
An Absolutely Conserved Tryptophan in the Stem of the Envelope Protein E of Flaviviruses Is Essential for the Formation of Stable Particles


Abstract
:
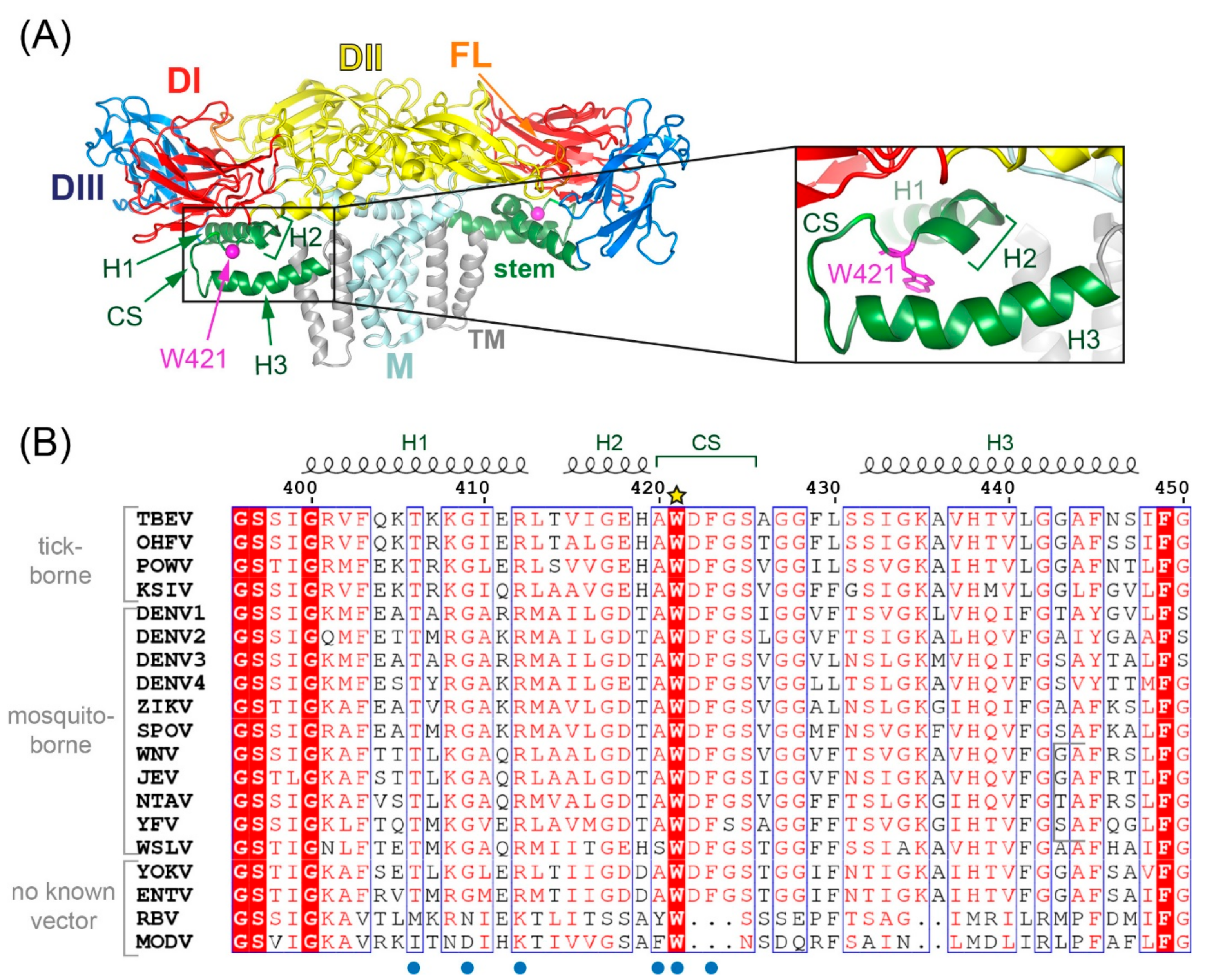
1. Introduction
2. Materials and Methods
2.1. Mutagenesis of cDNA Clones
2.2. Mutagenesis of Recombinant Subviral Particles (SVPs)
2.3. Production of SVPs
2.4. In Vitro Transcription of RNA
2.5. Production of Virus Particles
2.6. Immunofluorescence (IF) Staining
2.7. Serial Passaging Experiments
2.8. Sequence Analysis
2.9. Preparation of Cell Lysates
2.10. ELISA for the Quantification of E Protein
2.11. qPCR for the Quantification of RNA Copies
2.12. Focus Assay
2.13. ELISA for Determining the Maturation State of Viral Particles
2.14. ELISA with Conformation-Sensitive Mabs
2.15. Ultracentrifugation (Pelleting) of Viral Particles
2.16. Sedimentation Analyses
- (a)
- Separation of SVPs and virions: WT and mutant pellets were subjected to rate zonal centrifugation using 5 to 30% sucrose gradients in TAN buffer pH 8.0. After centrifugation for 70 min at 38,000 rpm and 4 °C (SW40 rotor, Beckman), the gradients were fractionated and the amount of E protein per fraction was determined by a quantitative four-layer ELISA as described above.
