

Gene Expression Analysis of Adapted Insect Cells during Influenza VLP Production Using RNA-Sequencing

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Media
2.2. Baculovirus Amplification and Storage
2.3. Production of Influenza HA-VLPs
2.4. Analytics
2.4.1. Cell Concentration and Viability
2.4.2. Metabolites Concentration
2.4.3. Baculovirus Titration
2.4.4. Western Blot
2.5. RNA Sequencing and Data Analysis
2.5.1. RNA Isolation and Library Preparation
2.5.2. Data Processing, Alignment and Counting
2.5.3. Differential Expression Analysis
2.5.4. Functional Annotation and Pathway Enrichment Analysis
3. Results
3.1. Production of Influenza HA-VLPs Using Adapted Insect High Five Cells
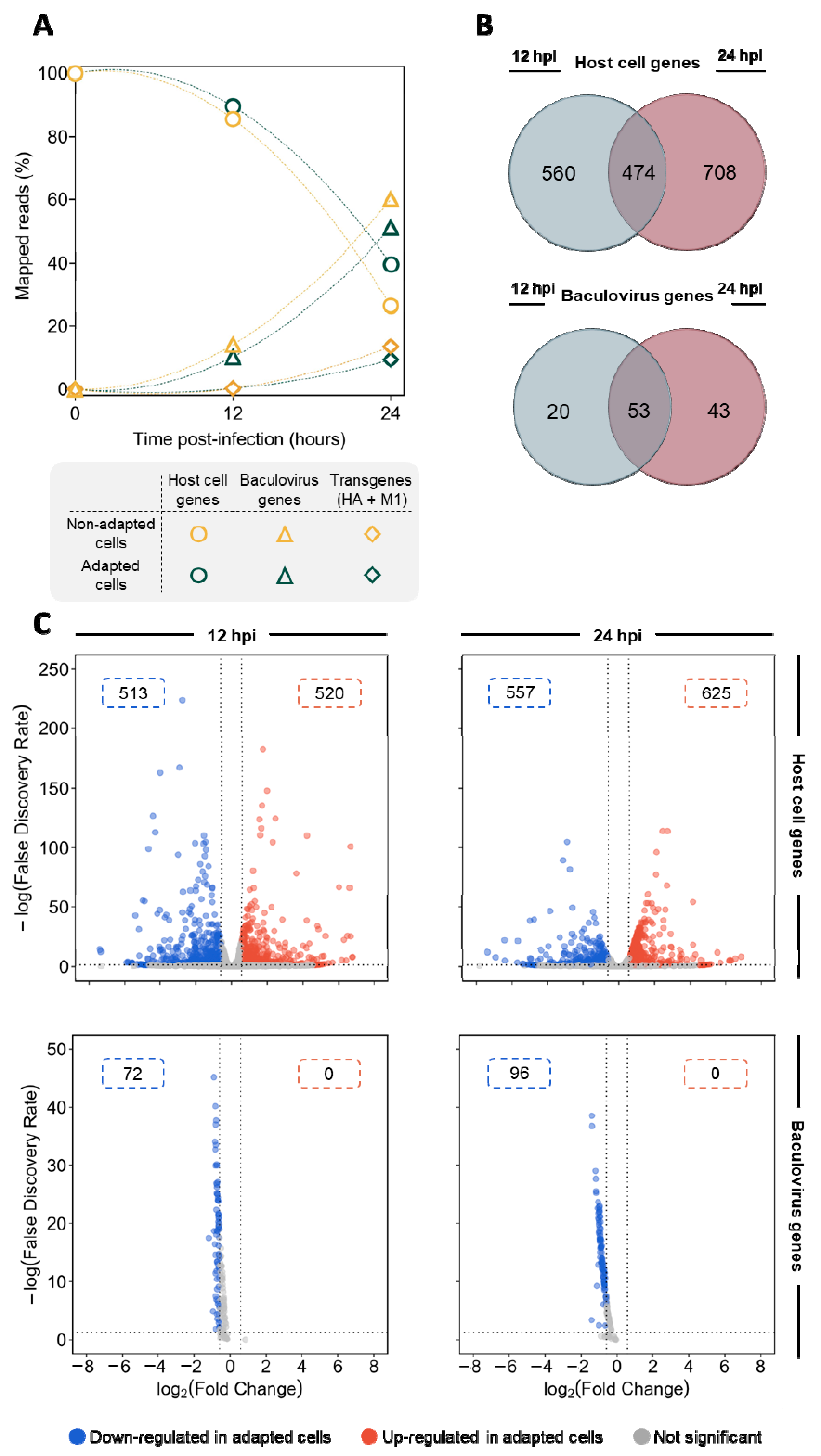
3.2. Gene Expression Profiling of Infected Adapted vs. Non-Adapted Cells
3.3. Pathway Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Oers, M.M.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus–insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Kost, T.A.; Kemp, C.W. Fundamentals of Baculovirus Expression and Applications. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2016; Volume 896, pp. 187–197. [Google Scholar] [CrossRef]
- Monteiro, F.; Carinhas, N.; Carrondo, M.J.T.; Bernal, V.; Alves, P.M. Toward System-Level Understanding of Baculovirus–Host Cell Interactions: From Molecular Fundamental Studies to Large-Scale Proteomics Approaches. In Frontiers in Microbiology; Frontiers Research Foundation: Lausanne, Switzerland, 2012; Volume 3, p. 391. [Google Scholar] [CrossRef]
- Abaandou, L.; Quan, D.; Shiloach, J. Affecting HEK293 Cell Growth and Production Performance by Modifying the Expression of Specific Genes. Cells 2021, 10, 1667. [Google Scholar] [CrossRef]
- Hong, J.K.; Lakshmanan, M.; Goudar, C.; Lee, D.-Y. Towards next generation CHO cell line development and engineering by systems approaches. Curr. Opin. Chem. Eng. 2018, 22, 1–10. [Google Scholar] [CrossRef]
- Babar, M.M.; Afzaal, H.; Pothineni, V.R.; Zaidi, N.-U.S.; Ali, Z.; Zahid, M.A.; Gul, A. Omics Approaches in Industrial Biotechnology and Bioprocess Engineering. In Omics Technologies and Bio-Engineering: Towards Improving Quality of Life; Elsevier: Amsterdam, The Netherlands, 2018; Volume 2, pp. 251–269. [Google Scholar] [CrossRef]
- Chen, Y.-R.; Zhong, S.; Fei, Z.; Hashimoto, Y.; Xiang, J.Z.; Zhang, S.; Blissard, G.W. The Transcriptome of the Baculovirus Autographa californica Multiple Nucleopolyhedrovirus in Trichoplusia ni Cells. J. Virol. 2013, 87, 6391–6405. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-R.; Zhong, S.; Fei, Z.; Gao, S.; Zhang, S.; Li, Z.; Wang, P.; Blissard, G.W. Transcriptome Responses of the Host Trichoplusia ni to Infection by the Baculovirus Autographa californica Multiple Nucleopolyhedrovirus. J. Virol. 2014, 88, 13781–13797. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Thiem, S.M. Responses of insect cells to baculovirus infection: Protein synthesis shutdown and apoptosis. J. Virol. 1997, 71, 7866–7872. [Google Scholar] [CrossRef] [PubMed]
