
Prevalence and Genomic Sequence Analysis of Domestic Cat Hepadnavirus in the United States

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Isolation
2.3. PCR Protocols
2.4. Genome Sequencing of Positive Sample
3. Results
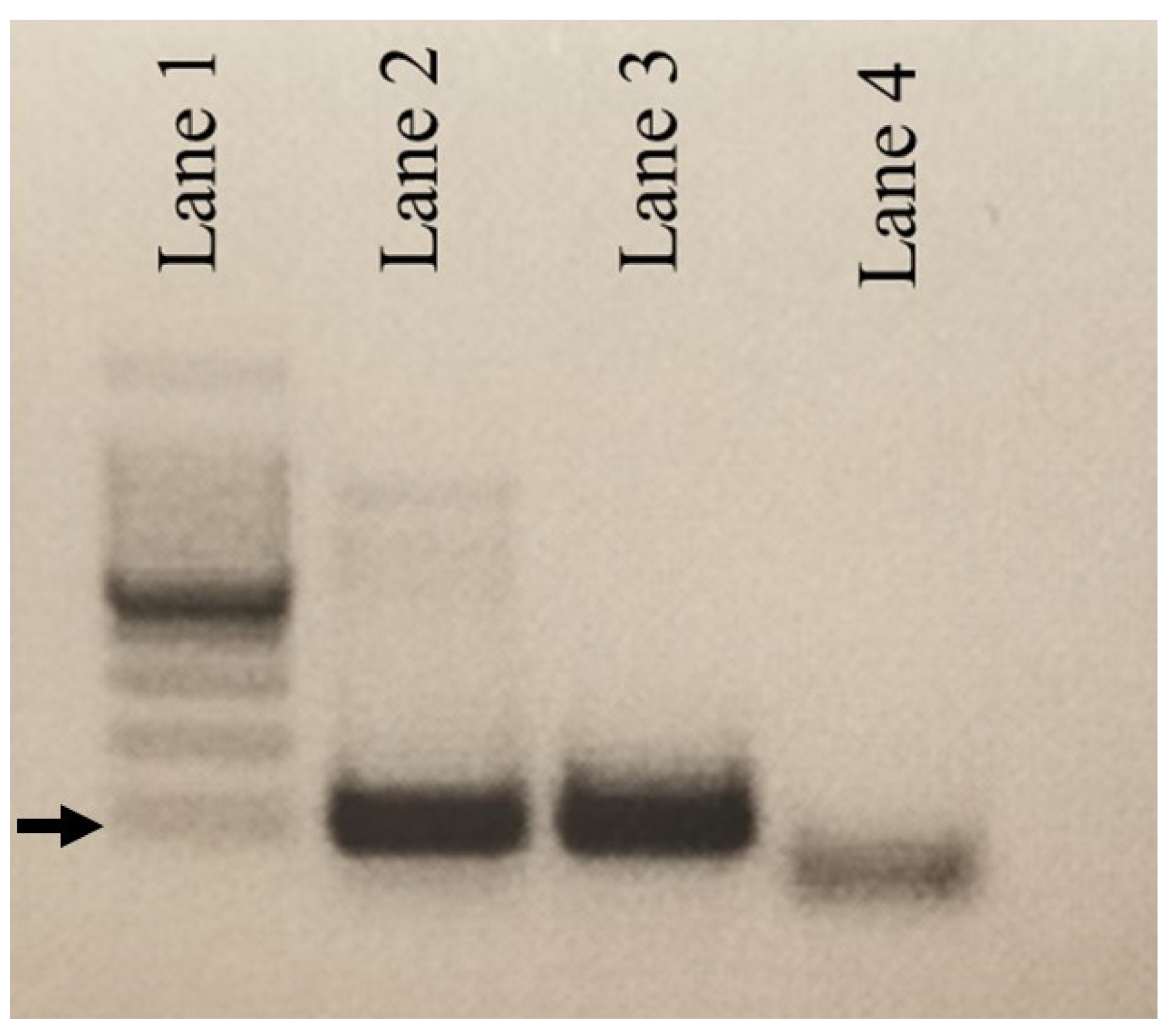
3.1. Molecular Detection of DCH
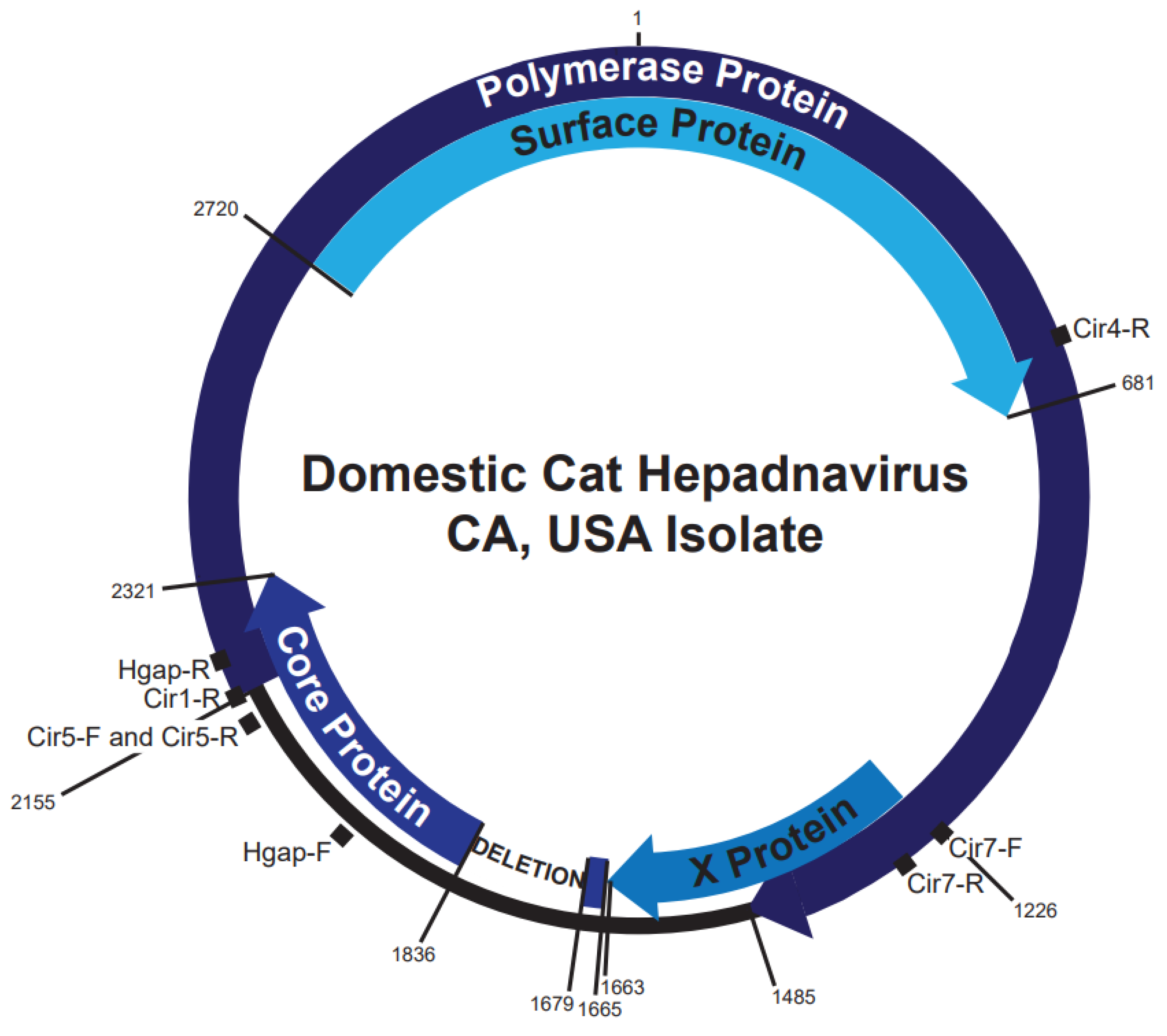
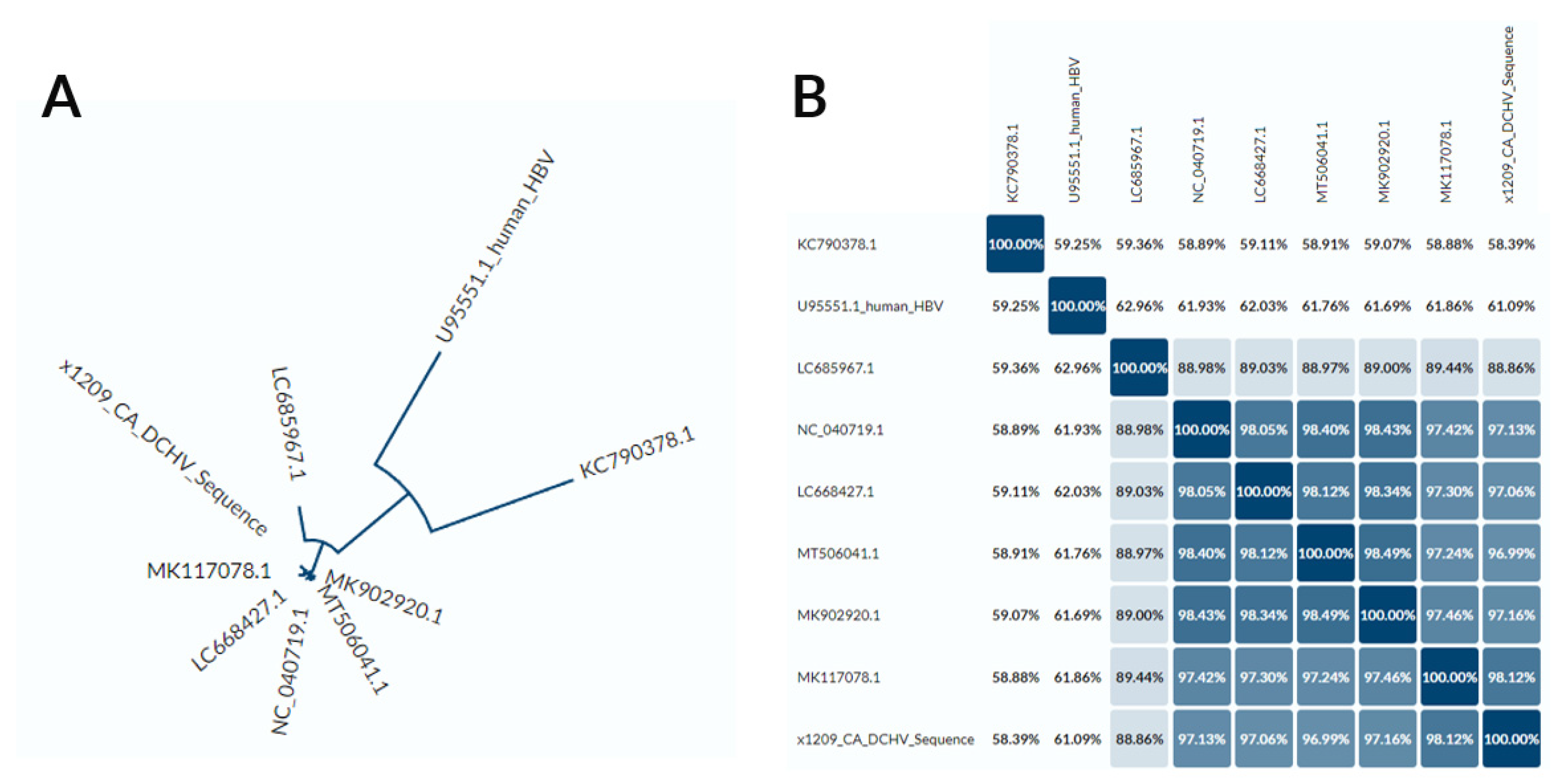
3.2. DCH Genomic Analysis
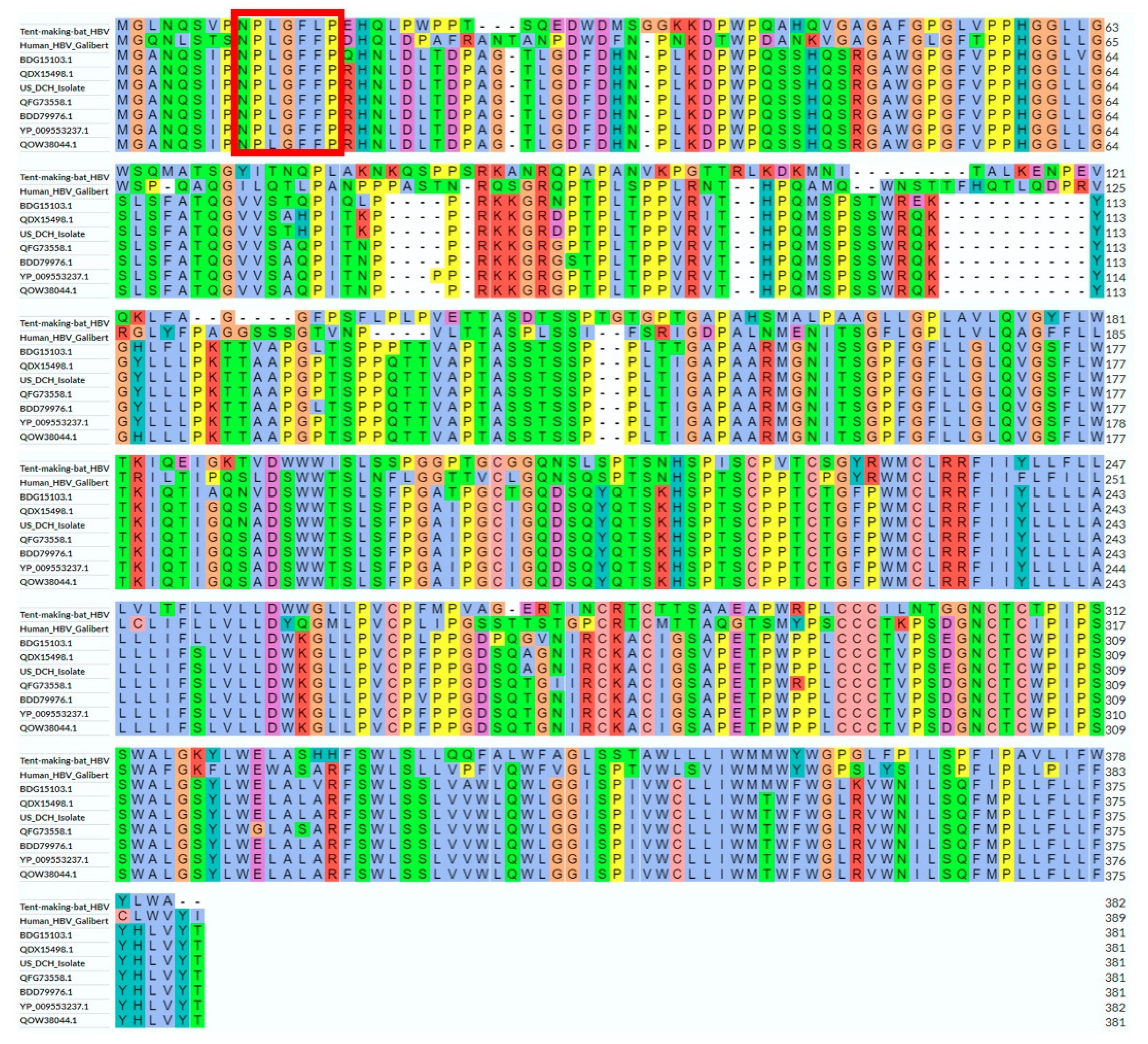
3.3. DCH Surface Protein Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aghazadeh, M.; Shi, M.; Barrs, V.R.; McLuckie, A.J.; Lindsay, S.A.; Jameson, B.; Hampson, B.; Holmes, E.C.; Beatt, J.A. A Novel Hepadnavirus Identified in an Immunocompromised Domestic Cat in Australia. Viruses 2018, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, A. Molecular Biology of the Hepatitis B Virus; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Seeger, C.; Mason, W. Hepatitis B Virus Biology. Microbiol. Mol. Biol. Rev. 2000, 64, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Ju-Tao, G.; Zhou, H.; Liu, C.; Aldrich, C.; Saputelli, J.; Whitaker, T.; Barrasa, M.I.; Mason, W.S.; Seeger, C. Apoptosis and Regeneration of Hepatocytes during Recovery from Transient Hepadnavirus Infections. J. Virol. 2000, 74, 1495–1505. [Google Scholar]
- Gwak, G.-Y.; Cho, J.; Paik, S.W.; Yoo, B.C. Comparison of clinical Manifestations and Outcomes between Hepatitis B Virus- and Hepatitis C Virus-Related Hepatocellular Carcinoma: Analysis of a Nationwide Cohort. PLoS ONE 2014, 9, e0116652. [Google Scholar]