- (b)
- Separation of E dimers from E trimers: SVPs were incubated at low pH as described previously [25,41,42]. After back neutralization and solubilization with Triton X-100, sedimentation analysis was performed using 7% to 20% sucrose gradients in TAN buffer pH 8.0 containing 0.1% Triton X-100. After centrifugation for 20 h at 38,000 rpm and 15 °C (SW40 rotor, Beckman), the gradients were fractionated and the amount of E protein per fraction was determined by a quantitative four-layer ELISA as described above.
2.17. Statistics
2.18. GenBank Accession Numbers
3. Results
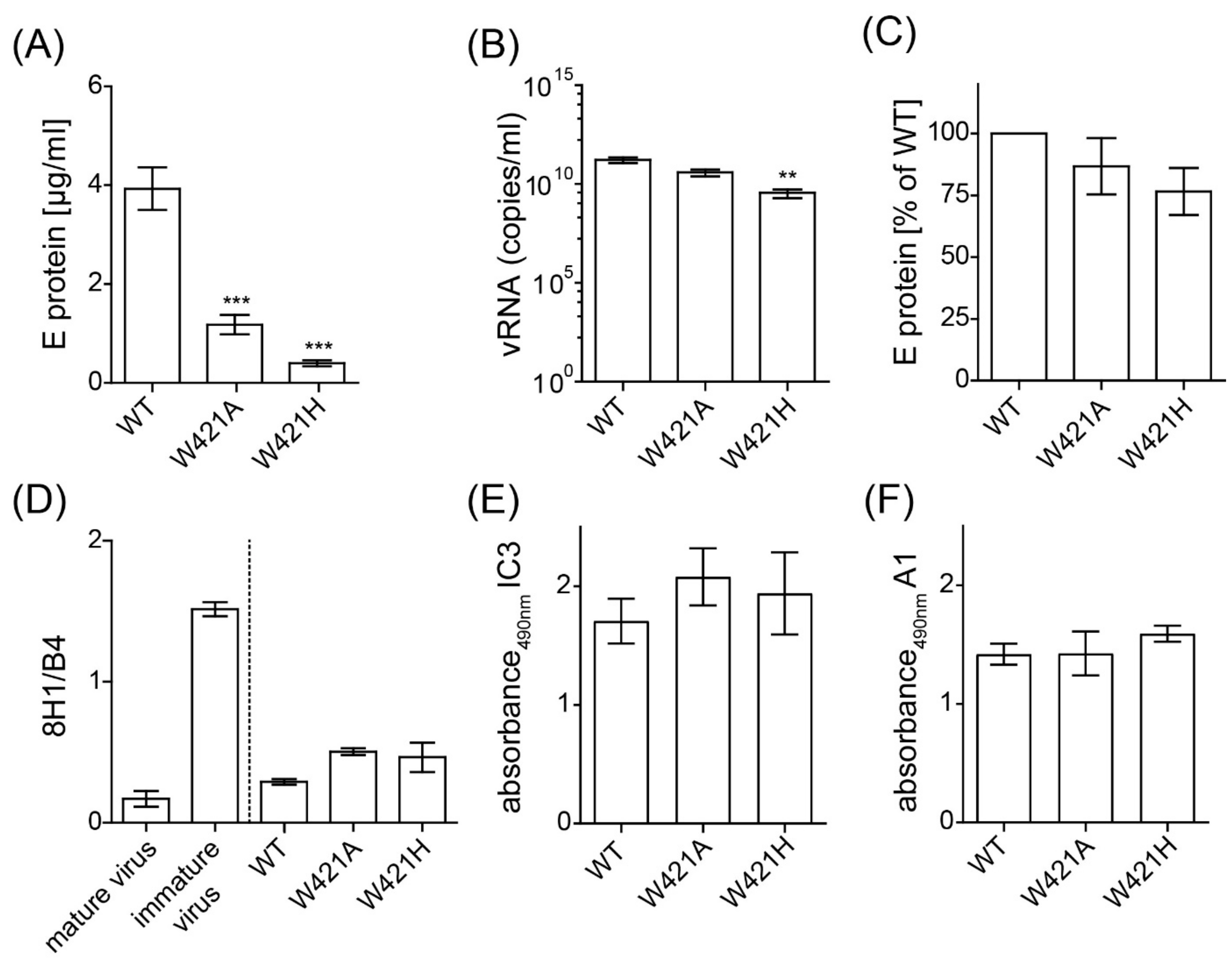
3.1. Secretion and Formation of Mutant Viral Particles
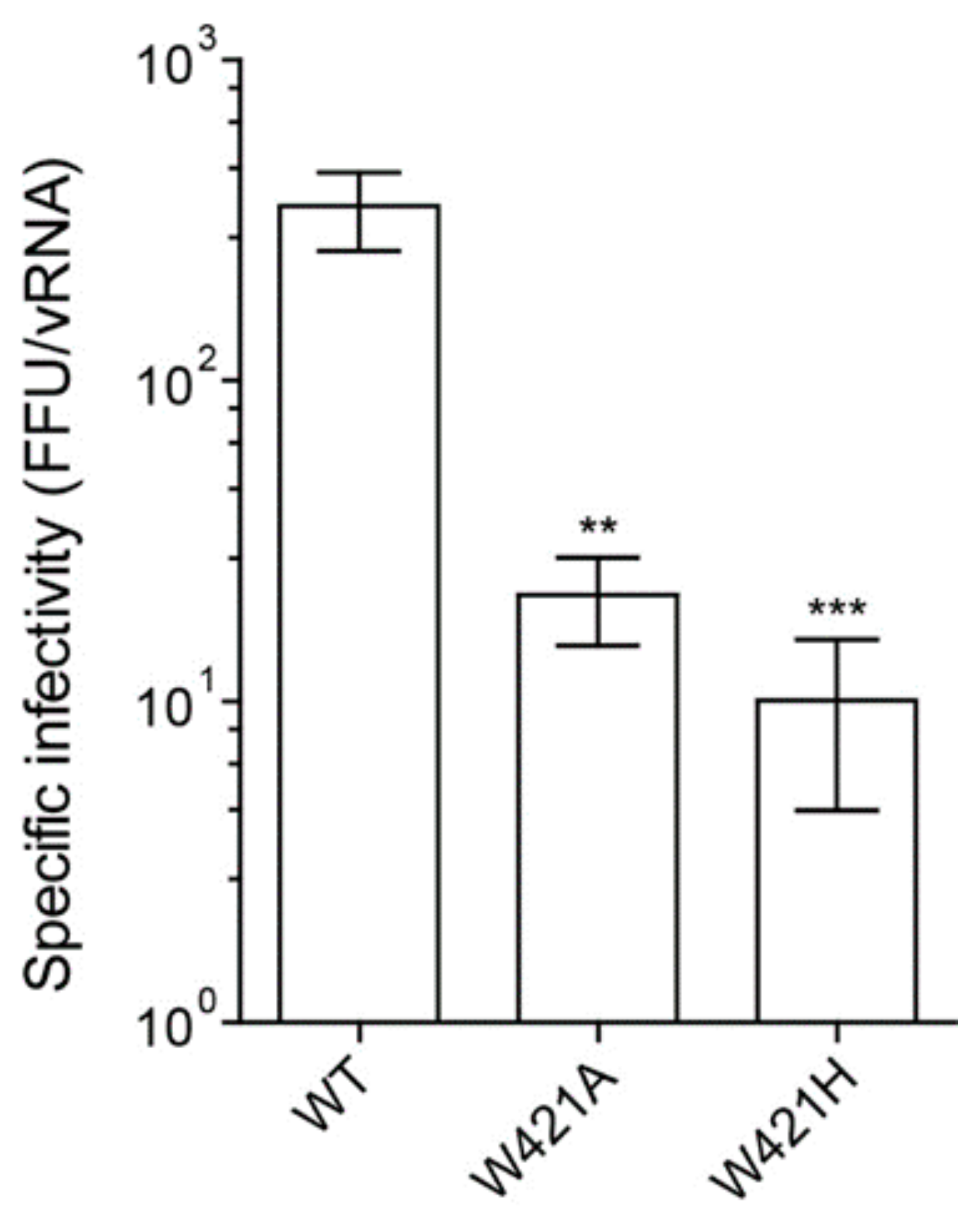
3.2. Effect of Mutations on Virion Infectivity and Stability
3.3. Effect of Mutations on Fusion-Related Processes
3.4. Rescue of Infectivity by Passaging Experiments
3.5. Effect of Mutations on Thermostability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef]
- Rey, F.A.; Stiasny, K.; Vaney, M.C.; Dellarole, M.; Heinz, F.X. The bright and the dark side of human antibody responses to flaviviruses: Lessons for vaccine design. EMBO Rep. 2018, 19, 206–224. [Google Scholar] [CrossRef]
- Zhang, X.; Ge, P.; Yu, X.; Brannan, J.M.; Bi, G.; Zhang, Q.; Schein, S.; Zhou, Z.H. Cryo-EM structure of the mature dengue virus at 3.5-A resolution. Nat. Struct. Mol. Biol. 2013, 20, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Kostyuchenko, V.A.; Zhang, Q.; Tan, J.L.; Ng, T.S.; Lok, S.M. Immature and mature dengue serotype 1 virus structures provide insight into the maturation process. J. Virol. 2013, 87, 7700–7707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostyuchenko, V.A.; Lim, E.X.; Zhang, S.; Fibriansah, G.; Ng, T.S.; Ooi, J.S.; Shi, J.; Lok, S.M. Structure of the thermally stable Zika virus. Nature 2016, 533, 425–428. [Google Scholar] [CrossRef] [Green Version]
- Sirohi, D.; Chen, Z.; Sun, L.; Klose, T.; Pierson, T.C.; Rossmann, M.G.; Kuhn, R.J. The 3.8 A resolution cryo-EM structure of Zika virus. Science 2016, 352, 467–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevvana, M.; Long, F.; Miller, A.S.; Klose, T.; Buda, G.; Sun, L.; Kuhn, R.J.; Rossmann, M.G. Refinement and analysis of the mature Zika virus cryo-EM structure at 3.1 Å resolution. Structure 2018, 26, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, S.H.; Zhu, L.; Nian, Q.G.; Yuan, S.; Gao, Q.; Hu, Z.; Ye, Q.; Li, X.F.; Xie, D.Y.; et al. Near-atomic structure of Japanese encephalitis virus reveals critical determinants of virulence and stability. Nat. Commun. 