
Comprehensive Analysis of Codon Usage Patterns in Chinese Porcine Circoviruses Based on Their Major Protein-Coding Sequences

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Selection
2.2. Composition Analysis of the Selected PCV Strains
2.3. Relative Synonymous Codon Usage (RSCU) Value and Principal Component Analysis (PCA) Analysis
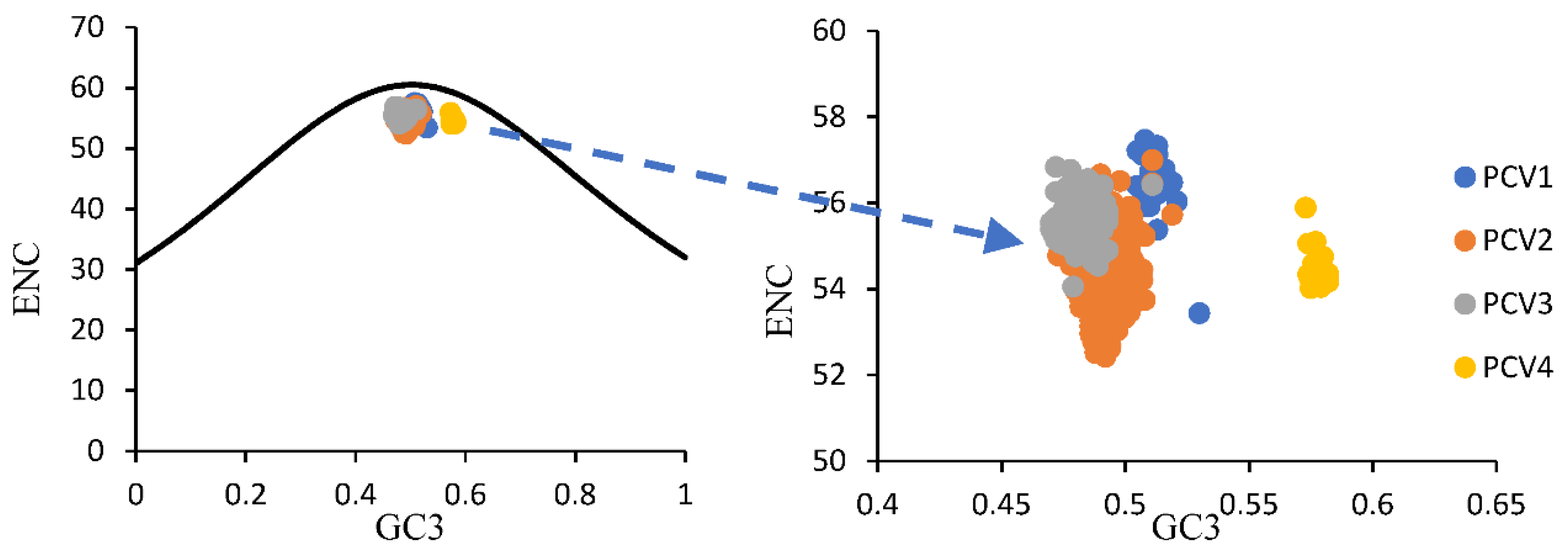
2.4. Effective Number of Codons (ENC) and ENC-Plot
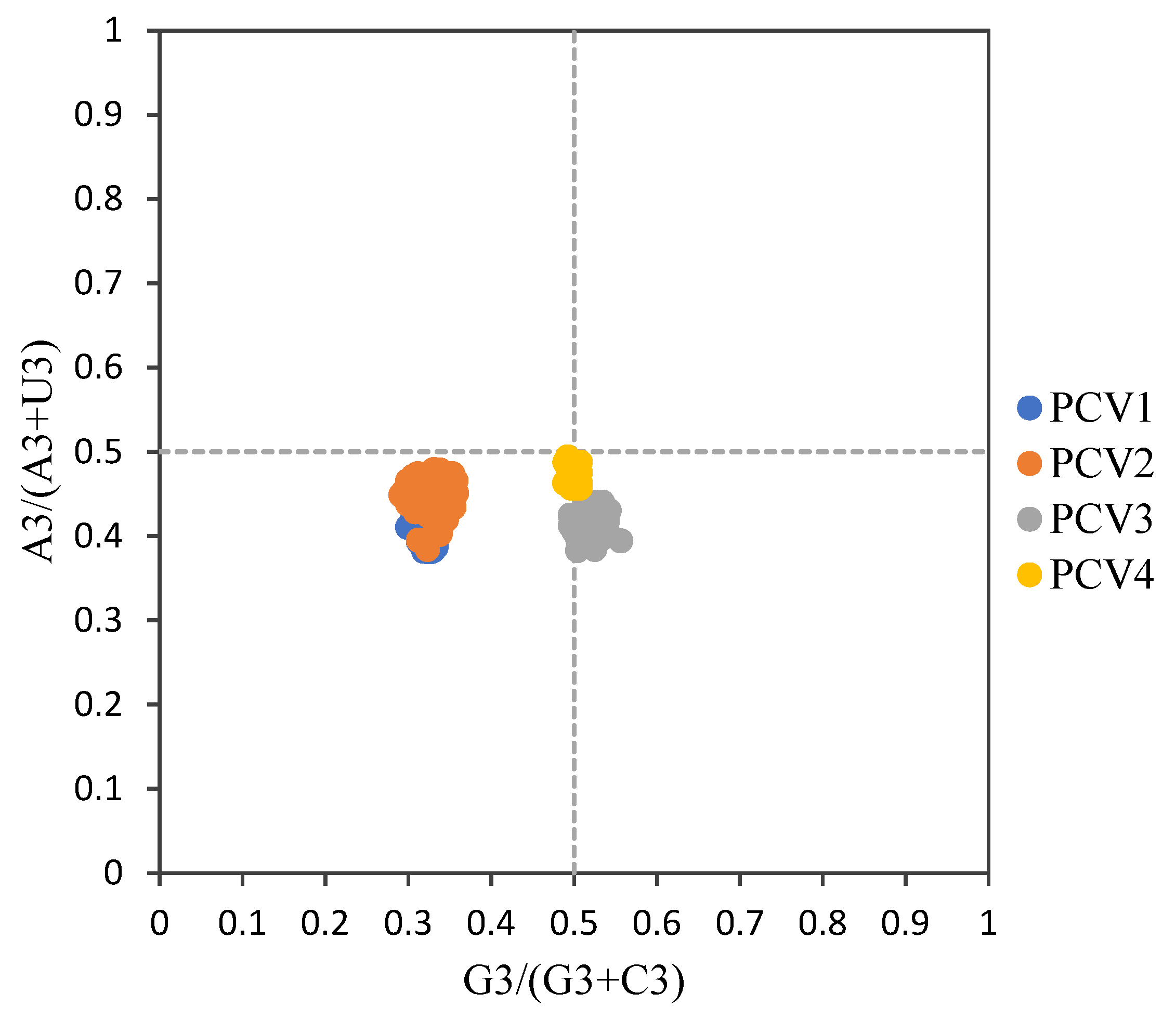
2.5. Parity Rule 2 (PR2) Analysis
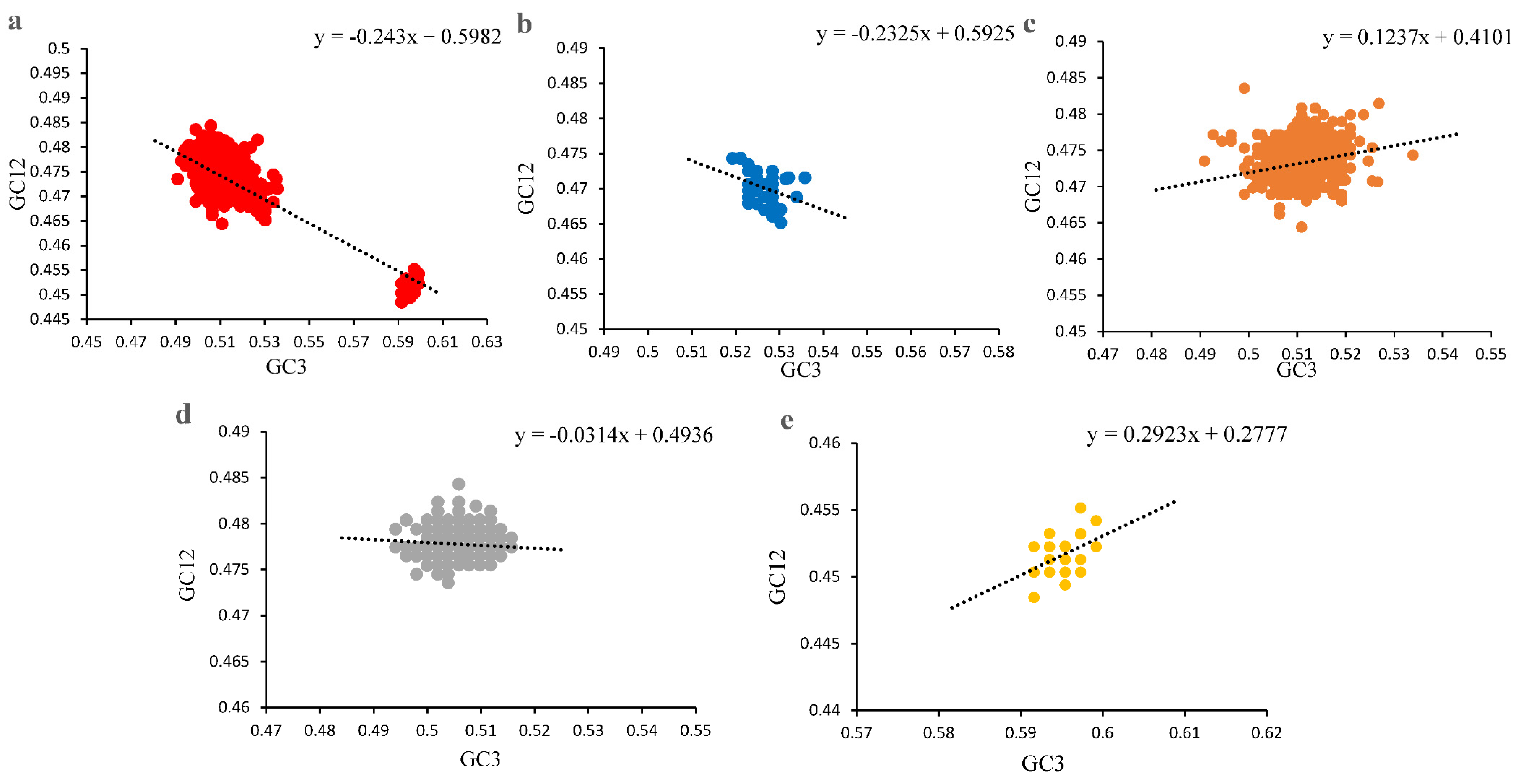
2.6. Neutrality Plot Analysis
2.7. Codon Adaptation Index (CAI) Analysis
2.8. Hydropathicity and Aromaticity Analysis
2.9. Correlation Analysis
3. Results
3.1. Characteristics of Nucleotide Composition of PCVs
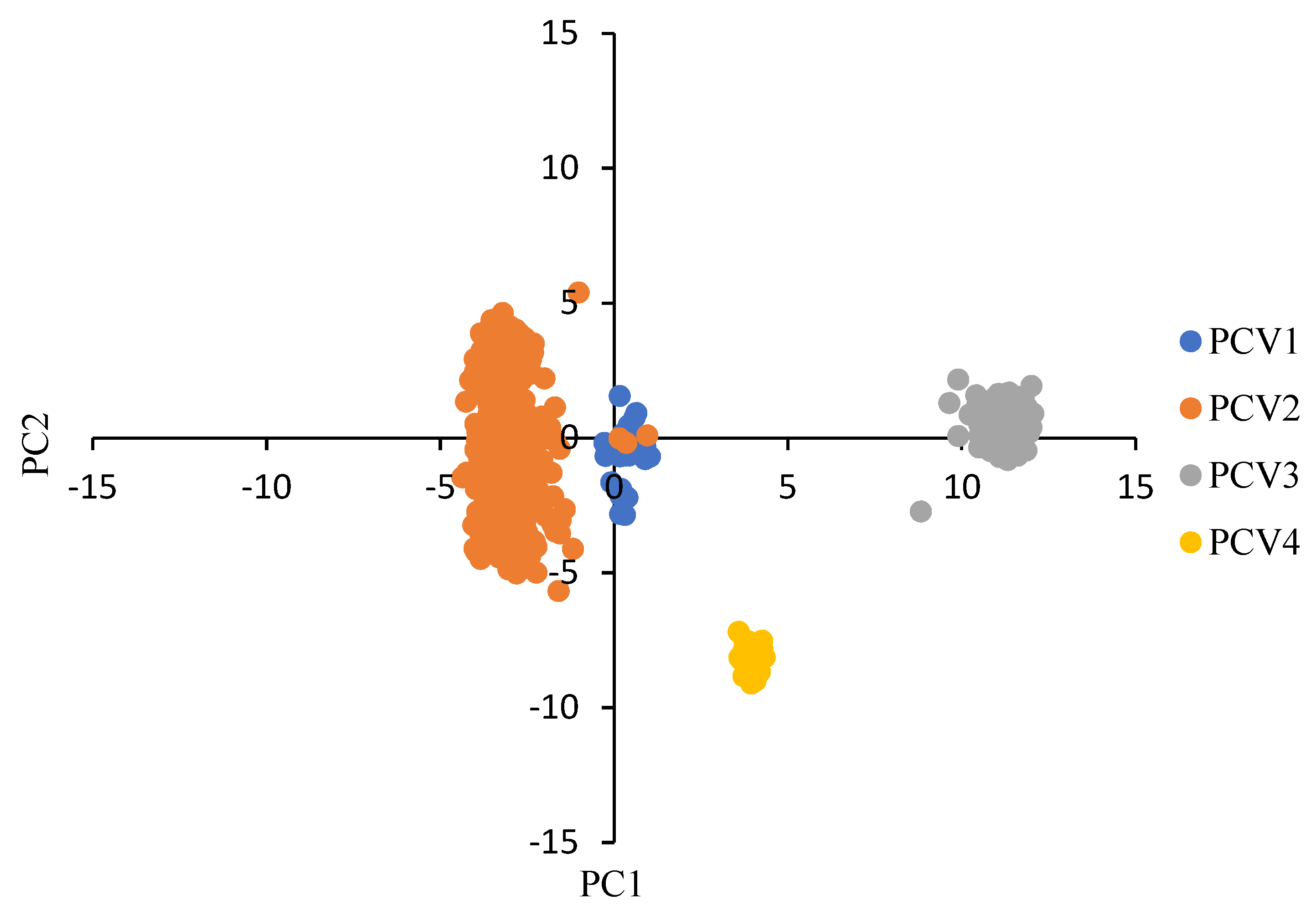
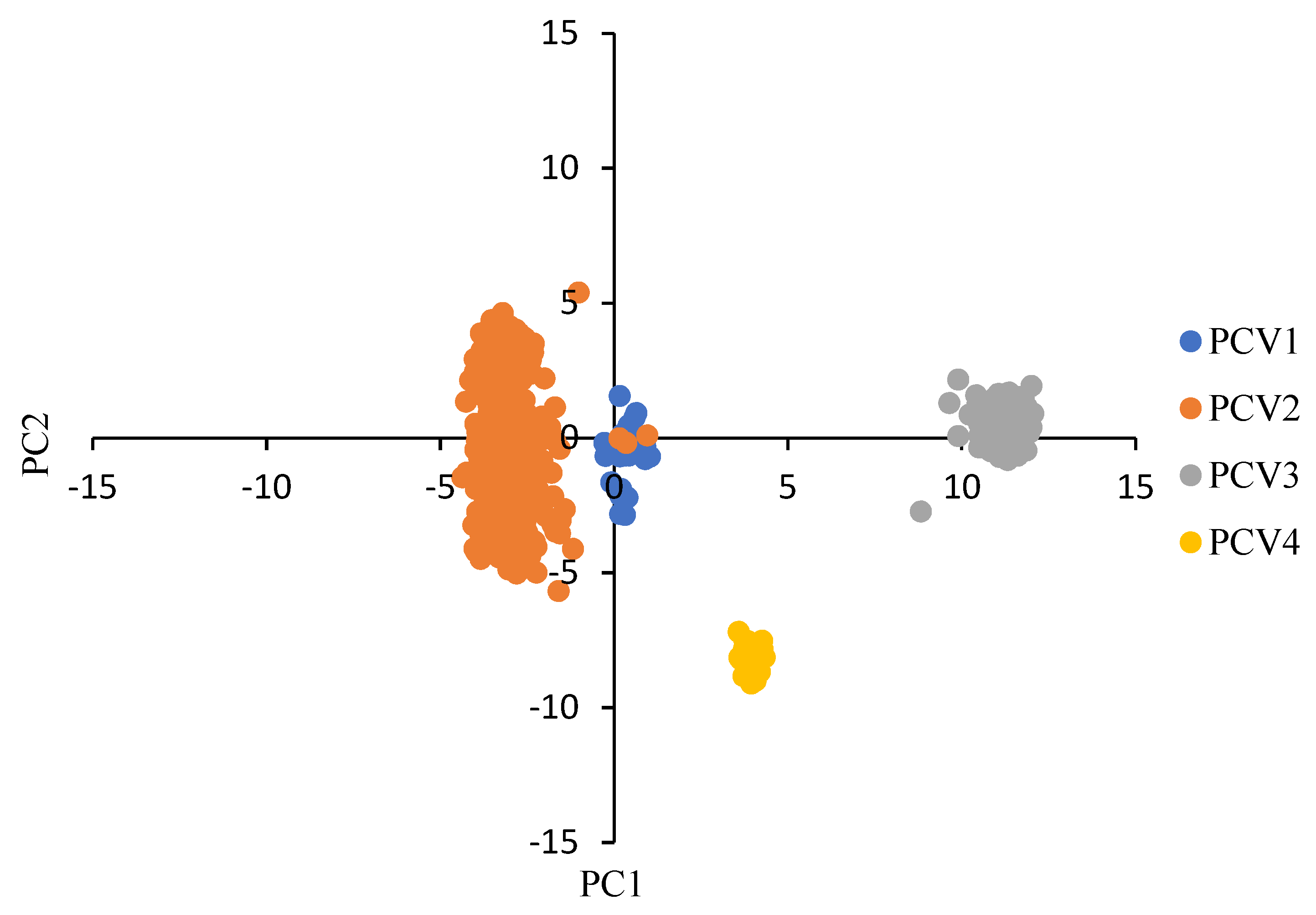
3.2. Characteristics of the Codon Usage Bias of PCVs
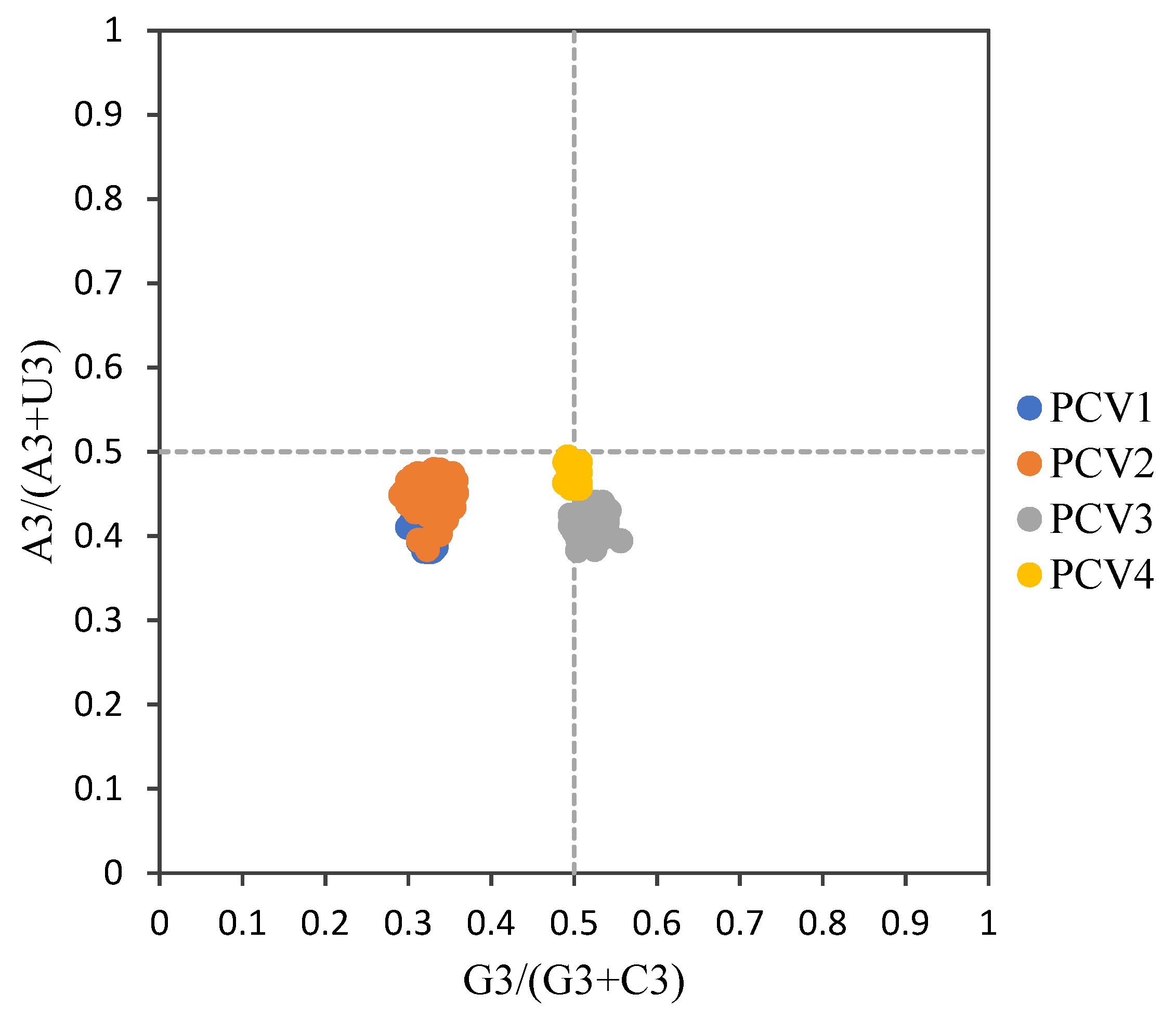
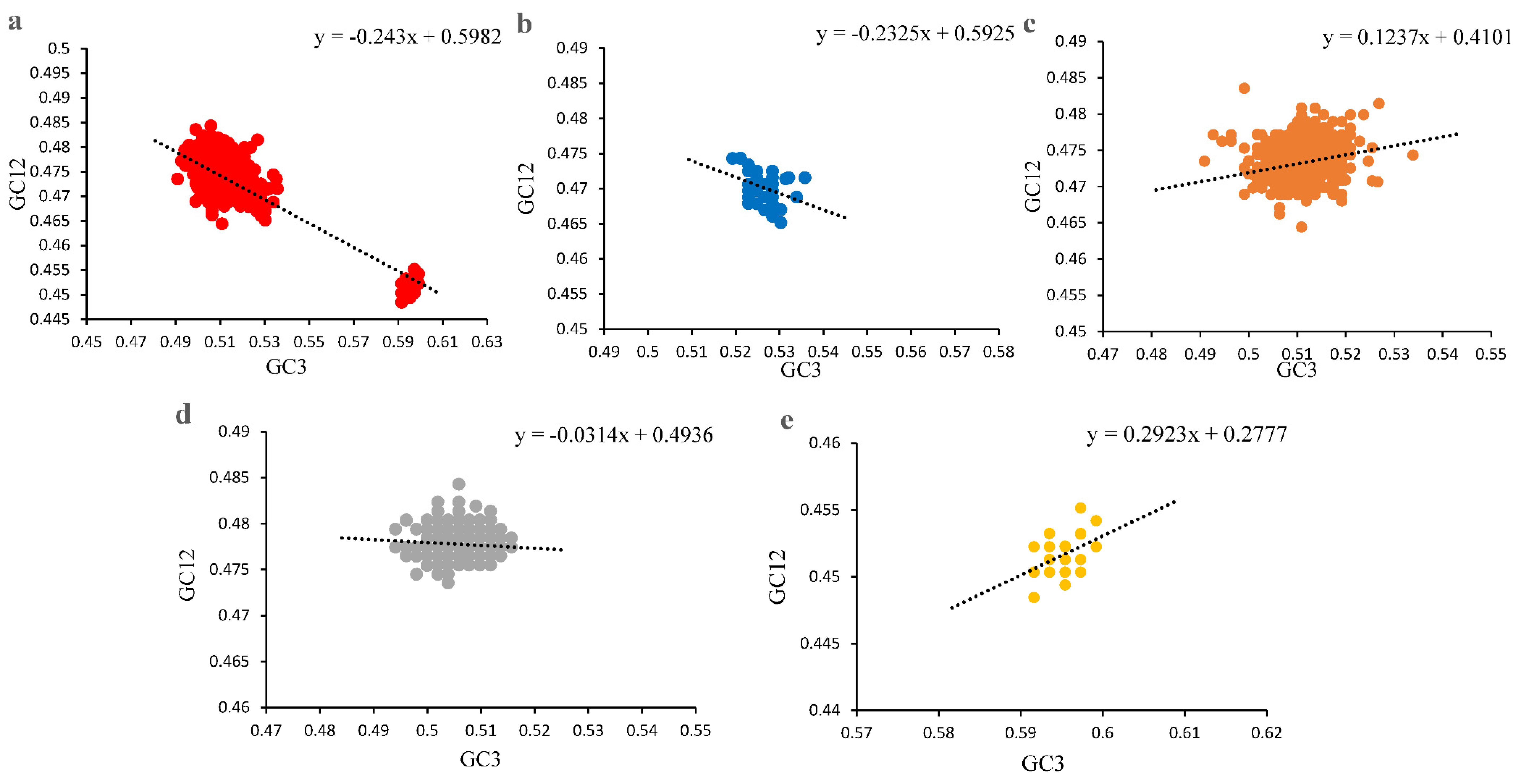
3.3. Codon Usage Bias of PCVs Is Influenced Mainly by Natural Selection
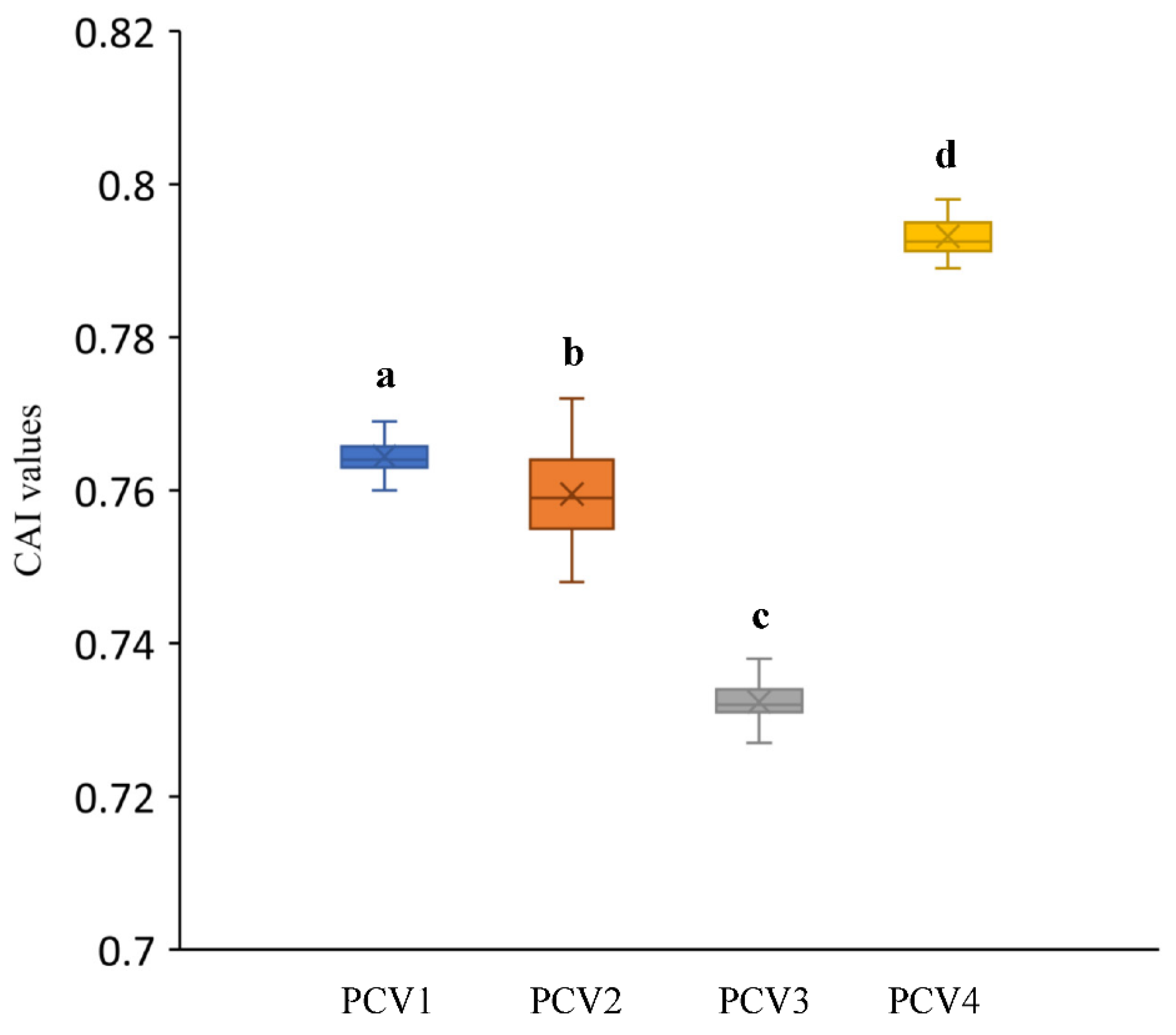
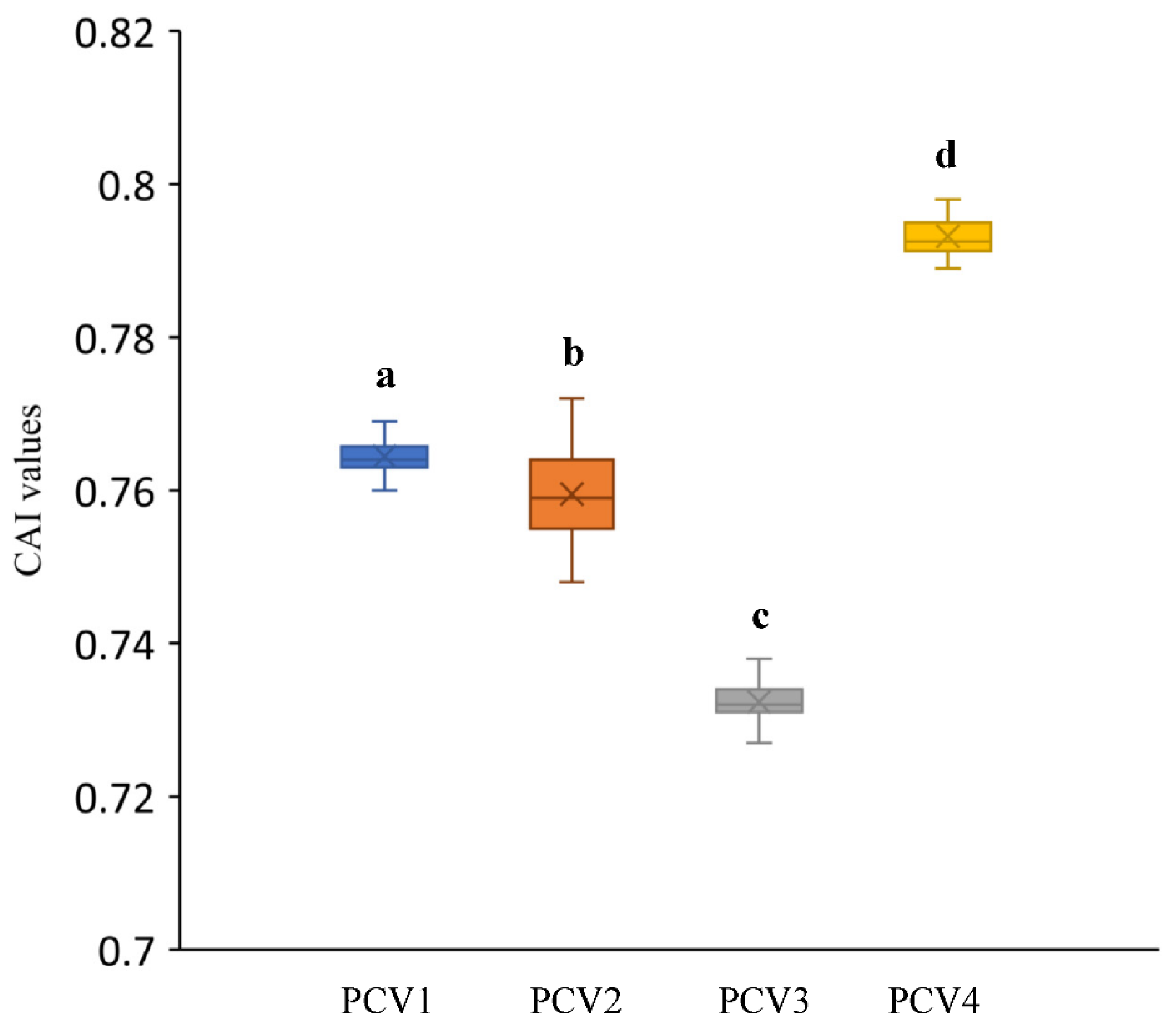
3.4. Codon Adaptation Index Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tischer, I.; Glederblom, H.; Vettermann, W.; Koch, M.A. A very small porcine virus with circular single stranded DNA. Nature 1982, 295, 64–66. [Google Scholar] [CrossRef]
- Rakibuzzaman, A.; Ramamoorthy, S. Comparative immunopathogenesis and biology of recently discovered porcine circoviruses. Transbound. Emerg. Dis. 2021, 68, 2957–2968. [Google Scholar] [CrossRef]
- Opriessnig, T.; Karuppannan, A.K.; Castro, A.; Xiao, C.T. Porcine circoviruses: Current status, knowledge gaps and challenges. Virus Res. 2020, 286, 198044. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J. Porcine circovirus: A historical perspective. Vet. Pathol. 2014, 51, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.L.; Lu, S.S.; Wei, W.K.; Lv, D.H.; Wen, X.H.; Zhai, Q.; Chen, Q.L.; Sun, Y.W.; Xi, Y. Reservoirs of Porcine Circoviruses: A Mini Review. Front. Vet. Sci. 2019, 6, 319. [Google Scholar] [CrossRef] [Green Version]