- Robinson, M.D.; William, S. Molecular Events in the Pathogenesis of Hepadnavirus-Associated Hepatocellular Carcinoma. Annu. Rev. Med. 1994, 45, 297–323. [Google Scholar] [CrossRef]
- Guidotti, L.G. Viral Clearance Without Destruction of Infected Cells During Acute HBV Infection. Science 1999, 284, 825–829. [Google Scholar] [CrossRef]
- Tuttleman, J.; Summers, P. Formation of the Pool of Covalently Closed Circular Viral DNA in Hepadnavirus-Infected Cells. Cell 1986, 3, 451–460. [Google Scholar] [CrossRef]
- Maclachlan, J.; Dubovi, E. Fenner’s Veterinary Virology; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Kodama, K.; Yoshikawa, O.; Murakami, S. Nucleotide Sequence of a Cloned Woodchuck Hepatitis Virus Genome: Evolutional Relationship between Hepadnaviruses. J. Virol. 1995, 56, 978–986. [Google Scholar] [CrossRef]
- Wang, B.; Yang, X.-L.; Li, W.; Zhu, Y.; Ge, X.-Y.; Zhang, L.-B.; Zhang, Y.-Z.; Bock, C.-T.; Shi, Z.-L. Detection and Genome Characterization of Four Novel Bat Hepadnaviruses and a Hepevirus in China. Virol. J. 2017, 14, 40. [Google Scholar] [CrossRef]
- Freiman, J.S.; Cossart, Y.E. Natural duck hepatitis B virus infection in Australia. Aust. J. Exp. Biol. Med. Sci. 1986, 64, 477–484. [Google Scholar] [CrossRef]
- Yokosuka, O.; Omata, M.; Zhou, Y.Z.; Okuda, K. Duck hepatitis B virus DNA in liver and serum of Chinese ducks: Integration of viral DNA in a hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 1985, 18, 5180–5184. [Google Scholar] [CrossRef] [PubMed]