2017, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Fuzik, T.; Formanova, P.; Ruzek, D.; Yoshii, K.; Niedrig, M.; Plevka, P. Structure of tick-borne encephalitis virus and its neutralization by a monoclonal antibody. Nat. Commun. 2018, 9, 436. [Google Scholar] [CrossRef] [Green Version]
- DiNunno, N.M.; Goetschius, D.J.; Narayanan, A.; Majowicz, S.A.; Moustafa, I.; Bator, C.M.; Hafenstein, S.L.; Jose, J. Identification of a pocket factor that is critical to Zika virus assembly. Nat. Commun. 2020, 11, 4953. [Google Scholar] [CrossRef]
- Renner, M.; Dejnirattisai, W.; Carrique, L.; Martin, I.S.; Karia, D.; Ilca, S.L.; Ho, S.F.; Kotecha, A.; Keown, J.R.; Mongkolsapaya, J.; et al. Flavivirus maturation leads to the formation of an occupied lipid pocket in the surface glycoproteins. Nat. Commun. 2021, 12, 1238. [Google Scholar] [CrossRef]
- Hardy, J.M.; Newton, N.D.; Modhiran, N.; Scott, C.A.P.; Venugopal, H.; Vet, L.J.; Young, P.R.; Hall, R.A.; Hobson-Peters, J.; Coulibaly, F.; et al. A unified route for flavivirus structures uncovers essential pocket factors conserved across pathogenic viruses. Nat. Commun. 2021, 12, 3266. [Google Scholar] [CrossRef]
- Medits, I.; Vaney, M.C.; Rouvinski, A.; Rey, M.; Chamot-Rooke, J.; Rey, F.A.; Heinz, F.X.; Stiasny, K. Extensive flavivirus E trimer breathing accompanies stem zippering of the post-fusion hairpin. EMBO Rep. 2020, 21, e50069. [Google Scholar] [CrossRef]
- Harrison, S.C. Viral membrane fusion. Virology 2015, 479–480, 498–507. [Google Scholar]
- Rey, F.A.; Stiasny, K.; Heinz, F.X. Flavivirus structural heterogeneity: Implications for cell entry. Curr. Opin. Virol. 2017, 24, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, C.M.R.; Sevvana, M.; Kuhn, R.J. Structure-guided paradigm shifts in flavivirus assembly and maturation mechanisms. Adv. Virus Res. 2020, 108, 33–83. [Google Scholar] [PubMed]