- Koczka, K.; Peters, P.; Ernst, W.; Himmelbauer, H.; Nika, L.; Grabherr, R. Comparative transcriptome analysis of a Trichoplusia ni cell line reveals distinct host responses to intracellular and secreted protein products expressed by recombinant baculoviruses. J. Biotechnol. 2018, 270, 61–69. [Google Scholar] [CrossRef]
- Martínez-Solís, M.; Herrero, S.; Targovnik, A.M. Engineering of the baculovirus expression system for optimized protein production. In Applied Microbiology and Biotechnology; Springer: Berlin/Heidelberg, Germany, 2018; Volume 103, pp. 113–123. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Huang, Z.; Wen, W.; Wu, A.; Wang, C.; Niu, L. Enhancing Protein Expression in HEK-293 Cells by Lowering Culture Temperature. PLoS ONE 2015, 10, e0123562. [Google Scholar] [CrossRef]
- Fernandes, B.; Vidigal, J.; Correia, R.; Carrondo, M.J.; Alves, P.M.; Teixeira, A.P.; Roldão, A. Adaptive laboratory evolution of stable insect cell lines for improved HIV-Gag VLPs production. J. Biotechnol. 2020, 307, 139–147. [Google Scholar] [CrossRef]
- Correia, R.; Fernandes, B.; Alves, P.M.; Carrondo, M.J.; Roldão, A. Improving Influenza HA-Vlps Production in Insect High Five Cells via Adaptive Laboratory Evolution. Vaccines 2020, 8, 589. [Google Scholar] [CrossRef] [PubMed]
- Vieira, H.; Estêvão, C.; Roldao, A.; Peixoto, C.; Sousa, M.; Cruz, P.; Carrondo, M.; Alves, P. Triple layered rotavirus VLP production: Kinetics of vector replication, mRNA stability and recombinant protein production. J. Biotechnol. 2005, 120, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Tennant, J.R. Evaluation of The Trypan Blue Technique for Determination of Cell Viability. Transplantation 1964, 2, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Mena, J.A.; Ramírez, O.T.; Palomares, L.A. Titration of Non-Occluded Baculovirus Using a Cell Viability Assay. BioTechniques 2003, 34, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Roldão, A.; Oliveira, R.; Carrondo, M.J.; Alves, P.M. Error assessment in recombinant baculovirus titration: Evaluation of different methods. J. Virol. Methods 2009, 159, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, Y.; Zhang, H.; Farley, G.; Wang, J.; Quarles, A.K.; Weng, Z.; Zamore, P.D. The genome of the Hi5 germ cell line from Trichoplusia ni, an agricultural pest and novel model for small RNA biology. eLife 2018, 7, e04147. [Google Scholar] [CrossRef]
- Ayres, M.D.; Howard, S.C.; Kuzio, J.; Lopez-Ferber, M.; Possee, R.D. The Complete DNA Sequence of Autographa californica Nuclear Polyhedrosis Virus. Virology 1994, 202, 586–605. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Putri, G.H.; Anders, S.; Pyl, P.T.; Pimanda, E.J.; Zanini, F. Analysing high-throughput sequencing data in Python with HTSeq 2.0. Bioinformatics 2022, 38, 2943–2945. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Smyth, G.K. Small-sample estimation of negative binomial dispersion, with applications to SAGE data. Biostatistics 2007, 9, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Götz, S.; Garcia-Gomez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acid Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Benjaminit, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar]