- Freiman, J.; Jilbert, A.R.; Dixon, R.J.; Holmes, M.; Gowans, E.J.; Burrell, C.J.; Wills, E.J.; Cossart, E.Y. Experimental duck hepatitis B virus infection: Pathology and evolution of hepatic and extrahepatic infection. Am. Assocation Study Liver Dis. 1988, 8, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Munson, L.; Worley, M.B. Veno-Occlusive Disease in Snow Leopards (Panthera uncia) from Zoological Parks. Vet. Pathol. 1991, 28, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Lanave, G.; Capozza, P.; Diakoudi, G.; Catella, C.; Catucci, L.; Ghergo, P.; Stasi, F.; Barrs, V.; Beatty, J.; Decaro, N.; et al. Identification of Hepadnavirus in the Sera of Cats. Sci. Rep. 2019, 9, 10668. [Google Scholar] [CrossRef] [PubMed]
- Pesavento, P.; Jackson, K.; Scase, T.; Tse, T.; Hampson, B.; Munda, J.S.; Barrs, V.R.; Beatty, J.A. A Novel Hepadnavirus Is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats. Viruses 2019, 11, 969. [Google Scholar] [CrossRef]
- Goussev, S.; Center, S.A.; Randolph, J.F.; Kathrani, A.; Butler, B.P.; McDonough, S.P. Clinical Characteristics of Hepatocellular Carcinoma in 19 Cats from a Single institution. J. Am. Anim. Hosp. Assoc. 2016, 52, 36–41. [Google Scholar] [CrossRef]
- Carver, S.; Bevins, S.N.; Lappin, M.R.; Boydston, E.E.; Lyren, L.M.; Alldredge, M.; Logan, K.A.; Sweanor, L.L.; Riley, S.P.D.; Serieys, L.E.K.; et al. Pathogen exposure varies widely among sympatric populations of wild and domestic felids across the United States. Ecol. Appl. 2015, 26, 367–381. [Google Scholar] [CrossRef]