- Yu, I.M.; Zhang, W.; Holdaway, H.A.; Li, L.; Kostyuchenko, V.A.; Chipman, P.R.; Kuhn, R.J.; Rossmann, M.G.; Chen, J. Structure of the immature dengue virus at low pH primes proteolytic maturation. Science 2008, 319, 1834–1837. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Firth, A.E. A Review of Flaviviruses that Have No Known Arthropod Vector. Viruses 2017, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Mandl, C.W.; Ecker, M.; Holzmann, H.; Kunz, C.; Heinz, F.X. Infectious cDNA clones of tick-borne encephalitis virus European subtype prototypic strain Neudoerfl and high virulence strain Hypr. J. Gen. Virol. 1997, 78 Pt 5, 1049–1057. [Google Scholar] [CrossRef]
- Bond, S.R.; Naus, C.C. RF-Cloning.org: An online tool for the design of restriction-free cloning projects. Nucleic Acids Res. 2012, 40, W209–W213. [Google Scholar] [CrossRef] [Green Version]
- Sanger, F.; Coulson, A.R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol. 1975, 94, 441–448. [Google Scholar] [CrossRef]
- Allison, S.L.; Mandl, C.W.; Kunz, C.; Heinz, F.X. Expression of cloned envelope protein genes from the flavivirus tick-borne encephalitis virus in mammalian cells and random mutagenesis by PCR. Virus Genes 1994, 8, 187–198. [Google Scholar] [CrossRef]
- Schalich, J.; Allison, S.L.; Stiasny, K.; Mandl, C.W.; Kunz, C.; Heinz, F.X. Recombinant subviral particles from tick-borne encephalitis virus are fusogenic and provide a model system for studying flavivirus envelope glycoprotein functions. J. Virol. 1996, 70, 4549–4557. [Google Scholar] [CrossRef] [Green Version]
- Pangerl, K.; Heinz, F.X.; Stiasny, K. Mutational analysis of the zippering reaction during flavivirus membrane fusion. J. Virol. 2011, 85, 8495–8501. [Google Scholar] [CrossRef] [Green Version]
- Blazevic, J.; Rouha, H.; Bradt, V.; Heinz, F.X.; Stiasny, K. Membrane anchors of the structural flavivirus proteins and their role in virus assembly. J. Virol. 2016, 90, 6365–6378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritz, R.; Blazevic, J.; Taucher, C.; Pangerl, K.; Heinz, F.X.; Stiasny, K. The unique transmembrane hairpin of flavivirus fusion protein E is essential for membrane fusion. J. Virol. 2011, 85, 4377–4385. [Google Scholar] [CrossRef] [Green Version]