- Zhai, S.L.; Chen, S.N.; Xu, Z.H.; Tang, M.H.; Wang, F.G.; Li, X.J.; Sun, B.B.; Deng, S.F.; Hu, J.; Lv, D.H.; et al. Porcine circovirus type 2 in China: An update on and insights to its prevalence and control. Virol. J. 2014, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Allan, G.M.; McNeilly, F.; Cassidy, J.P.; Reilly, G.A.; Adair, B.; Ellis, W.A.; McNulty, M.S. Pathogenesis of porcine circovirus; experimental infections of colostrum deprived piglets and examination of pig foetal material. Vet. Microbiol. 1995, 44, 49–64. [Google Scholar] [CrossRef]
- Tischer, I.; Mields, W.; Wolff, D.; Vagt, M.; Griem, W. Studies on epidemiology and pathogenicity of porcine circovirus. Arch. Virol. 1986, 91, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J. Best practice and future challenges for vaccination against porcine circovirus type 2. Expert Rev. Vaccines 2015, 14, 473–487. [Google Scholar] [CrossRef]
- Jiang, C.G.; Wang, G.; Tu, Y.B.; Liu, Y.G.; Wang, S.J.; Cai, X.H.; An, T.Q. Genetic analysis of porcine circovirus type 2 in China. Arch. Virol. 2017, 162, 2715–2726. [Google Scholar] [CrossRef]
- Phan, T.G.; Giannitti, F.; Rossow, S.; Marthaler, D.; Knutson, T.; Li, L.; Deng, X.; Resende, T.; Vannucci, F.; Delwart, E. Detection of a novel circovirus PCV3 in pigs with cardiac and multi-systemic inflammation. Virol. J. 2016, 13, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palinski, R.; Pineyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A novel porcine circovirus distantly related to known circoviruses is associated with porcine dermatitis and nephropathy syndrome and reproductive failure. J. Virol. 2016, 91, e01879-16. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, T.; Niu, G.; Liu, X.; Zhang, X.; Zhang, Y.; Ren, L. Recent progress on porcine circovirus type 3. Infect. Genet. Evol. 2019, 73, 227–233. [Google Scholar] [CrossRef]
- Zhang, H.H.; Hu, W.Q.; Li, J.Y.; Liu, T.N.; Zhou, J.Y.; Opriessnig, T.; Xiao, C.T. Novel circovirus species identified in farmed pigs designated as Porcine circovirus 4, Hunan province, China. Transbound. Emerg. Dis. 2020, 67, 1057–1061. [Google Scholar] [CrossRef]