- Powers, J.; Chiu, E.S.; Kraberger, S.J.; Roelke-Parker, M.; Lowery, I.; Erbeck, K.; Troyer, R.; Carver, S.; Woude, S.V. Feline Leukemia Virus (FeLV) Disease Outcomes in a Domestic Cat Breeding Colony: Relationship to Endogenous FeLV and Other Chronic Viral Infections. J. Virol. 2018, 92, e00649-18. [Google Scholar] [CrossRef]
- Ledesma-Felicano, C.; Hagen, S.; Troyer, R.; Zheng, X.; Musselman, E.; Lukic, D.S.; Franke, A.-M.; Maeda, D.; Zielonka, J. Replacement of feline foamy virus bet by feline immunodeficiency virus vif yields replicative virus with novel vaccine candidate potential. Retrovirology 2018, 15, 38. [Google Scholar] [CrossRef]
- Ledesma-Feliciano, C.; Troyer, R.M.; Zheng, X.; Miller, C.; Cianciolo, R.; Bordicchia, M.; Dannemiller, N.; Gagne, R.; Beatty, J.; Quimby, J.; et al. Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease. Viruses 2019, 11, 662. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2020, 49, D480–D489. [Google Scholar]
- Bologna, G.; Yvon, C.; Duvaud, S.; Veuthey, A.-L. N-terminal Mysristoylation Predictions by Ensembles of Neural Networks. Proteomics 2004, 6, 1626–1632. [Google Scholar] [CrossRef] [PubMed]
- Fallows, D.A.; Goff, S.P. Mutations in the epsilon sequences of human hepatitis B virus affect both RNA encapsidation and reverse transcription. J. Virol. 1995, 69, 3067–3073. [Google Scholar] [CrossRef] [PubMed]
- Tu, T.; Budzinska, M.A.; Shackel, N.A.; Urban, S. HBV DNA Integration: Molecular Mechanisms and Clinical Implications. Viruses 2017, 9, 75. [Google Scholar] [CrossRef]