- Kofler, R.M.; Hoenninger, V.M.; Thurner, C.; Mandl, C.W. Functional analysis of the tick-borne encephalitis virus cyclization elements indicates major differences between mosquito-borne and tick-borne flaviviruses. J. Virol. 2006, 80, 4099–4113. [Google Scholar] [CrossRef] [Green Version]
- Iacono-Connors, L.C.; Smith, J.F.; Ksiazek, T.G.; Kelley, C.L.; Schmaljohn, C.S. Characterization of Langat virus antigenic determinants defined by monoclonal antibodies to E, NS1 and preM and identification of a protective, non-neutralizing preM-specific monoclonal antibody. Virus Res. 1996, 43, 125–136. [Google Scholar] [CrossRef]
- Orlinger, K.K.; Kofler, R.M.; Heinz, F.X.; Hoenninger, V.M.; Mandl, C.W. Selection and analysis of mutations in an encephalomyocarditis virus internal ribosome entry site that improve the efficiency of a bicistronic flavivirus construct. J. Virol. 2007, 81, 12619–12629. [Google Scholar] [CrossRef] [Green Version]
- Heinz, F.X.; Tuma, W.; Guirakhoo, F.; Kunz, C. A model study of the use of monoclonal antibodies in capture enzyme immunoassays for antigen quantification exploiting the epitope map of tick-borne encephalitis virus. J. Biol. Stand. 1986, 14, 133–141. [Google Scholar] [CrossRef]
- Heinz, F.X.; Stiasny, K.; Puschner-Auer, G.; Holzmann, H.; Allison, S.L.; Mandl, C.W.; Kunz, C. Structural changes and functional control of the tick-borne encephalitis virus glycoprotein E by the heterodimeric association with protein prM. Virology 1994, 198, 109–117. [Google Scholar] [CrossRef]
- Heinz, F.X.; Kunz, C. Homogeneity of the structural glycoprotein from European isolates of tick-borne encephalitis virus: Comparison with other flaviviruses. J. Gen. Virol. 1981, 57 Pt 2, 263–274. [Google Scholar] [CrossRef]
- Taucher, C.; Berger, A.; Mandl, C.W. A trans-complementing recombination trap demonstrates a low propensity of flaviviruses for intermolecular recombination. J. Virol. 2010, 84, 599–611. [Google Scholar] [CrossRef] [Green Version]
- Haslwanter, D.; Blaas, D.; Heinz, F.X.; Stiasny, K. A novel mechanism of antibody-mediated enhancement of flavivirus infection. PLoS Pathog. 2017, 13, e1006643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guirakhoo, F.; Heinz, F.X.; Kunz, C. Epitope model of tick-borne encephalitis virus envelope glycoprotein E: Analysis of structural properties, role of carbohydrate side chain, and conformational changes occurring at acidic pH. Virology 1989, 169, 90–99. [Google Scholar] [CrossRef]
