
Genomics, Transcriptomics, and Proteomics of SSV1 and Related Fusellovirus: A Minireview

 ,
,  ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
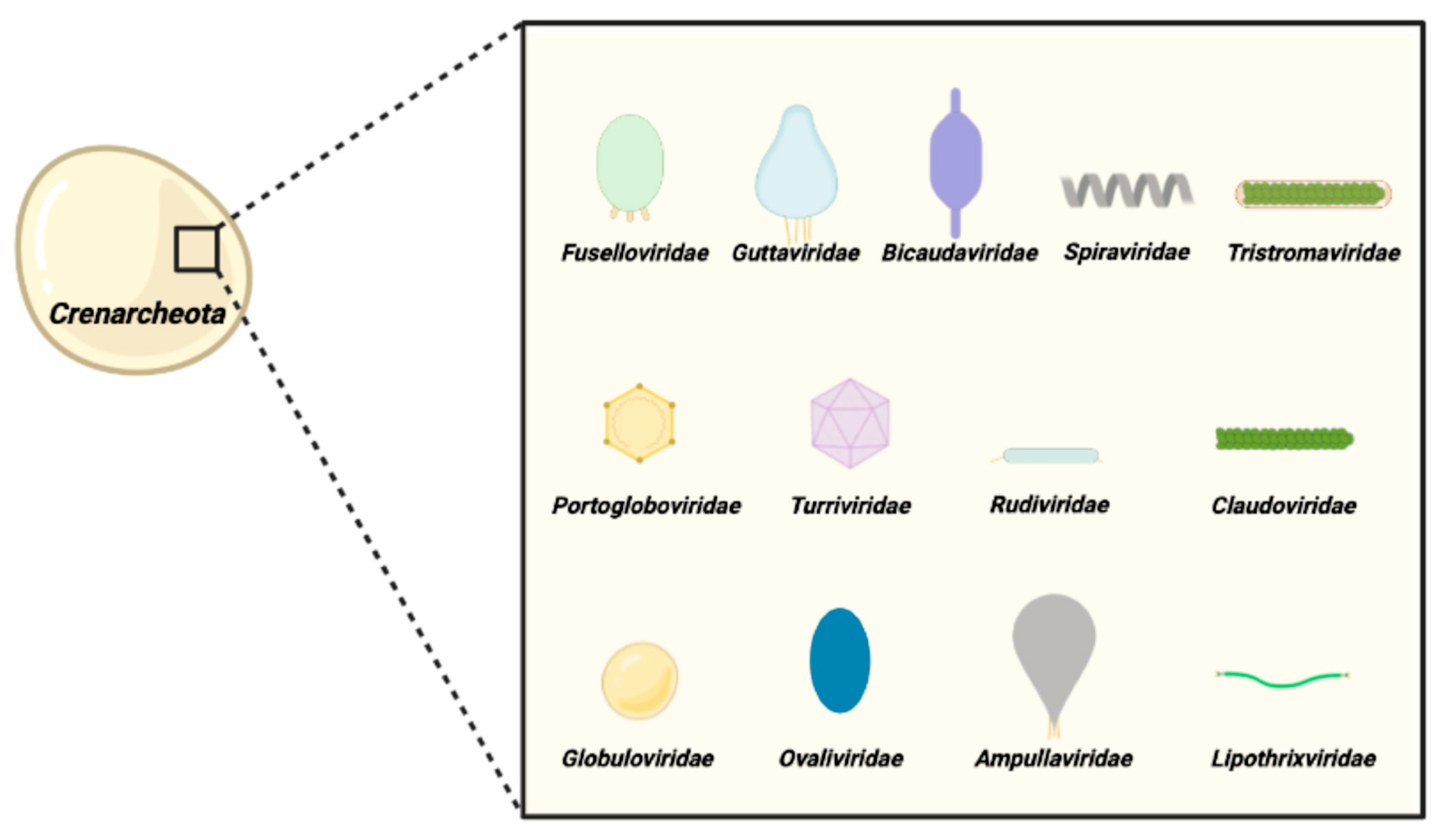
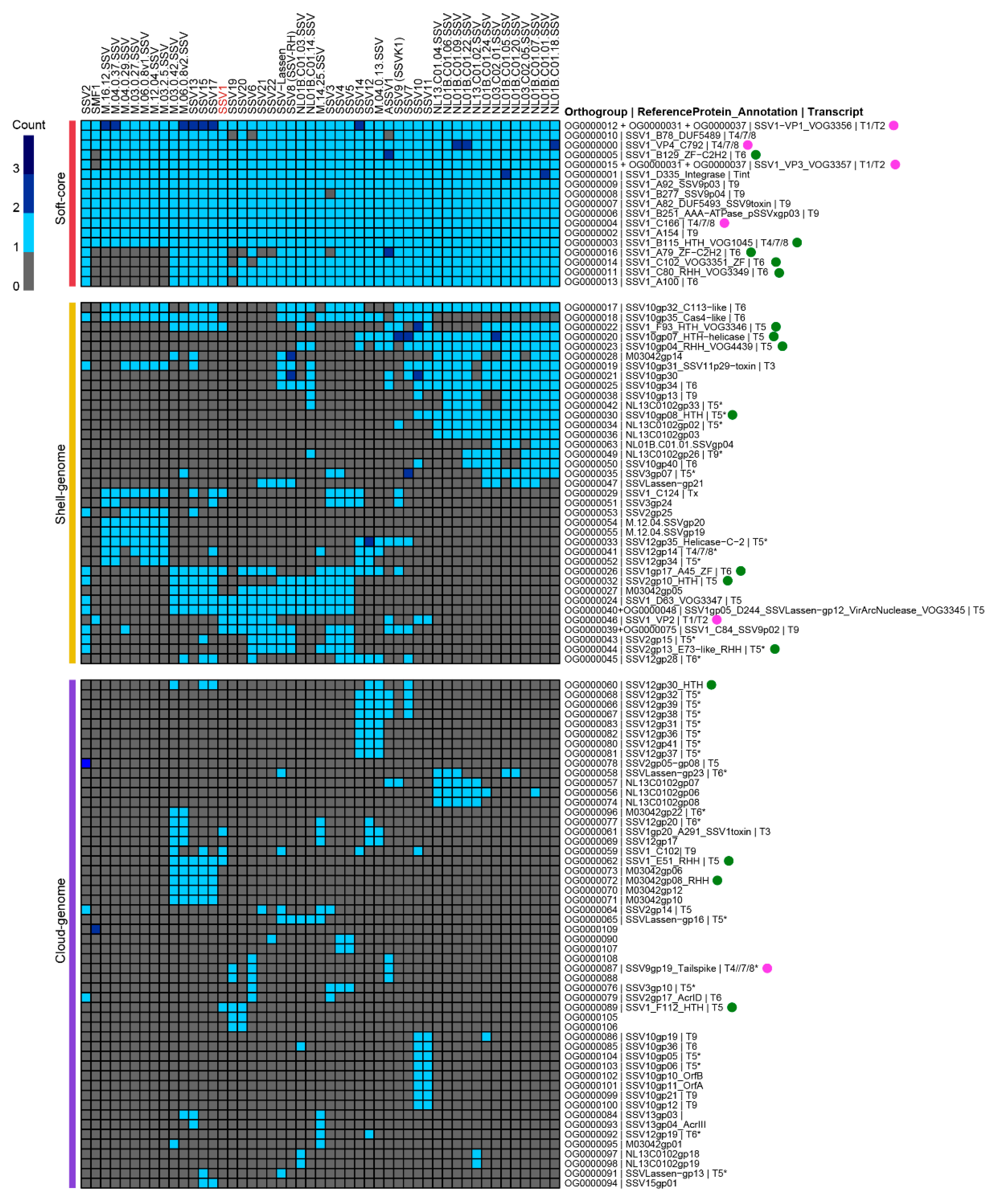
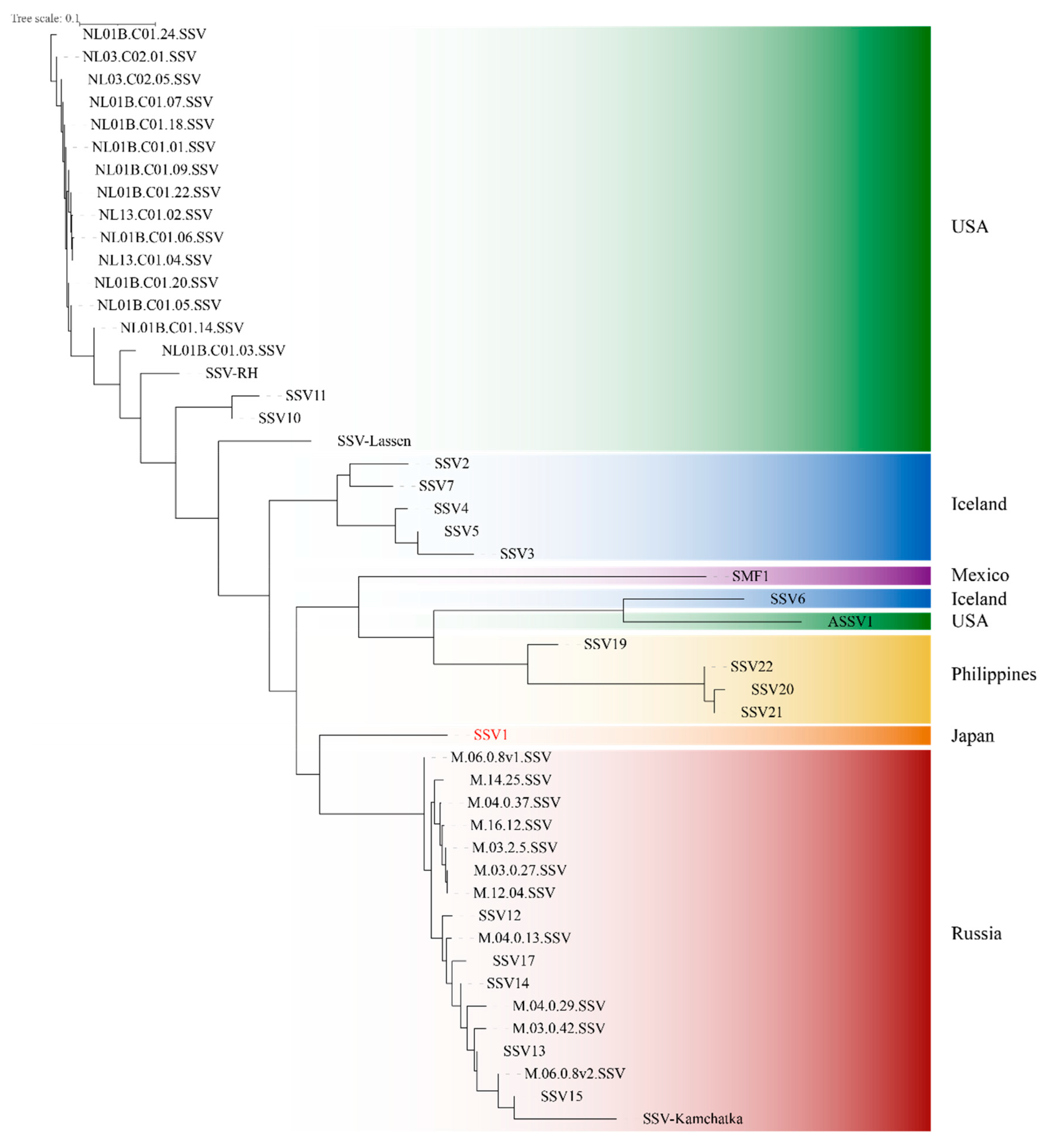
2. Genomic Analysis
2.1. The Fusellovirus Core Genome
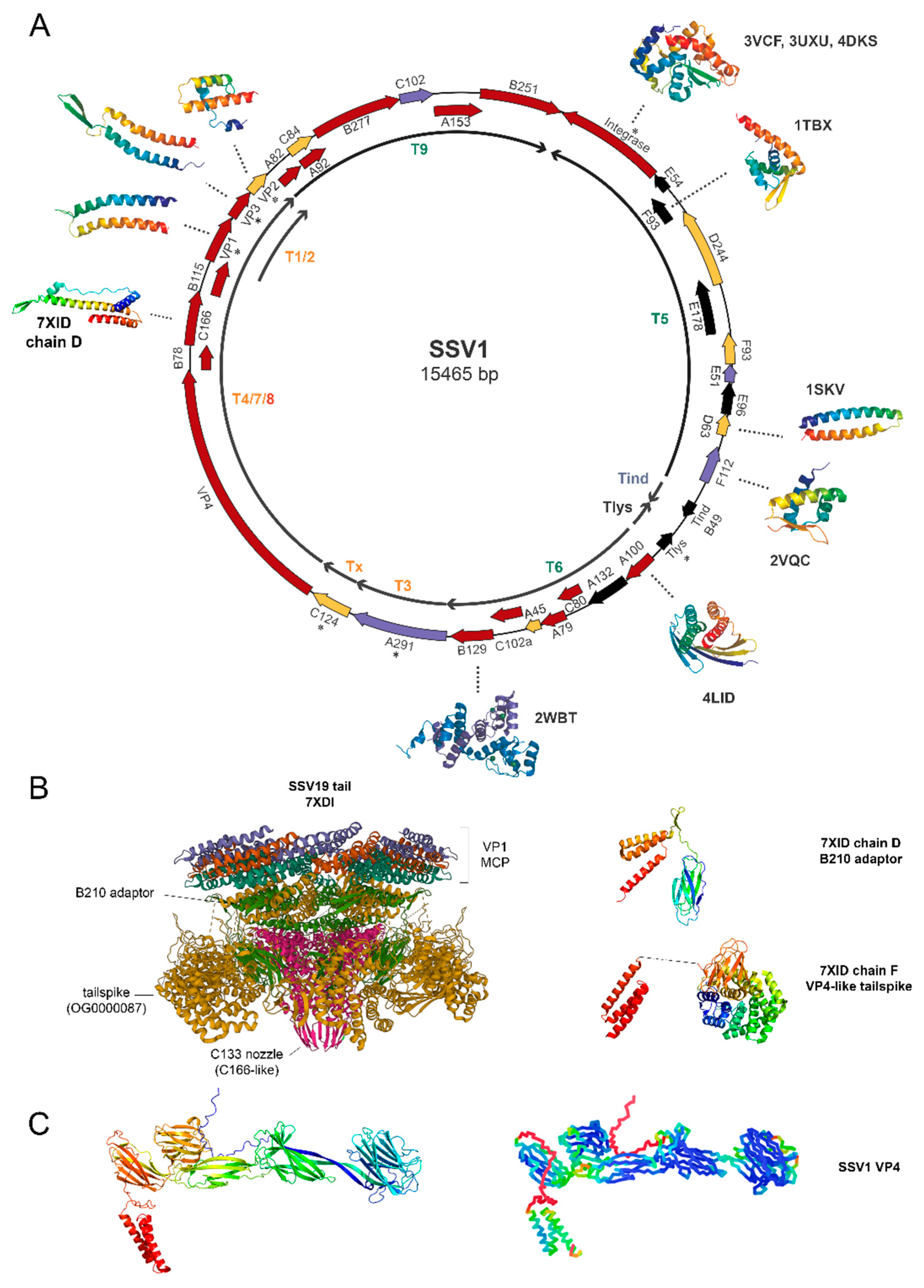
2.2. Location of Core Genes
2.3. Functional Annotation of Core Genes
2.4. The Fusellovirirus Pangenome Contains Multiple Transcriptional Regulators
3. Transcriptomic Analysis
3.1. SSV1 Genes Regulation
3.2. SSV1 Genes Expressed in the Carrier State
3.3. SSV1 Genes Expressed upon UV Induction
3.4. Promoters and Terminators of SSV1
3.5. SSV1 Transcription Regulation
4. Proteomic Analysis
5. Host–Virus Interaction
5.1. Transcriptome Analysis of S. Solfataricus
5.2. Regulation of CRISPR-Cas System during the Infection
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Prangishvili, D.; Bamford, D.H.; Forterre, P.; Iranzo, J.; Koonin, E.V.; Krupovic, M. The enigmatic archaeal virosphere. Nat. Rev. Microbiol. 2017, 15, 724–739. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.F.; Mesa, V.; Estrada-Ortiz, N.; Lopez-Obando, M.; Gómez, A.; Plácido, J. Viruses in extreme environments, current overview, and biotechnological potential. Viruses 2021, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Maaty, W.S.A.; Ortmann, A.C.; Dlakić, M.; Schulstad, K.; Hilmer, J.K.; Liepold, L.; Weidenheft, B.; Khayat, R.; Douglas, T.; Young, M.J.; et al. Characterization of the Archaeal Thermophile Sulfolobus Turreted Icosahedral Virus Validates an Evolutionary Link among Double-Stranded DNA Viruses from All Domains of Life. J. Virol. 2006, 80, 7625–7635. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M.; Cvirkaite-Krupovic, V.; Iranzo, J.; Prangishvili, D.; Koonin, E.V. Viruses of archaea: Structural, functional, environmental and evolutionary genomics. Virus Res. 2018, 244, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Baquero, D.P.; Contursi, P.; Piochi, M.; Bartolucci, S.; Liu, Y.; Cvirkaite-Krupovic, V.; Prangishvili, D.; Krupovic, M. New virus isolates from Italian hydrothermal environments underscore the biogeographic pattern in archaeal virus communities. ISME J. 2020, 14, 1821–1833. [Google Scholar] [CrossRef]
- Contursi, P.; Fusco, S.; Cannio, R.; She, Q. Molecular biology of fuselloviruses and their satellites. Extremophiles 2014, 18, 473–489. [Google Scholar] [CrossRef]
- Sakai, H.D.; Kurosawa, N. Saccharolobus caldissimus gen. nov., sp. nov., a facultatively anaerobic iron-reducing hyperthermophilic archaeon isolated from an acidic terrestrial hot spring, and reclassification of Sulfolobus solfataricus as saccharolobus solfataricus comb. nov. and. Int. J. Syst. Evol. Microbiol. 2018, 68, 1271–1278. [Google Scholar] [CrossRef]
- Wiedenheft, B.; Stedman, K.; Roberto, F.; Willits, D.; Gleske, A.-K.; Zoeller, L.; Snyder, J.; Douglas, T.; Young, M. Comparative Genomic Analysis of Hyperthermophilic Archaeal Fuselloviridae Viruses. J. Virol. 2004, 78, 1954–1961. [Google Scholar] [CrossRef]