- Drugmand, J.-C.; Schneider, Y.-J.; Agathos, S.N. Insect cells as factories for biomanufacturing. Biotechnol. Adv. 2012, 30, 1140–1157. [Google Scholar] [CrossRef]
- Goodwin, R.H. Replacement of vertebrate serum with lipids and other factors in the culture of invertebrate cells, tissues, parasites, and pathogens. In Source. Vitr. Cell. Dev. Biol. 1991, 27, 470–478. [Google Scholar] [CrossRef]
- Chan, R.B.; Tanner, L.; Wenk, M.R. Implications for lipids during replication of enveloped viruses. Chem. Phys. Lipids 2010, 163, 449–459. [Google Scholar] [CrossRef]
- Monteiro, F.; Bernal, V.; Chaillet, M.; Berger, I.; Alves, P.M. Targeted supplementation design for improved production and quality of enveloped viral particles in insect cell-baculovirus expression system. J. Biotechnol. 2016, 233, 34–41. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nayyar, N.; Kaur, I.; Malhotra, P.; Bhatnagar, R.K. Quantitative proteomics of Sf21 cells during Baculovirus infection reveals progressive host proteome changes and its regulation by viral miRNA. Sci. Rep. 2017, 7, 879. [Google Scholar] [CrossRef] [PubMed]
- Kalthoff, S.; Ehmer, U.; Freiberg, N.; Manns, M.P.; Strassburg, C.P. Interaction between Oxidative Stress Sensor Nrf2 and Xenobiotic-activated Aryl Hydrocarbon Receptor in the Regulation of the Human Phase II Detoxifying UDP-glucuronosyltransferase 1A10. J. Biol. Chem. 2010, 285, 5993–6002. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Sun, R.; Veeran, S.; Shu, B.; Yuan, H.; Zhong, G. Combined transcriptomic and proteomic analysis of harmine on Spodoptera frugiperda Sf9 cells to reveal the potential resistance mechanism. J. Proteom. 2019, 211, 103573. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.; Yang, Q.; Jiang, L.; Li, Q.; Xiao, Y.; Ye, M.; Xia, Q. RNA-Seq Analyses for Two Silkworm Strains Reveals Insight into Their Susceptibility and Resistance to Beauveria bassiana Infection. Int. J. Mol. Sci. 2017, 18, 234. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.T.; Imler, J.-L. The diversity of insect antiviral immunity: Insights from viruses. In Current Opinion in Microbiology; Elsevier: Amtserdam, The Netherlands, 2016; Volume 32, pp. 71–76. [Google Scholar] [CrossRef]
- Kataoka, C.; Kaname, Y.; Taguwa, S.; Abe, T.; Fukuhara, T.; Tani, H.; Moriishi, K.; Matsuura, Y. Baculovirus GP64-Mediated Entry into Mammalian Cells. J. Virol. 2012, 86, 2610–2620. [Google Scholar] [CrossRef] [PubMed]
- Blissard, G.W.; Wenz, J.R.; Thompson, B. Baculovirus gp64 Envelope Glycoprotein Is Sufficient to Mediate pH-Dependent Membrane Fusion. J. Virol. 1992, 66, 6829–6835. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, Y.; Ning, Y.-J.; Deng, F.; Vlak, J.M.; Hu, Z.; Wang, H.; Wang, M. The Major Hurdle for Effective Baculovirus Transduction into Mammalian Cells Is Passing Early Endosomes. J. Virol. 2019, 93, e00709-19. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | hpi | FC | logCPM | Biological Process (P) or Molecular Function (M) |
