 18/52 | 49,030/98.4 | |||
| Picobirnavirus green monkey/KNA/2015 (31) | dsRNA | 6/24.9 | 25/6.9 | ||||
| Picobirnavirus dog/KNA/2015 (125) | dsRNA | 125/29 | |||||
| Human Picobirnavirus (75) | dsRNA | 75/12.5 | |||||
| Coronaviridae | SARS-CoV-2 (2) | 2/1 | |||||
| Retroviridae | Human endogenous retrovirus K113 (1287) | ssRNA-RT | 208/17.2 | 109/16.8 | 107/16.1 | 784/22.5 | 79/20 |
| Moloney murine leukaemia virus (1528) | ssRNA | 120/13.7 | 1404/22.4 | 4/7 | |||
| Equine infectious anaemia virus (651) | ssRNA | 28/4.8 | 107/16.1 | 404/20 | 112/19 | ||
| Koala retrovirus (2) | ssRNA-RT | 2/4.3 | |||||
| Reticuloendotheliosis virus (489) | ssRNA-RT | 61/9.6 | 428/13.2 | ||||
| RD114 Retrovirus (13) | ssRNA-RT | 13/5.8 | |||||
| Chick syncytial virus (3) | ssRNA-RT | 3/23.9 | |||||
| Atlantic salmon swim bladder sarcoma virus (79) | ssRNA-RT | 79/8.4 | |||||
| Feline leukaemia virus (82) | ssRNA | 70/7 | 12/4.7 | ||||
| Baboon endogenous virus strain M7 ssRNA (14) | ssRNA-RT | 14/4.5 | |||||
| Friend murine leukaemia virus (31) | ssRNA | 31/5.1 | |||||
| Gibbon ape leukaemia virus (14) | ssRNA | 14/3.7 | |||||
| Bovine retrovirus CH15 (9) | ssRNA-RT | 9/8.7 | |||||
| Chrysoviridae | Penicillium chrysogenum virus segment 3 (2) | dsRNA | 2/2.7 | ||||
| Virgaviridae | Tobacco Mosaic Virus (60) | ssRNA | 60/10.2 | ||||
| Pepper mild mottle virus (3) | ssRNA | 3/1.1 | |||||
| Betaflexiviridae | Apple chlorotic leafspot virus (622) | ssRNA | 527/41.4 | 95/18.4 | |||
| Apple stem grooving virus (33) | ssRNA | 33/6.6 | |||||
| Herpesviridae | Saimiriine gammaherpesvirus 2 (2) | dsDNA | 2/0.3 | ||||
| Phycodnaviridae | Micromonas pusilla virus 12T (26) | dsDNA | 26/0.1 | ||||
| Endornaviridae | Grapevine endophyte alphaendornavirus (94) | ssRNA | 94/10.2 | ||||
| Bell pepper alphaendornavirus (4) | ssRNA | 4/2.4 | |||||
| Circoviridae | Porcine stool-associated circular virus (16) | ssDNA | 16/19 | ||||
| Unclassified | Bovine serum-associated circular virus (331) | Unknown | 28/99.2 | 303/99.8 |
| Family | Viral Species/Read Number | Genome Structure | E1 | F1 | G1 |
|---|---|---|---|---|---|
| Picornaviridae | HRV-C (231) | ssRNA | 130/30.9 | 69/26.5 | 32/7.5 |
| HRV-A (234) | ssRNA | 6/3.5 | 26/13 | 206/52 | |
| Enterovirus B (126) | ssRNA | 31/11.4 | 95/30.7 | ||
| Pneumoviridae | RSV-B (186) | ssRNA | 126/18.7 | 15/5.5 | 145/16 |
| RSV-A (1) | ssRNA | 1/0.7 | |||
| Retroviridae | Human endogenous retrovirus K113 (39) | ssRNA-RT | 11/1.8 | 28/8.4 | |
| Picobirnaviridae | Otarine Picobirnavirus (segment 2) (9) | dsRNA | 9/15.3 | ||
| Picobirnavirus green monkey/KNA/2015 (325) | dsRNA | 325/82 | |||
| Chicken Picobirnavirus (segment RNA 1) (126) | dsRNA | 126/17.5 | |||
| Endornaviridae | BPA (6) | ssRNA | 5/0.6 | 1/0.4 | |
| Coronaviridae | Human coronavirus NL63 (3) | ssRNA | 3/0.8 | ||
| Virgaviridae | Tobacco mosaic virus (6) | ssRNA | 6/5.4 | ||
| Retroviridae | Equine infectious anaemia virus (4) | ssRNA | 2/3.7 | 2/1.8 | |
| Totiviridae | Scheffersomyces segobiensis virus L (40) | dsRNA | 40/10.2 | ||
| Tombusviridae | Bermuda grass latent virus (8) | ssRNA | 8/4.5 |
| Family | Viral Species/Read Number | Genome Structure | A2 | B2 | C2 | D2 |
|---|---|---|---|---|---|---|
| Picornaviridae | HRV-C (7926) | ssRNA | 1388/85.5 | 6495/93.3 | 31/8.2 | 12/1.2 |
| HRV-A (29420) | ssRNA | 726/53.1 | 27923/92.9 | 746/86 | 25/4.2 | |
| HRV-B (347) | ssRNA | 347/65.8 | ||||
| Enterovirus J (71) | ssRNA | 60/7.9 | 11/1.0 | |||
| Enterovirus B (513) | ssRNA | 513/51.2 | ||||
| Enterovirus A (17) | ssRNA | 17/9 | ||||
| Totiviridae | Scheffersomyces segobiensis virus L (2204) | dsRNA | 785/82.6 | 1298/70.2 | 105/31 | 16/11.8 |