- Contursi, P.; Jensen, S.; Aucelli, T.; Rossi, M.; Bartolucci, S.; She, Q. Characterization of the Sulfolobus host-SSV2 virus interaction. Extremophiles 2006, 10, 615–627. [Google Scholar] [CrossRef]
- Aucelli, T.; Contursi, P.; Girfoglio, M.; Rossi, M.; Cannio, R. A spreadable, non-integrative and high copy number shuttle vector for Sulfolobus solfataricus based on the genetic element pSSVx from Sulfolobus islandicus. Nucleic Acids Res. 2006, 34, e114. [Google Scholar] [CrossRef]
- Goodman, D.A.; Stedman, K.M. Comparative genetic and genomic analysis of the novel fusellovirus Sulfolobus spindle-shaped virus 10. Virus Evol. 2018, 4, vey022. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, R.M.; Marceau, C.D.; Marceau, J.O.; Morris, S.; Clore, A.J.; Stedman, K.M. Differential virus host-ranges of the Fuselloviridae of hyperthermophilic Archaea: Implications for evolution in extreme environments. Front. Microbiol. 2012, 3, 295. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, R.M.; Drummond, C.G.; Stacy, C.L.; Padilla-Crespo, E.; Stedman, K.M. Host-Dependent Differences in Replication Strategy of the Sulfolobus Spindle-Shaped Virus Strain SSV9 (a.k.a., SSVK1): Infection Profiles in Hosts of the Family Sulfolobaceae. Front. Microbiol. 2020, 11, 1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, X.; Wang, H.; Jiang, H.; Dong, H.; Huang, L. Novel Sulfolobus Fuselloviruses with Extensive Genomic Variations. J. Virol. 2020, 94, 1624–1643. [Google Scholar] [CrossRef]
- Fusco, S.; Aulitto, M.; Bartolucci, S.; Contursi, P. A standardized protocol for the UV induction of Sulfolobus spindle-shaped virus 1. Extremophiles 2015, 19, 539–546. [Google Scholar] [CrossRef][Green Version]
- Iverson, E.A.; Goodman, D.A.; Gorchels, M.E.; Stedman, K.M. Extreme Mutation Tolerance: Nearly Half of the Archaeal Fusellovirus Sulfolobus Spindle-Shaped Virus 1 Genes Are Not Required for Virus Function, Including the Minor Capsid Protein Gene vp3. J. Virol. 2017, 91, e02406–e2416. [Google Scholar] [CrossRef]
- Reiter, W.D.; Palm, P.; Zillig, W. Analysis of transcription in the archaebacterium Sulfolobus indicates that archaebacterial promoters are homologous to eukaryotic pol II promoters. Nucleic Acids Res. 1988, 16, 1–19. [Google Scholar] [CrossRef]
- Jonuscheit, M.; Martusewitsch, E.; Stedman, K.M.; Schleper, C. A reporter gene system for the hyperthermophilic archaeon Sulfolobus solfataricus based on a selectable and integrative shuttle vector. Mol. Microbiol. 2003, 48, 1241–1252. [Google Scholar] [CrossRef]
- Palm, P.; Schleper, C.; Grampp, B.; Yeats, S.; McWilliam, P.; Reiter, W.D.; Zillig, W. Complete nucleotide sequence of the virus SSV1 of the archaebacterium Sulfolobus shibatae. Virology 1991, 185, 242–250. [Google Scholar] [CrossRef]
- Schleper, C.; Kubo, K.; Zillig, W. The particle SSV1 from the extremely thermophilic archaeon Sulfolobus is a virus: Demonstration of infectivity and of transfection with viral DNA. Proc. Natl. Acad. Sci. USA 1992, 89, 7645–7649. [Google Scholar] [CrossRef]