|---|---|---|---|---|---|
| LOC113500835 | xaa-Pro aminopeptidase 1 | 12 | 273.6 | 3.1 | hydrolase activity (M) |
| 24 | 756.1 | 1.6 | |||
| LOC113493104 | potassium channel subfamily K member 18-like | 12 | 102.1 | 1.7 | transmembrane transport (P) |
| 24 | 433.4 | 0.8 | |||
| LOC113505620 | serine protease HP21 precursor | 12 | 104.1 | −0.1 | proteolysis (P) |
| 24 | 116.2 | −0.9 | |||
| LOC113496461 | lipase member H-like | 12 | 111.6 | −1.6 | lipid metabolic process (P) |
| 24 | 79.4 | −1.3 | |||
| LOC113506781 | organic cation transporter protein-like | 12 | 91.9 | −0.3 | transmembrane transport (P) |
| 24 | 92.8 | −1.1 | |||
| LOC113504337 | GATA zinc finger domain-containing protein 4-like | 12 | 79.8 | −0.5 | regulation of transcription (P) |
| 24 | 74.3 | −1.3 | |||
| LOC113500515 | alkylglycerol monooxygenase-like | 12 | 97.8 | 1.0 | lipid metabolic process (P) |
| 24 | 46.6 | −0.3 | |||
| LOC113501396 | transcription factor glial cells missing 2-like | 12 | 108.6 | −1.7 | regulation of transcription (P) |
| 24 | 29.1 | −2.2 | |||
| LOC113501938 | monocarboxylate transporter 1-like | 12 | 86.2 | −1.8 | transmembrane transport (P) |
| 24 | 36.5 | −1.9 | |||
| LOC113494762 | octopamine receptor Oamb isoform X1 | 12 | 65.3 | 1.1 | regulation of transcription (P) |
| 24 | 18.4 | 0.7 | |||
| LOC113495982 | uncharacterized protein | 12 | 7.7 | −1.0 | n.a. |
| 24 | 74.8 | −1.3 | |||
| LOC113501619 | organic cation transporter protein-like | 12 | 43.4 | −2.5 | transmembrane transport (P) |
| 24 | 24.1 | −2.3 | |||
| LOC113506861 | uncharacterized protein | 12 | 37.4 | −2.6 | n.a. |
| 24 | 24.0 | −2.3 | |||
| LOC113492056 | glucose dehydrogenase [FAD, quinone]-like | 12 | 34.6 | −2.7 | oxidoreductase activity (M) |
| 24 | 26.4 | −2.2 | |||
| LOC113495496 | thyrotropin-releasing hormone receptor-like isoform X1 | 12 | 3.8 | −0.2 | signaling (P) |
| 24 | 51.7 | 1–6 | |||
| LOC113500144 | uncharacterized protein | 12 | 39.2 | −0.2 | n.a. |
| 24 | 14.2 | −1.6 | |||
| LOC113506931 | proton-coupled amino acid transporter-like protein pathetic | 12 | 36.3 | −1.4 | transmembrane transport (P) |
| 24 | 6.1 | −1.2 | |||
| LOC113500825 | acid sphingomyelinase-like phosphodiesterase 3a | 12 | 4.5 | −0.9 | hydrolase activity (M) |
| 24 | 34.2 | −2.1 | |||
| LOC113498260 | mitochondrial import receptor subunit TOM40 homolog | 12 | 29.8 | 0.3 | transmembrane transport (P) |
| 24 | 7.7 | −0.6 | |||
| LOC113500192 | glutamate receptor ionotropic, kainate 2-like | 12 | 18.8 | 3.0 | regulation of transcription (P) |
| 24 | 18.4 | 2.2 |
| Gene ID | Gene name | hpi | FC | logCPM | Biological Process (P) or Molecular Function (M) |
|---|---|---|---|---|---|
| LOC113496224 | cuticle protein 16.5-like | 12 | 168.3 | −1.0 | structural molecule activity (M) |
| 24 | 84.3 | −1.4 | |||
| LOC113492802 | juvenile hormone esterase-like | 12 | 11.5 | −0.1 | hydrolase activity (M) |
| 24 | 165.3 | −0.5 | |||
| LOC113495404 | cytochrome P450 9e2-like | 12 | 163.0 | −1.1 | oxidoreductase activity (M) |