- Segalés, J. Porcine circovirus type 2 (PCV2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef]
- Meehan, B.M.; McNeilly, F.; Todd, D.; Kennedy, S.; Jewhurst, V.A.; Ellis, J.A.; Hassard, L.E.; Clark, E.G.; Haines, D.M.; Allan, G.M. Characterization of novel circovirus DNAs associated with wasting syndromes in pigs. J. Gen. Virol. 1998, 79 Pt 9, 2171–2179. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 associated disease: Update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J. Vet. Diagn. Investig. 2007, 19, 591–615. [Google Scholar] [CrossRef]
- Butt, A.M.; Nasrullah, I.; Qamar, R.; Tong, Y. Evolution of codon usage in Zika virus genomes is host and vector specific. Emerg. Microbes Infect. 2016, 5, e107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershberg, R.; Petrov, D.A. Selection on codon bias. Annu. Rev. Genet. 2008, 42, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Andersson, S.G.; Kurland, C.G. Codon preferences in free-living microorganisms. Microbiol. Rev. 1990, 54, 198–210. [Google Scholar] [CrossRef]
- Behura, S.K.; Severson, D.W. Codon usage bias: Causative factors, quantification methods and genome-wide patterns: With emphasis on insect genomes. Biol. Rev. Camb. Philos. Soc. 2013, 88, 49–61. [Google Scholar] [CrossRef]
- Rahman, S.U.; Yao, X.; Li, X.; Chen, D.; Tao, S. Analysis of codon usage bias of Crimean-Congo hemorrhagic fever virus and its adaptation to hosts. Infect. Genet. Evol. 2018, 58, 1–16. [Google Scholar] [CrossRef]
- Yao, X.; Fan, Q.; Yao, B.; Lu, P.; Rahman, S.U.; Chen, D.; Tao, S. Codon Usage Bias Analysis of Bluetongue Virus Causing Livestock Infection. Front. Microbiol. 2020, 11, 655. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Puigbo, P.; Bravo, I.G.; Garcia-Vallve, S. CAIcal: A combined set of tools to assess codon usage adaptation. Biol. Direct. 2008, 3, 38. [Google Scholar] [CrossRef] [Green Version]
- Athey, J.; Alexaki, A.; Osipova, E.; Rostovtsev, A.; Santana-Quintero, L.V.; Katneni, U.; Simonyan, V.; Kimchi-Sarfaty, C. A new and updated resource for codon usage tables. BMC Bioinform. 2017, 18, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischer, I.R.; Tochtermann, G.R. Characterization of papovavirus-and picornavirus-like particles in permanent pig kidney cell lines. Zent. Fur Bakteriol. Parasitenkd. Infekt. Und Hyg. Erste Abt. Orig. Reihe A Med. Mikrobiol. Und Parasitol. 1974, 226, 153–167. [Google Scholar]