- Mandl, C.W.; Guirakhoo, F.; Holzmann, H.; Heinz, F.X.; Kunz, C. Antigenic structure of the flavivirus envelope protein E at the molecular level, using tick-borne encephalitis virus as a model. J. Virol. 1989, 63, 564–571. [Google Scholar] [CrossRef] [Green Version]
- Stiasny, K.; Kiermayr, S.; Holzmann, H.; Heinz, F.X. Cryptic properties of a cluster of dominant flavivirus cross-reactive antigenic sites. J. Virol. 2006, 80, 9557–9568. [Google Scholar] [CrossRef] [Green Version]
- Stiasny, K.; Brandler, S.; Kossl, C.; Heinz, F.X. Probing the flavivirus membrane fusion mechanism by using monoclonal antibodies. J. Virol. 2007, 81, 11526–11531. [Google Scholar] [CrossRef] [Green Version]
- Allison, S.L.; Stiasny, K.; Stadler, K.; Mandl, C.W.; Heinz, F.X. Mapping of functional elements in the stem-anchor region of tick-borne encephalitis virus envelope protein E. J. Virol. 1999, 73, 5605–5612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, S.L.; Schalich, J.; Stiasny, K.; Mandl, C.W.; Heinz, F.X. Mutational evidence for an internal fusion peptide in flavivirus envelope protein E. J. Virol. 2001, 75, 4268–4275. [Google Scholar] [CrossRef] [Green Version]
- Fritz, R.; Stiasny, K.; Heinz, F.X. Identification of specific histidines as pH sensors in flavivirus membrane fusion. J. Cell Biol. 2008, 183, 353–361. [Google Scholar] [CrossRef]
- Lindenbach, B.D.; Murray, C.L.; Thiel, H.J.; Rice, C.M. Flaviviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Racaniello, V.R., Roizman, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 712–746. [Google Scholar]
- Newton, N.D.; Hardy, J.M.; Modhiran, N.; Hugo, L.E.; Amarilla, A.A.; Bibby, S.; Venugopal, H.; Harrison, J.J.; Traves, R.J.; Hall, R.A.; et al. The structure of an infectious immature flavivirus redefines viral architecture and maturation. Sci. Adv. 2021, 7, eabe4507. [Google Scholar] [CrossRef]
- Chen, L.; Wang, M.; Zhu, D.; Sun, Z.; Ma, J.; Wang, J.; Kong, L.; Wang, S.; Liu, Z.; Wei, L.; et al. Implication for alphavirus host-cell entry and assembly indicated by a 3.5Å resolution cryo-EM structure. Nat. Commun. 2018, 9, 5326. [Google Scholar] [CrossRef] [Green Version]