- Yang, W.; Mason, W.S.; Summers, J. Covalently closed circular viral DNA formed from two types of linear DNA in woodchuck hepatitis virus-infected liver. J. Virol. 1996, 70, 4567–4575. [Google Scholar] [CrossRef]
- Yang, W.; Summers, J. Illegitimate replication of linear hepadnavirus DNA through nonhomologous recombination. J. Virol. 1995, 69, 4029–4036. [Google Scholar] [CrossRef]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef]
- Yi, N.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Fälth, M.; Stindt, J.; Königer, C.; Nassal, M.; Kubitz, R.; et al. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology 2014, 146, 1070–1083.e6. [Google Scholar]
- Drexler, J.; Geipel, A.; König, A.; Drosten, C. Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 24043818. [Google Scholar] [CrossRef]
- Schulze, A.; Schieck, A.; Ni, Y.; Mier, W.; Urban, S. Fine Mapping of Pre-S Sequence Requirements for Hepatitis B Virus Large Envelope Protein-Mediated Receptor Interaction. J. Virol. 2010, 84, 1989–2000. [Google Scholar] [CrossRef]
- Shofa, M.; Kaneko, Y.; Takahashi, K.; Okabayashi, T.; Saito, A. Global Prevalence of Domestic Cat Hepadnavirus: An Emerging Threat to Cats’ Health? Front. Microbiol. 2022, 13, 938154. [Google Scholar] [CrossRef] [PubMed]
- Asabe, S.; Wieland, S.F.; Chattopadhyay, P.K.; Roederer, M.; Engle, R.E.; Purcell, R.H.; Chisar, F.V. The size of the viral inoculum contributes to the outcome of hepatitis B virus infection. J. Virol. 2009, 83, 9652–9662. [Google Scholar] [CrossRef] [PubMed]