- Iverson, E.; Stedman, K. A genetic study of SSV1, the prototypical fusellovirus. Front. Microbiol. 2012, 3, 200. [Google Scholar] [CrossRef] [PubMed]
- Muskhelishvili, G.; Palm, P.; Zillig, W. SSV1-encoded site-specific recombination system in Sulfolobus shibatae. MGG Mol. Gen. Genet. 1993, 237, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Fusco, S.; Liguori, R.; Limauro, D.; Bartolucci, S.; She, Q.; Contursi, P. Transcriptome analysis of Sulfolobus solfataricus infected with two related fuselloviruses reveals novel insights into the regulation of CRISPR-Cas system. Biochimie 2015, 118, 322–332. [Google Scholar] [CrossRef]
- Reiter, W.D.; Palm, P.; Yeats, S.; Zillig, W. Gene expression in archaebacteria: Physical mapping of constitutive and UV-inducible transcripts from the Sulfolobus virus-like particle SSV1. MGG Mol. Gen. Genet. 1987, 209, 270–275. [Google Scholar] [CrossRef]
- Fröls, S.; Gordon, P.M.K.; Panlilio, M.A.; Schleper, C.; Sensen, C.W. Elucidating the transcription cycle of the UV-inducible hyperthermophilic archaeal virus SSV1 by DNA microarrays. Virology 2007, 365, 48–59. [Google Scholar] [CrossRef]
- Reiter, W.D.; Palm, P.; Zillig, W. Transcription termination in the archaebacterium Sulfolobus: Signal structures and linkage to transcription initiation. Nucleic Acids Res. 1988, 16, 2445–2460. [Google Scholar] [CrossRef] [PubMed]
- Best, A.A.; Olsen, G.J. Similar subunit architecture of archaeal and eukaryal RNA polymerases. FEMS Microbiol. Lett. 2001, 195, 85–90. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Reiter, W.-D.; Palm, P.; Henschen, A.; Lottspeich, F.; Zillig, W.; Grampp, B. Identification and characterization of the genes encoding three structural proteins of the Sulfolobus virus-like particle SSV1. Mol. Gen. Genet. MGG 1987, 206, 144–153. [Google Scholar] [CrossRef]
- Held, N.L.; Whitaker, R.J. Viral biogeography revealed by signatures in Sulfolobus islandicus genomes. Environ. Microbiol. 2009, 11, 457–466. [Google Scholar] [CrossRef]
- Redder, P.; Peng, X.; Brügger, K.; Shah, S.A.; Roesch, F.; Greve, B.; She, Q.; Schleper, C.; Forterre, P.; Garrett, R.A.; et al. Four newly isolated fuselloviruses from extreme geothermal environments reveal unusual morphologies and a possible interviral recombination mechanism. Environ. Microbiol. 2009, 11, 2849–2862. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Aulitto, M.; Martinez-Alvarez, L.; Fiorentino, G.; Limauro, D.; Peng, X.; Contursi, P. A Comparative Analysis of Weizmannia coagulans Genomes Unravels the Genetic Potential for Biotechnological Applications. Int. J. Mol. Sci. 2022, 23, 3135. [Google Scholar] [CrossRef]
- Fusco, S.; She, Q.; Bartolucci, S.; Contursi, P. Tlys, a Newly Identified Sulfolobus Spindle-Shaped Virus 1 Transcript Expressed in the Lysogenic State, Encodes a DNA-Binding Protein Interacting at the Promoters of the Early Genes. J. Virol. 2013, 87, 5926–5936. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Iverson, E.A.; Goodman, D.A.; Gorchels, M.E.; Stedman, K.M. Genetic analysis of the major capsid protein of the archaeal fusellovirus SSV1: Mutational flexibility and conformational change. Genes 2017, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, L. Construction of a Fusellovirus with a Minimal Set of Genes. ACS Synth. Biol. 2021, 10, 2617–2627. [Google Scholar] [CrossRef] [PubMed]