- Pinto, R.M.; Perez-Rodriguez, F.J.; D’Andrea, L.; de Castellarnau, M.; Guix, S.; Bosch, A. Hepatitis A Virus Codon Usage: Implications for Translation Kinetics and Capsid Folding. Cold Spring Harb. Perspect. Med. 2018, 8, a031781. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.R.; Papamichail, D.; Skiena, S.; Futcher, B.; Wimmer, E.; Mueller, S. Virus attenuation by genome-scale changes in codon pair bias. Science 2008, 320, 1784–1787. [Google Scholar] [CrossRef] [Green Version]
- Kekarainen, T.; Gonzalez, A.; Llorens, A.; Segalés, J. Genetic variability of porcine circovirus 2 in vaccinating and non-vaccinating commercial farms. J. Gen. Virol. 2014, 95, 1734–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Sun, J.; Tong, X.; Xu, J.; Deng, H.; Jiang, Z.; Jiang, C.; Duan, J.; Li, J.; Zhou, P.; et al. First analysis of synonymous codon usage in porcine circovirus. Arch. Virol. 2014, 159, 2145–2151. [Google Scholar] [CrossRef]
- Liu, X.S.; Zhang, Y.G.; Fang, Y.Z.; Wang, Y.L. Patterns and influencing factor of synonymous codon usage in porcine circovirus. Virol. J. 2012, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Gao, K.; Pi, M.; Li, H.; Zhong, W.; Li, B.; Ning, Z. Phylogenetic and codon usage analysis for replicase and capsid genes of porcine circovirus 3. Vet. Res. Commun. 2021, 45, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Segales, J.; Tucciarone, C.M.; Cecchinato, M.; Drigo, M. The analysis of genome composition and codon bias reveals distinctive patterns between avian and mammalian circoviruses which suggest a potential recombinant origin for Porcine circovirus 3. PLoS ONE 2018, 13, e0199950. [Google Scholar] [CrossRef]
- Jia, W.; Higgs, P.G. Codon usage in mitochondrial genomes: Distinguishing context-dependent mutation from translational selection. Mol. Biol. Evol. 2008, 25, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, Q.; Zhao, F. Synonymous but Not Silent: The Codon Usage Code for Gene Expression and Protein Folding. Annu. Rev. Biochem. 2021, 90, 375–401. [Google Scholar] [CrossRef]