- Beczkowski, P.; Litsterc, A.; Lin, T.L.; Mellore, D.J.; Willetta, B.J.; Hosie, M.J. Contrasting clinical outcomes in two cohorts of cats naturally infected with feline immunodeficiency virus (FIV) Author links open overlay panel. Vet. Microbiol. 2015, 176, 50–60. [Google Scholar] [CrossRef]
- Yang, W.; Summers, J. Infection of Ducklings with Virus Particles Containing Linear Double-Stranded Duck Hepatitis B Virus DNA: Illegitimate Replication and Reversion. J. Virol. 1998, 72, 8710–8717. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-L.; Ho, M.-C.; Lin, Y.-Y.; Tzeng, S.-T.; Chen, Y.-J.; Pai, H.-Y.; Wang, Y.-C.; Chen, C.-L.; Lee, Y.-H.; Chen, D.-S.; et al. Cell-Free Virus-Host Chimera DNA From Hepatitis B Virus Integration Sites as a Circulating Biomarker of Hepatocellular Cancer. Am. Assoc. Study Liver Dis. Hepatol. 2020, 72, 2063–2076. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cui, X.; Huo, Q.; Qi, Y.; Sun, Y.; Meihua, T.; Qingsheng, K. Profile of HBV Integration in the Plasma DNA of Hepatocellular Carcinoma Patients. Curr. Genom. 2019, 20, 61–68. [Google Scholar] [CrossRef]
- Lin, S.; Su, Y.-P.; Trauger, E.R.; Song, B.P.; Thompson, E.G.C.; Hoffman, M.C.; Chang, T.-T.; Lin, Y.-J.; Kao, Y.-L.; Cui, Y.; et al. Detection of Hepatitis B Virus–Host Junction Sequences in Urine of Infected Patients. Am. Assoc. Study Liver Dis. Hepatol. Commun. 2021, 5, 1649–1659. [Google Scholar] [CrossRef]