- Quemin, E.R.J.; Pietilä, M.K.; Oksanen, H.M.; Forterre, P.; Rijpstra, W.I.C.; Schouten, S.; Bamford, D.H.; Prangishvili, D.; Krupovic, M. Sulfolobus Spindle-Shaped Virus 1 Contains Glycosylated Capsid Proteins, a Cellular Chromatin Protein, and Host-Derived Lipids. J. Virol. 2015, 89, 11681–11691. [Google Scholar] [CrossRef]
- Wang, F.; Cvirkaite-Krupovic, V.; Vos, M.; Beltran, L.C.; Kreutzberger, M.A.B.; Winter, J.M.; Su, Z.; Liu, J.; Schouten, S.; Krupovic, M.; et al. Spindle-shaped archaeal viruses evolved from rod-shaped ancestors to package a larger genome. Cell 2022, 185, 1297–1307.e11. [Google Scholar] [CrossRef]
- Han, Z.; Yuan, W.; Xiao, H.; Wang, L.; Zhang, J.; Peng, Y.; Cheng, L.; Liu, H.; Huang, L. Structural insights into a spindle-shaped archaeal virus with a sevenfold symmetrical tail. Proc. Natl. Acad. Sci. USA 2022, 119, e2119439119. [Google Scholar] [CrossRef]
- Koonin, E.V. Archaebacterial virus SSV1 encodes a putative DNAA-like protein. Nucleic Acids Res. 1992, 20, 1143. [Google Scholar] [CrossRef]
- Arnold, H.P.; She, Q.; Phan, H.; Stedman, K.; Prangishvili, D.; Holz, I.; Kristjansson, J.K.; Garrett, R.; Zillig, W. The genetic element pSSVx of the extremely thermophilic crenarchaeon Sulfolobus is a hybrid between a plasmid and a virus. Mol. Microbiol. 1999, 34, 217–226. [Google Scholar] [CrossRef]
- Dewerff, S.J.; Zhang, C.; Schneider, J.; Whitaker, R.J. Intraspecific antagonism through viral toxin encoded by chronic Sulfolobus spindle-shaped virus. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 1842. [Google Scholar] [CrossRef]
- Kraft, P.; Oeckinghaus, A.; Kümmel, D.; Gauss, G.H.; Gilmore, J.; Wiedenheft, B.; Young, M.; Lawrence, C.M. Crystal Structure of F-93 from Sulfolobus Spindle-Shaped Virus 1, a Winged-Helix DNA Binding Protein. J. Virol. 2004, 78, 11544–11550. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.K.; Maaty, W.S.; Corn, G.J.; Kwok, S.C.; Eilers, B.J.; Kraft, P.; Gillitzer, E.; Young, M.J.; Bothner, B.; Lawrence, C.M. Cysteine usage in Sulfolobus spindle-shaped virus 1 and extension to hyperthermophilic viruses in general. Virology 2008, 376, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.M.; Menon, S.; Eilers, B.J.; Bothner, B.; Khayat, R.; Douglas, T.; Young, M.J. Structural and functional studies of archaeal viruses. J. Biol. Chem. 2009, 284, 12599–12603. [Google Scholar] [CrossRef] [PubMed]
- Fusco, S.; Aulitto, M.; Iacobucci, I.; Crocamo, G.; Pucci, P.; Bartolucci, S.; Monti, M.; Contursi, P. The interaction between the F55 virus-encoded transcription regulator and the RadA host recombinase reveals a common strategy in Archaea and Bacteria to sense the UV-induced damage to the host DNA. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194493. [Google Scholar] [CrossRef]