- Butt, A.M.; Nasrullah, I.; Tong, Y. Genome-wide analysis of codon usage and influencing factors in chikungunya viruses. PLoS ONE 2014, 9, e90905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzo, G.; Ruiz, A.; Grassi, L.; Sibila, M.; Drigo, M.; Segales, J. Lack of Porcine circovirus 4 Genome Detection in Pig Samples from Italy and Spain. Pathogens 2020, 9, 433. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Kim, H.R.; Park, J.H.; Kwon, N.Y.; Kim, J.M.; Kim, J.K.; Park, J.H.; Lee, K.K.; Kim, S.H.; Kim, W.I.; et al. Detection of a novel porcine circovirus 4 in Korean pig herds using a loop-mediated isothermal amplification assay. J. Virol. Methods 2021, 299, 114350. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catalogs | PCV | PCV-1 | PCV-2 | PCV-3 | PCV-4 |
|---|---|---|---|---|---|
| A% | 28.01 ± 0.27 | 27.47 ± 0.15 | 28.11 ± 0.17 | 27.67 ± 0.13 | 28.61 ± 0.11 |
| C% | 24.46 ± 1.64 | 25.58 ± 0.11 | 25.32 ± 0.19 | 21.36 ± 0.15 | 22.28 ± 0.12 |
| U% | 23.35 ± 0.32 | 23.61 ± 0.12 | 23.31 ± 0.17 | 23.61 ± 0.11 | 21.44 ± 0.09 |
| G% | 24.18 ± 1.72 | 23.34 ± 0.19 | 23.27 ± 0.14 | 27.37 ± 0.13 | 27.67 ± 0.07 |
| A3% | 21.84 ± 0.66 | 20.42 ± 0.40 | 21.95 ± 0.37 | 21.91 ± 0.37 | 17.95 ± 0.24 |
| C3% | 28.68 ± 2.83 | 29.99 ± 0.38 | 30.12 ± 0.36 | 23.17 ± 0.39 | 30.06 ± 0.19 |
| U3% | 26.98 ± 0.74 | 26.86 ± 0.26 | 26.94 ± 0.45 | 27.46 ± 0.32 | 22.55 ± 0.23 |
| G3% | 22.50 ± 2.77 | 22.73 ± 0.32 | 20.99 ± 0.34 | 27.45 ± 0.29 | 29.45 ± 0.18 |
| GC% | 48.65 ± 0.28 | 48.92 ± 0.20 | 48.59 ± 0.22 | 48.72 ± 0.17 | 49.95 ± 0.14 |
| GC1% | 50.15 ± 0.88 | 50.03 ± 0.37 | 50.59 ± 0.29 | 48.52 ± 0.19 | 50.67 ± 0.16 |
| GC2% | 44.61 ± 1.38 | 43.99 ± 0.25 | 44.07 ± 0.26 | 47.03 ± 0.25 | 39.65 ± 0.21 |
| GC3% | 51.18 ± 1.19 | 52.72 ± 0.44 | 51.10 ± 0.40 | 50.62 ± 0.45 | 59.51 ± 0.20 |
| GC12% | 47.38 ± 0.40 | 47.01 ± 0.23 | 47.33 ± 0.22 | 47.77 ± 0.14 | 45.16 ± 0.15 |
| AA | Codons | PCV-1 | PCV-2 | PCV-3 | PCV-4 | Susscrofa |
|---|---|---|---|---|---|---|