- Georget, R.; Le Mire, M.F.; Mason, W.S.; Jilbert, A.R. The persistence in the liver of residual duck hepatitis B virus covalently closed circular DNA is not dependent upon new viral DNA synthesis. Virology 2010, 406, 286–292. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Cohort | Reference | Tissue Sampled | Positive Conventional PCR Analyses/Total Analyses | Positive qPCR Confirmatory Analyses/Total Analyses | DCH Positive Samples/Total Samples | Positive (%) |
|---|---|---|---|---|---|---|
| Multi-cat household | Powers, Jordan, et al., Journal of Virology [20] | WB DNA | 0/57 | 0/57 | 0/57 | 0 |
| Cats presented to shelters in Colorado, California, Florida | Carver, Scott, et al., Ecological Applications [19] | WB DNA | 1/286 | 1/286 | 1/286 | 0.35 |
| CSU Hematopathology Laboratory | Gift of Anne Avery, Laboratory Director | Hepatic DNA | 0/84 | 0/84 | 0/84 | 0 |
| CSU VDL necropsy | CSU Diagnostic Laboratory | Hepatic DNA | 0/11 | 0/11 | 0/11 | 0 |
| Memphis animal shelters and households | Ledesma-Felicano, Carmen, et al., Retrovirology [21] | Plasma | 0/58 | 0/58 | 0/58 | 0 |
| TOTAL US | 496 | 496 | 1/496 | 0.2 | ||
| University of Sydney, Sydney Australia | Ledesma-Feliciano, Carmen, et al., Viruses [22] | WB DNA/Plasma | 4/67 | 7/67 | 7/67 | 10.4 |
| qPCR Primers | Sequence 5′–3′ | Binding Site |
|---|---|---|
| FHBV F | CGTCATCATGGGTTTAGGAA | Nt 458→478, surface and polymerase proteins |
| HBV-R | TCCATATAAGCAAACACCATACAAT | Nt 589→565, surface and polymerase proteins |
| FHBV P | [FAM]TCCTCCTAACCATTGAAGCCAGACTACT [BHQ] | Nt 528→555, surface and polymerase proteins |
| Conventional PCR Primers | Sequence 5′–3′ | Binding Site |
| Cir 5f | TTGGCACCTGGATTCGCA | Nt 2113→2130, core protein |