| 24 | 13.6 | −1.3 | |||
| LOC113496454 | homeobox protein abdominal-A homolog isoform X1 | 12 | 41.7 | 0.7 | programmed cell death (P) |
| 24 | 120.1 | −0.9 | |||
| LOC113508341 | uncharacterized protein | 12 | 31.2 | 1.2 | n.a. |
| 24 | 83.5 | 0.2 | |||
| LOC113498352 | apolipophorins isoform X2 | 12 | 16.2 | 1.3 | lipid transport (P) |
| 24 | 49.9 | 0.2 | |||
| LOC113493958 | solute carrier family 15 member 1-like | 12 | 36.2 | −1.3 | transmembrane transport (P) |
| 24 | 26.3 | −2.3 | |||
| LOC113496890 | Acanthoscurrin-1-like | 12 | 25.3 | −1.7 | n.a. |
| 24 | 34.5 | −2.1 | |||
| LOC113501789 | uncharacterized protein | 12 | 19.4 | 2.7 | anatomical structure development (P) |
| 24 | 31.2 | 1.3 | |||
| LOC113499613 | gloverin-like | 12 | 21.3 | 2.9 | n.a. |
| 24 | 26.4 | 1.5 | |||
| LOC113503497 | chondroitin proteoglycan 2-like | 12 | 14.2 | −0.7 | chitin binding (M) |
| 24 | 31.3 | −1.0 | |||
| LOC113496839 | protein toll-like | 12 | 36.6 | 0.3 | immune system process (P) |
| 24 | 8.8 | −1.3 | |||
| LOC113492804 | esterase FE4-like | 12 | 29.1 | 1.3 | hydrolase activity (M) |
| 24 | 15.1 | 0.5 | |||
| LOC113494776 | myb-like protein D | 12 | 7.0 | −2.3 | n.a. |
| 24 | 34.2 | −2.1 | |||
| LOC113494175 | alpha-tocopherol transfer protein-like | 12 | 12.7 | −1.0 | n.a. |
| 24 | 25.3 | −2.3 | |||
| LOC113503288 | acetylcholine receptor subunit alpha-like 1 | 12 | 23.4 | −0.3 | signaling (P) |
| 24 | 14.4 | −0.8 | |||
| LOC113507593 | probable E3 ubiquitin-protein ligase bre1 isoform X1 | 12 | 6.1 | −1.0 | n.a. |
| 24 | 30.2 | −1.0 | |||
| LOC113503500 | odorant receptor 67c-like | 12 | 4.7 | −1.8 | signaling (P) |
| 24 | 31.3 | −2.2 | |||
| LOC113505115 | irregular chiasm C-roughest protein-like isoform X1 | 12 | 16.5 | −0.3 | programmed cell death (P) |
| 24 | 15.3 | −1.7 | |||
| LOC113506962 | neural retina-specific leucine zipper protein-like | 12 | 25.1 | 2.3 | regulation of transcription (P) |
| 24 | 5.5 | 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvano, M.; Correia, R.; Virgolini, N.; Clarke, C.; Alves, P.M.; Isidro, I.A.; Roldão, A. Gene Expression Analysis of Adapted Insect Cells during Influenza VLP Production Using RNA-Sequencing. Viruses 2022, 14, 2238. https://doi.org/10.3390/v14102238
Silvano M, Correia R, Virgolini N, Clarke C, Alves PM, Isidro IA, Roldão A. Gene Expression Analysis of Adapted Insect Cells during Influenza VLP Production Using RNA-Sequencing. Viruses. 2022; 14(10):2238. https://doi.org/10.3390/v14102238
Chicago/Turabian StyleSilvano, Marco, Ricardo Correia, Nikolaus Virgolini, Colin Clarke, Paula M. Alves, Inês A. Isidro, and António Roldão. 2022. "Gene Expression Analysis of Adapted Insect Cells during Influenza VLP Production Using RNA-Sequencing" Viruses 14, no. 10: 2238. https://doi.org/10.3390/v14102238
APA StyleSilvano, M., Correia, R., Virgolini, N., Clarke, C., Alves, P. M., Isidro, I. A., & Roldão, A. (2022). Gene Expression Analysis of Adapted Insect Cells during Influenza VLP Production Using RNA-Sequencing. Viruses, 14(10), 2238. https://doi.org/10.3390/v14102238