| Phe | UUU | 1.13 | 1.15 | 1.08 | 0.87 | 0.93 |
| UUC | 0.87 | 0.85 | 0.92 | 1.13 | 1.07 | |
| Leu | UUA | 0.27 | 0.16 | 0.38 | 0.18 | 0.49 |
| UUG | 1.64 | 1.07 | 1.54 | 0.71 | 0.78 | |
| CUU | 1.25 | 1.25 | 0.59 | 0.87 | 0.81 | |
| CUC | 1.22 | 1.24 | 1.30 | 1.40 | 1.18 | |
| CUA | 0.65 | 0.77 | 0.76 | 0.51 | 0.39 | |
| CUG | 0.96 | 1.51 | 1.44 | 2.33 | 2.35 | |
| Ile | AUU | 0.96 | 1.19 | 2.01 | 1.12 | 1.06 |
| AUC | 1.20 | 1.01 | 0.14 | 1.27 | 1.42 | |
| AUA | 0.85 | 0.79 | 0.86 | 0.61 | 0.52 | |
| Val | GUU | 1.14 | 0.82 | 2.20 | 0.41 | 0.73 |
| GUC | 0.46 | 0.81 | 0.44 | 1.10 | 0.98 | |
| GUA | 1.29 | 1.19 | 0.62 | 0.33 | 0.45 | |
| GUG | 1.12 | 1.18 | 0.73 | 2.15 | 1.83 | |
| Ser | UCU | 0.99 | 0.45 | 0.81 | 1.13 | 1.09 |
| UCC | 2.01 | 2.63 | 1.24 | 1.03 | 1.30 | |
| UCA | 0.42 | 0.35 | 0.20 | 0.37 | 0.87 | |
| UCG | 0.19 | 0.01 | 0.93 | 0.00 | 0.37 | |
| AGU | 0.80 | 1.28 | 0.62 | 1.26 | 0.91 | |
| AGC | 1.59 | 1.27 | 2.21 | 2.19 | 1.46 | |
| Pro | CCU | 0.98 | 0.94 | 1.03 | 0.34 | 1.12 |
| CCC | 1.68 | 1.60 | 0.90 | 1.53 | 1.31 | |
| CCA | 1.02 | 1.12 | 1.22 | 1.36 | 1.06 | |
| CCG | 0.32 | 0.35 | 0.85 | 0.76 | 0.51 | |
| Thr | ACU | 1.35 | 1.24 | 1.13 | 1.58 | 0.94 |
| ACC | 1.66 | 1.51 | 1.39 | 1.28 | 1.41 | |
| ACA | 0.46 | 0.70 | 1.05 | 0.79 | 1.10 | |
| ACG | 0.52 | 0.55 | 0.43 | 0.35 | 0.55 | |
| Ala | GCU | 1.57 | 2.28 | 1.24 | 1.00 | 1.02 |
| GCC | 1.59 | 0.82 | 0.92 | 1.55 | 1.64 | |
| GCA | 0.42 | 0.42 | 0.64 | 0.55 | 0.89 | |
| GCG | 0.42 | 0.48 | 1.20 | 0.90 | 0.45 | |
| Tyr | UAU | 1.05 | 0.93 | 1.22 | 1.12 | 0.87 |
| UAC | 0.95 | 1.07 | 0.78 | 0.88 | 1.13 | |
| His | CAU | 0.53 | 0.65 | 0.19 | 0.90 | 0.80 |
| CAC | 1.47 | 1.35 | 1.81 | 1.10 | 1.20 | |
| Gln | CAA | 0.73 | 0.84 | 1.34 | 0.93 | 0.50 |
| CAG | 1.27 | 1.16 | 0.66 | 1.07 | 1.50 | |
| Asn | AAU | 1.23 | 1.39 | 0.87 | 0.73 | 0.93 |
| AAC | 0.77 | 0.61 | 1.13 | 1.27 | 1.07 | |
| Lys | AAA | 1.03 | 1.16 | 1.17 | 0.90 | 0.87 |
| AAG | 0.97 | 0.84 | 0.83 | 1.10 | 1.13 | |
| Asp | GAU | 1.00 | 1.06 | 1.22 | 0.90 | 0.92 |
| GAC | 1.00 | 0.94 | 0.78 | 1.10 | 1.08 | |
| Glu | GAA | 1.03 | 1.30 | 0.90 | 0.74 | 0.86 |
| GAG | 0.97 | 0.70 | 1.10 | 1.26 | 1.14 | |