- Yang, Y.; Junfeng, L.; Xiaofei, F.; Zhou, F.; Zhang, S.; Zhang, X.; Huang, Q.; Krupovic, M.; She, Q.; Ni, J.; et al. A novel RHH family transcription factor aCcr1 and its viral homologs dictate cell cycle progression in archaea. bioRxiv 2022. [Google Scholar] [CrossRef]
- Li, X.; Lozano-Madueño, C.; Martínez-Alvarez, L.; Peng, X. A clade of RHH proteins ubiquitous in Sulfolobales and their viruses regulates cell cycle progression. bioRxiv 2022. [Google Scholar] [CrossRef]
- Michelin, M.; Gomes, D.G.; Romaní, A.; de Maria, L.T.M.P.; Teixeira, J.A. Nanocellulose production: Exploring the enzymatic route and residues of pulp and paper industry. Molecules 2020, 25, 3411. [Google Scholar] [CrossRef]
- Bhoobalan-Chitty, Y.; Johansen, T.B.; Di Cianni, N.; Peng, X. Inhibition of Type III CRISPR-Cas Immunity by an Archaeal Virus-Encoded Anti-CRISPR Protein. Cell 2019, 179, 448–458.e11. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. Affiliations STAG: Species Tree Inference from All Genes. bioRxiv 2018. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. STRIDE: Species tree root inference from gene duplication events. Mol. Biol. Evol. 2017, 34, 3267–3278. [Google Scholar] [CrossRef] [PubMed]
- Stedman, K.M.; DeYoung, M.; Saha, M.; Sherman, M.B.; Morais, M.C. Structural insights into the architecture of the hyperthermophilic fusellovirus SSV1. Virology 2015, 474, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Fusco, S.; She, Q.; Fiorentino, G.; Bartolucci, S.; Contursi, P. Unravelling the Role of the F55 Regulator in the Transition from Lysogeny to UV Induction of Sulfolobus Spindle-Shaped Virus 1. J. Virol. 2015, 89, 6453–6461. [Google Scholar] [CrossRef] [PubMed]
- Contursi, P.; Fusco, S.; Limauro, D.; Fiorentino, G. Host and viral transcriptional regulators in Sulfolobus: An overview. Extremophiles 2013, 17, 881–895. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Towards functional characterization of archaeal genomic dark matter. Biochem. Soc. Trans. 2019, 47, 389–398. [Google Scholar] [CrossRef]
- Jarrell, K.F.; Ding, Y.; Meyer, B.H.; Albers, S.-V.; Kaminski, L.; Eichler, J. N-Linked Glycosylation in Archaea: A Structural, Functional, and Genetic Analysis. Microbiol. Mol. Biol. Rev. 2014, 78, 304–341. [Google Scholar] [CrossRef]
- Fulton, J.; Bothner, B.; Lawrence, M.; Johnson, J.E.; Douglas, T.; Young, M. Genetics, biochemistry and structure of the archaeal virus STIV. Biochem. Soc. Trans. 2009, 37, 114–117. [Google Scholar] [CrossRef]
- Zhan, Z.; Ouyang, S.; Liang, W.; Zhang, Z.; Liu, Z.-J.; Huang, L. Biological Crystallography Structural and functional characterization of the C-terminal catalytic domain of SSV1 integrase. Res. Pap. Acta Cryst. 2012, 68, 659–670. [Google Scholar] [CrossRef]
- Eberle, W.; Pastore, A.; Sander, C.; Rösch, P. The structure of ColE1 rop in solution. J. Biomol. NMR 1991, 1, 71–82. [Google Scholar] [CrossRef]