| Cir 4R | AGATGTTCCACACTCTTAGCC | Nt 625→605, surface and polymerase proteins |
| Cir 7R | CGTAGACGAAGGACACGTC | Nt 1281→1262, polymerase and X proteins |
| Cir 7F | CCATCGATTTACACACTTCCCA | Nt 1205→1226, polymerase protein |
| Cir 5R | TGCGAATCCAGGTGCCAA | Nt 2130→2113, core protein |
| Cir 1R | ATAACCGTATGCTCCGGAAG | Nt 2194→2175, core and polymerase proteins |
| Hgap F | GTGCTCTGATAACCGTATGCTC | Nt 1945→1968, core protein |
| Hgap R | CTAGAATGGCTACATGGGAG | Nt 2203→2181, core and polymerase proteins |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stone, C.; Petch, R.; Gagne, R.B.; Nehring, M.; Tu, T.; Beatty, J.A.; VandeWoude, S. Prevalence and Genomic Sequence Analysis of Domestic Cat Hepadnavirus in the United States. Viruses 2022, 14, 2091. https://doi.org/10.3390/v14102091
Stone C, Petch R, Gagne RB, Nehring M, Tu T, Beatty JA, VandeWoude S. Prevalence and Genomic Sequence Analysis of Domestic Cat Hepadnavirus in the United States. Viruses. 2022; 14(10):2091. https://doi.org/10.3390/v14102091
Chicago/Turabian StyleStone, Cassidy, Raegan Petch, Roderick B. Gagne, Mary Nehring, Thomas Tu, Julia A. Beatty, and Sue VandeWoude. 2022. "Prevalence and Genomic Sequence Analysis of Domestic Cat Hepadnavirus in the United States" Viruses 14, no. 10: 2091. https://doi.org/10.3390/v14102091
APA StyleStone, C., Petch, R., Gagne, R. B., Nehring, M., Tu, T., Beatty, J. A., & VandeWoude, S. (2022). Prevalence and Genomic Sequence Analysis of Domestic Cat Hepadnavirus in the United States. Viruses, 14(10), 2091. https://doi.org/10.3390/v14102091