| Cys | UGU | 1.12 | 0.87 | 1.14 | 1.46 | 0.91 |
| UGC | 0.88 | 1.13 | 0.86 | 0.54 | 1.09 | |
| Arg | CGU | 0.67 | 0.51 | 0.76 | 0.57 | 0.45 |
| CGC | 1.32 | 1.93 | 0.78 | 0.75 | 1.05 | |
| CGA | 0.28 | 0.36 | 0.36 | 0.00 | 0.67 | |
| CGG | 0.65 | 0.50 | 0.90 | 1.50 | 1.24 | |
| AGA | 2.05 | 1.83 | 1.67 | 1.68 | 1.29 | |
| AGG | 1.04 | 0.88 | 1.53 | 1.50 | 1.30 | |
| Gly | GGU | 0.85 | 0.79 | 0.88 | 0.56 | 0.63 |
| GGC | 1.16 | 1.18 | 0.57 | 1.15 | 1.36 | |
| GGA | 1.09 | 1.11 | 1.02 | 1.33 | 1.00 | |
| GGG | 0.91 | 0.92 | 1.53 | 0.96 | 1.01 |
| Preferred Codons Ended with G/C | Preferred Codons Ended with A/U | Preferred Codons Commonly Used by PCVs and Sus scrofa | |
|---|---|---|---|
| PCV1 | AUC, UCC, CCC, ACC, GCC, CAC, GAC, GGC, UUG, CAG | GUA, AAA, AGA, GAA, UUU, UAU, AAU, UGU | AUC, CCC, ACC, GCC, CAC, CAG, GAC, GGC |
| PCV2 | UCC, CCC, ACC, UAC, CAC, UGC, CGC, GGC, CUG, CAG | GUA, AAA, GAA, UUU, AUU, GUU, AAU, GAU | CUG, CCC, ACC, UAC, CAC, CAG, GGC, UGC |
| PCV3 | AGC, ACC, CAC, AAC, UUG, GAG, GGG | CCA, CAA, AAA, AGA, UUU, AUU, GUU, GCU, UAU, GAU, UGU | AGC, ACC, CAC, AAC, GAG |
| PCV4 | UUC, AUC, AGC, CCC, GCC, CAC, AAC, GAC, CUG, GUG, CAG, AAG, GAG | AGA, GGA, ACU, UAU, UGU | UUC, CUG, AUC, GUG, AGC, CCC, GCC, CAC, CAG, AAC, AAG, GAC, GAG |
| A | C | U | G | GC | Gravy | Aroma | |
|---|---|---|---|---|---|---|---|
| PCV-1-ORF12 | |||||||
| A3 | 0.52 ** | 0.13 | −0.20 | −0.36 * | −0.25 | −0.22 | −0.11 |
| C3 | 0.13 | 0.74 ** | −0.29 | −0.33 * | 0.10 | 0.12 | 0.37 * |
| U3 | −0.20 | −0.56 ** | 0.66 ** | 0.00 | −0.30 | −0.04 | 0.02 |
| G3 | −0.20 | −0.46 ** | 0.04 | 0.42 ** | 0.13 | 0.09 | −0.05 |
| GC3 | −0.17 | 0.34 * | −0.28 | 0.14 | 0.33 * | 0.19 | 0.19 |
| ENC | −0.30 * | −0.21 | 0.55 ** | −0.06 | −0.10 | 0.29 | 0.44 ** |
| PCV-2-ORF12 | |||||||
| A3 | 0.41 ** | 0.35 ** | −0.56 ** | −0.42 ** | 0.06 * | −0.20 ** | 0.31 ** |
| C3 | −0.13 ** | 0.77 ** | −0.56 ** | −0.26 ** | 0.50 ** | −0.13 ** | 0.27 ** |
| U3 | 0.15 ** | −0.74 ** | 0.87 ** | −0.08 ** | −0.73 ** | 0.18 ** | −0.32 ** |
| G3 | −0.31 ** | −0.37 ** | 0.21 ** | 0.68 ** | 0.09 ** | 0.08 ** | −0.12 ** |
| GC3 | −0.53 ** | 0.53 ** | −0.47 ** | 0.42 ** | 0.76 ** | −0.06 * | 0.18 ** |