- Clore, A.J.; Stedman, K.M. The SSV1 viral integrase is not essential. Virology 2007, 361, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Jiang, Q.; Woodgate, R.; Cox, M.M.; Goodman, M.F. A New Model for SOS-induced Mutagenesis: How RecA Protein Activates DNA Polymerase V. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Yeats, S.; Janekovic, D.; Reiter, W.-D.; Aicher, W.; Zillig, W. SAV 1, a temperate u.v.-inducible DNA virus-like particle from the archaebacterium Sulfolobus acidocaldarius isolate B12. EMBO J. 1984, 3, 2165–2168. [Google Scholar] [CrossRef] [PubMed]
- Quemin, E.R.J.; Chlanda, P.; Sachse, M.; Forterre, P.; Prangishvili, D.; Krupovic, M. Eukaryotic-like virus budding in archaea. MBio 2016, 7, e01439-16. [Google Scholar] [CrossRef] [PubMed]
- Zink, I.A.; Pfeifer, K.; Wimmer, E.; Sleytr, U.B.; Schuster, B.; Schleper, C. CRISPR-mediated gene silencing reveals involvement of the archaeal S-layer in cell division and virus infection. Nat. Commun. 2019, 10, 4797. [Google Scholar] [CrossRef]
- Rowland, E.F.; Bautista, M.A.; Zhang, C.; Whitaker, R.J. Surface resistance to SSVs and SIRVs in pilin deletions of Sulfolobus islandicus. Mol. Microbiol. 2020, 113, 718–727. [Google Scholar] [CrossRef]
- Pauly, M.D.; Bautista, M.A.; Black, J.A.; Whitaker, R.J. Diversified local CRISPR-Cas immunity to viruses of Sulfolobus islandicus. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180093. [Google Scholar] [CrossRef]
- Koonin, E.V.; Senkevich, T.G.; Dolja, V.V. The ancient virus world and evolution of cells. Biol. Direct 2006, 1, 29. [Google Scholar] [CrossRef]
- Contursi, P.; Farina, B.; Pirone, L.; Fusco, S.; Russo, L.; Bartolucci, S.; Fattorusso, R.; Pedone, E. Structural and functional studies of Stf76 from the Sulfolobus islandicus plasmid-virus pSSVx: A novel peculiar member of the winged helix-turn-helix transcription factor family. Nucleic Acids Res. 2014, 42, 5993–6011. [Google Scholar] [CrossRef]
- Contursi, P.; D’Ambrosio, K.; Pirone, L.; Pedone, E.; Aucelli, T.; She, Q.; De Simone, G.; Bartolucci, S. C68 from the Sulfolobus islandicus plasmid-virus pSSVx is a novel member of the AbrB-like transcription factor family. Biochem. J. 2011, 435, 157–166. [Google Scholar] [CrossRef]
- Prato, S.; Vitale, R.M.; Contursi, P.; Lipps, G.; Saviano, M.; Rossi, M.; Bartolucci, S. Molecular modeling and functional characterization of the monomeric primase-polymerase domain from the Sulfolobus solfataricus plasmid pIT3. FEBS J. 2008, 275, 4389–4402. [Google Scholar] [CrossRef] [PubMed]
- Contursi, P.; Cannio, R.; Prato, S.; She, Q.; Rossi, M.; Bartolucci, S. Transcriptional analysis of the genetic element pSSVx: Differential and temporal regulation of gene expression reveals correlation between transcription and replication. J. Bacteriol. 2007, 189, 6339–6350. [Google Scholar] [CrossRef]
- Stedman, K.M.; Schleper, C.; Rumpf, E.; Zillig, W. Genetic requirements for the function of the archaeal virus SSV1 in Sulfolobus solfataricus: Construction and testing of viral shuttle vectors. Genetics 1999, 152, 1397–1405. [Google Scholar] [CrossRef]