| Red clover powdery mildew-associated totivirus 7 (41) | dsRNA | 4/12 | 37/3.8 | |||
| Diatom colony-associated dsRNA virus 3 (27) | dsRNA | 27/5.2 | ||||
| Diatom colony-associated dsRNA virus 16 (17) | dsRNA | 17/7.3 | ||||
| Paramoxyviridae | HPIV-2 (384) | ssRNA | 384/34.5 | |||
| HPIV-3 (305) | ssRNA | 295/35.7 | 10/4.7 | |||
| Coronaviridae | Human coronavirus NL63 (223) | ssRNA | 223/10.7 | |||
| Retroviridae | Feline leukaemia virus (4) | ssRNA | 4/6.1 | |||
| Human endogenous retrovirus K113 (126) | ssRNA-RT | 60/8.1 | 7/1.5 | 59/17.8 | ||
| Equine infectious anaemia virus (86) | ssRNA | 40/7.9 | 46/5.4 | |||
| Reticuloendotheliosis virus (136) | ssRNA | 75/8.5 | 61/9.6 | |||
| RD114 Retrovirus (13) | ssRNA | 13/5.7 | ||||
| Partitiviridae | Ustilaginoidea virens partitivirus 2 (segment RNA 1) (249) | dsRNA | 249/16.5 | |||
| Fusarium poae virus 1 (segment 1) (14) | dsRNA | 14/7 | ||||
| Chrysoviridae | Penicillium chrysogenum virus segment 2 (6) | dsRNA | 6/9.25 | |||
| Picobirnaviridae | Otarine picobirnavirus (segment 2) (1551) | dsRNA | 1514/69 | 37/30.5 | ||
| Endornaviridae | Phaseolus vulgaris alphaendornavirus 1 (174) | ssRNA | 174/32.9 | |||
| Phaseolus vulgaris alphaendornavirus 2 (93) | ssRNA | 93/18.7 | ||||
| Herpesviridae | Saimiriine gamma herpesvirus 2 (2) | dsDNA | 2/0.25 |
| Family | Viral Genus/Read Number | Genome Structure | E2 | F2 | G2 |
|---|---|---|---|---|---|
| Retroviridae | Equine infectious anaemia virus (209) | ssRNA | 36/11.2 | 173/16.3 | |
| Paramoxyviridae | HPIV-3 (9) | ssRNA | 9/1.0 | ||
| Bromoviridae | Ageratum latent virus (82) | ssRNA | 82/18.3 | ||
| Parietaria mottle virus (segment RNA 2) (63) | ssRNA | 63/11.9 | |||
| Reoviridae | Rotavirus A (segment 1) (17) | dsRNA | 17/13.3 | ||
| Rotavirus A (segment 2) (45) | dsRNA | 45/40.9 | |||
| Rotavirus A (segment 6) (5) | dsRNA | 5/16.7 | |||
| Bat Rotavirus (segment 4) (15) | dsRNA | 15/19 | |||
| Rotavirus A (segment 3) (6) | dsRNA | 6/11 | |||
| Picornaviridae | HRV-C (1) | ssRNA | 1//1.9 | ||
| HRV-A (17) | ssRNA | 17/2.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogunbayo, A.E.; Mogotsi, M.T.; Sondlane, H.; Nkwadipo, K.R.; Sabiu, S.; Nyaga, M.M. Metagenomic Analysis of Respiratory RNA Virome of Children with and without Severe Acute Respiratory Infection from the Free State, South Africa during COVID-19 Pandemic Reveals Higher Diversity and Abundance in Summer Compared with Winter Period. Viruses 2022, 14, 2516. https://doi.org/10.3390/v14112516
Ogunbayo AE, Mogotsi MT, Sondlane H, Nkwadipo KR, Sabiu S, Nyaga MM. Metagenomic Analysis of Respiratory RNA Virome of Children with and without Severe Acute Respiratory Infection from the Free State, South Africa during COVID-19 Pandemic Reveals Higher Diversity and Abundance in Summer Compared with Winter Period. Viruses. 2022; 14(11):2516. https://doi.org/10.3390/v14112516
Chicago/Turabian StyleOgunbayo, Ayodeji E., Milton T. Mogotsi, Hlengiwe Sondlane, Kelebogile R. Nkwadipo, Saheed Sabiu, and Martin M. Nyaga. 2022. "Metagenomic Analysis of Respiratory RNA Virome of Children with and without Severe Acute Respiratory Infection from the Free State, South Africa during COVID-19 Pandemic Reveals Higher Diversity and Abundance in Summer Compared with Winter Period" Viruses 14, no. 11: 2516. https://doi.org/10.3390/v14112516
APA StyleOgunbayo, A. E., Mogotsi, M. T., Sondlane, H., Nkwadipo, K. R., Sabiu, S., & Nyaga, M. M. (2022). Metagenomic Analysis of Respiratory RNA Virome of Children with and without Severe Acute Respiratory Infection from the Free State, South Africa during COVID-19 Pandemic Reveals Higher Diversity and Abundance in Summer Compared with Winter Period. Viruses, 14(11), 2516. https://doi.org/10.3390/v14112516