| ENC | 0.24 ** | −0.41 ** | 0.38 ** | 0.03 | −0.38 ** | 0.29 ** | −0.58 ** |
| PCV-3-ORF12 | |||||||
| A3 | 0.78 ** | −0.20 ** | −0.07 | −0.48 ** | −0.55 ** | −0.28 ** | 0.07 |
| C3 | −0.28 ** | 0.78 ** | −0.55 ** | −0.18 ** | 0.61 ** | 0.12 * | −0.11 * |
| U3 | −0.10 | −0.59 ** | 0.82 ** | 0.10 | −0.48 ** | 0.18 ** | 0.11 * |
| G3 | −0.43 ** | −0.33 ** | 0.10 | 0.80 ** | 0.25 ** | 0.16 ** | 0.05 |
| GC3 | −0.57 ** | 0.57 ** | −0.48 ** | 0.32 ** | 0.79 ** | 0.15 ** | −0.11 * |
| ENC | 0.03 | 0.37 ** | −0.28 ** | −0.25 ** | 0.19 ** | −0.18 ** | −0.11 * |
| PCV-4-ORF12 | |||||||
| A3 | 0.60 ** | −0.18 | −0.35 | −0.21 | −0.30 | −0.05 | 0.15 |
| C3 | −0.46 * | 0.73 ** | −0.58 ** | 0.16 | 0.77 ** | 0.41 * | 0.05 |
| U3 | −0.08 | −0.47 * | 0.88 ** | −0.15 | −0.48 * | −0.03 | 0.39 * |
| G3 | 0.09 | −0.31 | 0.23 | 0.16 | −0.21 | −0.32 | −0.34 |
| GC3 | −0.50 ** | 0.56 ** | −0.43 * | 0.39 * | 0.71 ** | 0.17 | −0.41 * |
| ENC | 0.28 | −0.08 | 0.13 | −0.39 * | −0.22 | −0.25 | −0.42 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, H.; Segalés, J.; Wang, F.; Jin, Q.; Wang, A.; Zhang, G.; Franzo, G. Comprehensive Analysis of Codon Usage Patterns in Chinese Porcine Circoviruses Based on Their Major Protein-Coding Sequences. Viruses 2022, 14, 81. https://doi.org/10.3390/v14010081
Feng H, Segalés J, Wang F, Jin Q, Wang A, Zhang G, Franzo G. Comprehensive Analysis of Codon Usage Patterns in Chinese Porcine Circoviruses Based on Their Major Protein-Coding Sequences. Viruses. 2022; 14(1):81. https://doi.org/10.3390/v14010081
Chicago/Turabian StyleFeng, Hua, Joaquim Segalés, Fangyu Wang, Qianyue Jin, Aiping Wang, Gaiping Zhang, and Giovanni Franzo. 2022. "Comprehensive Analysis of Codon Usage Patterns in Chinese Porcine Circoviruses Based on Their Major Protein-Coding Sequences" Viruses 14, no. 1: 81. https://doi.org/10.3390/v14010081
APA StyleFeng, H., Segalés, J., Wang, F., Jin, Q., Wang, A., Zhang, G., & Franzo, G. (2022). Comprehensive Analysis of Codon Usage Patterns in Chinese Porcine Circoviruses Based on Their Major Protein-Coding Sequences. Viruses, 14(1), 81. https://doi.org/10.3390/v14010081