- Liao, M.; Kielian, M. Functions of the stem region of the Semliki Forest virus fusion protein during virus fusion and assembly. J. Virol. 2006, 80, 11362–11369. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.R.; Zou, G.; Hsieh, S.C.; Qing, M.; Tsai, W.Y.; Shi, P.Y.; Wang, W.K. The helical domains of the stem region of dengue virus envelope protein are involved in both virus assembly and entry. J. Virol. 2011, 85, 5159–5171. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′to 3′) |
|---|---|
| rf 1 stem fwd 2 | GAG TCA TCA ATG GTT CCA AAA AGG GAG CAG CAT CGG AAG GGT TTT CC |
| rf stem rev 3 | GGC GCT TTC AAC AGC ATC TTC GGG GGA GTG GGG TTT CTA C |
| Mutation | Primer | Sequence (5′to 3′) |
|---|---|---|
| W421A | fwd 1 | GTG ATA GGA GAG CAC GCC GCA GAC TTC GGT TCT GCT GGA |
| W421A | rev 2 | TCC AGC AGA ACC GAA GTC TGC GGC GTG CTC TCC TAT CAC |
| W421H | fwd | ATA GGA GAG CAC GCC CAT GAC TTC GGT TCT GCT |
| W421H | rev | AGC AGA ACC GAA GTC ATG GGC GTG CTC TCC TAT |
| W421L | fwd | ATA GGA GAG CAC GCC CTT GAC TTC GGT TCT GCT |
| W421L | rev | AGC AGA ACC GAA GTC AAG GGC GTG CTC TCC TAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medits, I.; Heinz, F.X.; Stiasny, K. An Absolutely Conserved Tryptophan in the Stem of the Envelope Protein E of Flaviviruses Is Essential for the Formation of Stable Particles. Viruses 2021, 13, 1727. https://doi.org/10.3390/v13091727
Medits I, Heinz FX, Stiasny K. An Absolutely Conserved Tryptophan in the Stem of the Envelope Protein E of Flaviviruses Is Essential for the Formation of Stable Particles. Viruses. 2021; 13(9):1727. https://doi.org/10.3390/v13091727
Chicago/Turabian StyleMedits, Iris, Franz X. Heinz, and Karin Stiasny. 2021. "An Absolutely Conserved Tryptophan in the Stem of the Envelope Protein E of Flaviviruses Is Essential for the Formation of Stable Particles" Viruses 13, no. 9: 1727. https://doi.org/10.3390/v13091727
APA StyleMedits, I., Heinz, F. X., & Stiasny, K. (2021). An Absolutely Conserved Tryptophan in the Stem of the Envelope Protein E of Flaviviruses Is Essential for the Formation of Stable Particles. Viruses, 13(9), 1727. https://doi.org/10.3390/v13091727