- Cannio, R.; Contursi, P.; Rossi, M.; Bartolucci, S. An autonomously replicating transforming vector for Sulfolobus solfataricus. J. Bacteriol. 1998, 180, 3237–3240. [Google Scholar] [CrossRef]
- Contursi, P.; Cannio, R.; Prato, S.; Fiorentino, G.; Rossi, M.; Bartolucci, S. Development of a genetic system for hyperthermophilic Archaea: Expression of a moderate thermophilic bacterial alcohol dehydrogenase gene in Sulfolobus solfataricus. FEMS Microbiol. Lett. 2003, 218, 115–120. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aulitto, M.; Gallo, G.; Puopolo, R.; Mormone, A.; Limauro, D.; Contursi, P.; Piochi, M.; Bartolucci, S.; Fiorentino, G. Genomic Insight of Alicyclobacillus mali FL18 Isolated From an Arsenic-Rich Hot Spring. Front. Microbiol. 2021, 12, 639697. [Google Scholar] [CrossRef] [PubMed]
- Aulitto, M.; Tom, L.M.; Ceja-Navarro, J.A.; Simmons, B.A.; Singer, S.W. Whole-Genome Sequence of Brevibacillus borstelensis SDM, Isolated from a Sorghum-Adapted Microbial Community. Microbiol. Resour. Announc. 2020, 9, 8–9. [Google Scholar] [CrossRef]
- Aulitto, M.; Strazzulli, A.; Sansone, F.; Cozzolino, F.; Monti, M.; Moracci, M.; Fiorentino, G.; Limauro, D.; Bartolucci, S.; Contursi, P. Prebiotic properties of Bacillus coagulans MA-13: Production of galactoside hydrolyzing enzymes and characterization of the transglycosylation properties of a GH42 β-galactosidase. Microb. Cell Fact. 2021, 20, 71. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, S.; Bartolucci, S.; Franzén, C.J.; Contursi, P. Bacillus coagulans MA-13: A promising thermophilic and cellulolytic strain for the production of lactic acid from lignocellulosic hydrolysate. Biotechnol. Biofuels 2017, 10, 210. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, S.; Nickel, D.B.; Bartolucci, S.; Contursi, P.; Franzén, C.J. Seed culture pre-adaptation of Bacillus coagulans MA-13 improves lactic acid production in simultaneous saccharification and fermentation. Biotechnol. Biofuels 2019, 12, 45. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, S.; Franzén, C.J.; Strazzulli, A.; Moracci, M.; Bartolucci, S.; Contursi, P. Draft Genome Sequence of Bacillus coagulans MA-13, a Thermophilic Lactic Acid Producer from Lignocellulose. Microbiol. Resour. Announc. 2019, 8, 341–360. [Google Scholar] [CrossRef] [PubMed]
- Gallo, G.; Aulitto, M.; Contursi, P.; Limauro, D.; Bartolucci, S.; Fiorentino, G. Bioprospecting of Extremophilic Microorganisms to Address Environmental Pollution. J. Vis. Exp. 2021, 178, e63453. [Google Scholar] [CrossRef] [PubMed]
- Gaglione, R.; Pirone, L.; Farina, B.; Fusco, S.; Smaldone, G.; Aulitto, M.; Dell’Olmo, E.; Roscetto, E.; Del Gatto, A.; Fattorusso, R.; et al. Insights into the anticancer properties of the first antimicrobial peptide from Archaea. Biochim. Biophys. Acta - Gen. Subj. 2017, 1861, 2155–2164. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, S.; Limauro, D.; Fiorentino, G.; Bartolucci, S.; Contursi, P. Galactomannan degradation by thermophilic enzymes: A hot topic for biotechnological applications. World J. Microbiol. Biotechnol. 2019, 35, 32. [Google Scholar] [CrossRef] [PubMed]
- Aulitto, M.; Fusco, F.A.; Fiorentino, G.; Bartolucci, S.; Contursi, P.; Limauro, D. A thermophilic enzymatic cocktail for galactomannans degradation. Enzyme Microb. Technol. 2018, 111, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Ing, N.; Deng, K.; Chen, Y.; Aulitto, M.; Gin, J.W.; Pham, T.L.M.; Petzold, C.J.; Singer, S.W.; Bowen, B.; Sale, K.L.; et al. A multiplexed nanostructure-initiator mass spectrometry (NIMS) assay for simultaneously detecting glycosyl hydrolase and lignin modifying enzyme activities. Sci. Rep. 2021, 11, 11803. [Google Scholar] [CrossRef] [PubMed]
- Varanda, C.; Félix, M.D.R.; Campos, M.D.; Materatski, P. An overview of the application of viruses to biotechnology. Viruses 2021, 13, 13–18. [Google Scholar] [CrossRef]
- Michán, C.; Michán-Doña, A. Viruses: Friends or foes. Microb. Biotechnol. 2022, 15, 88–90. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
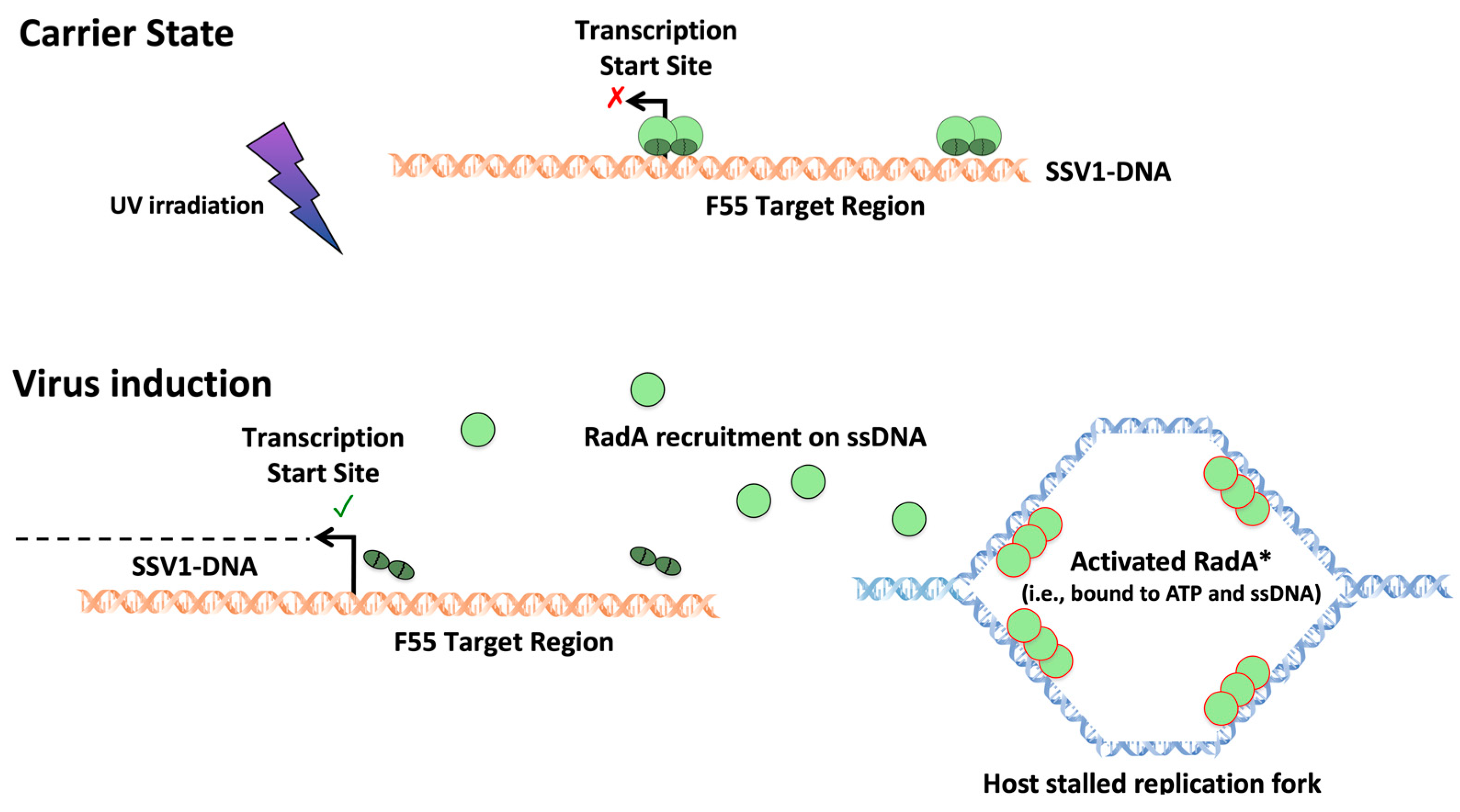{kind=link}
| Protein | Annotation | Core-Gene | Distribution in Crenarchaeal Viruses | Reference |
|---|---|---|---|---|
| F93 | winged helix-turn-helix (wHTH) | N | Fuselloviridae, Portogloboviridae, Guttaviridae | [48] |
| F112 | winged helix-turn-helix (wHTH) | N | unique to SSV1 | [43] |
| B115 | winged helix-turn-helix (wHTH) | Y | Fuselloviridae | |
| F55 | ribbon-helix-helix (RHH) | N | unique to SSV1 | [33,45] |
| C80 | ribbon-helix-helix (RHH) | Y | Lipothrixviridae, Bicaudaviridae, Rudiviridae, Spiraviridae | |
| E51 | ribbon-helix-helix (RHH) | N | Fuselloviridae | |
| A79 | zinc-finger | Y | Fuselloviridae, Rudiviridae, Portogloboviridae, Ovaloviridae | |
| B129 | zinc-finger | Y | Fuselloviridae | [44] |
| A45 | zinc-finger | N | Fuselloviridae, Lipothrixviridae | |
| C102 | zinc-finger, putative membrane protein | Y | Fuselloviridae, Guttaviridae | |
| A100 | Pur-alpha repeat | Y | Fuselloviridae, Rudiviridae, Portogloboviridae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aulitto, M.; Martinez-Alvarez, L.; Fusco, S.; She, Q.; Bartolucci, S.; Peng, X.; Contursi, P. Genomics, Transcriptomics, and Proteomics of SSV1 and Related Fusellovirus: A Minireview. Viruses 2022, 14, 2082. https://doi.org/10.3390/v14102082
Aulitto M, Martinez-Alvarez L, Fusco S, She Q, Bartolucci S, Peng X, Contursi P. Genomics, Transcriptomics, and Proteomics of SSV1 and Related Fusellovirus: A Minireview. Viruses. 2022; 14(10):2082. https://doi.org/10.3390/v14102082
Chicago/Turabian StyleAulitto, Martina, Laura Martinez-Alvarez, Salvatore Fusco, Qunxin She, Simonetta Bartolucci, Xu Peng, and Patrizia Contursi. 2022. "Genomics, Transcriptomics, and Proteomics of SSV1 and Related Fusellovirus: A Minireview" Viruses 14, no. 10: 2082. https://doi.org/10.3390/v14102082
APA StyleAulitto, M., Martinez-Alvarez, L., Fusco, S., She, Q., Bartolucci, S., Peng, X., & Contursi, P. (2022). Genomics, Transcriptomics, and Proteomics of SSV1 and Related Fusellovirus: A Minireview. Viruses, 14(10), 2082. https://doi.org/10.3390/v14102082