
Genotype Diversity and Spread of White Spot Syndrome Virus (WSSV) in Madagascar (2012–2016)

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
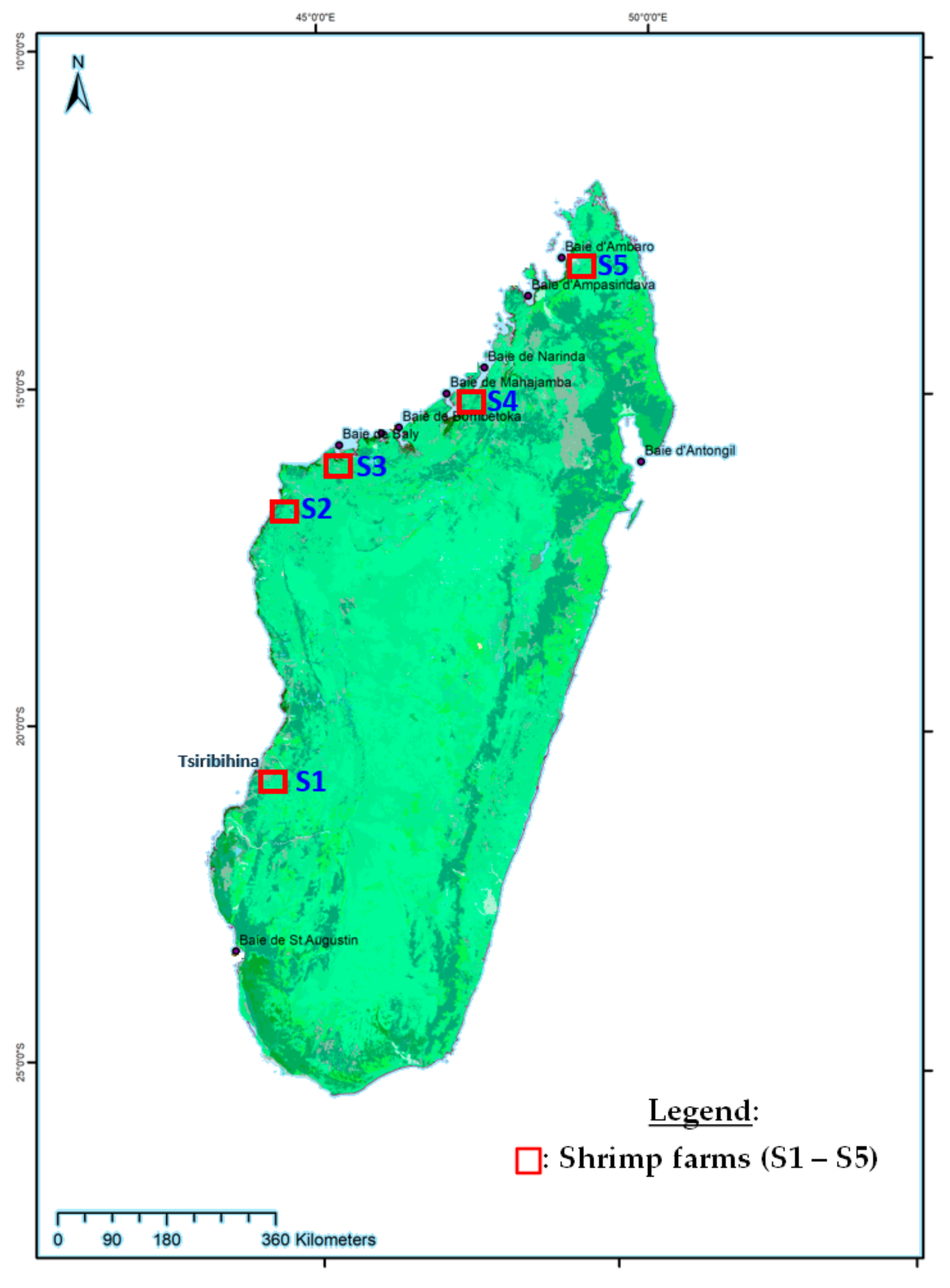
2.1. Specimen Collections
2.2. Amplification and Detection of WSSV
2.3. WSSV Genotyping
2.4. DNA Sequencing Analysis
3. Results
3.1. WSSV Screening and Genotyping
3.2. VNTR Analysis of ORF125
3.3. VNTR Analysis of ORF94
3.4. VNTR Analysis in ORF75
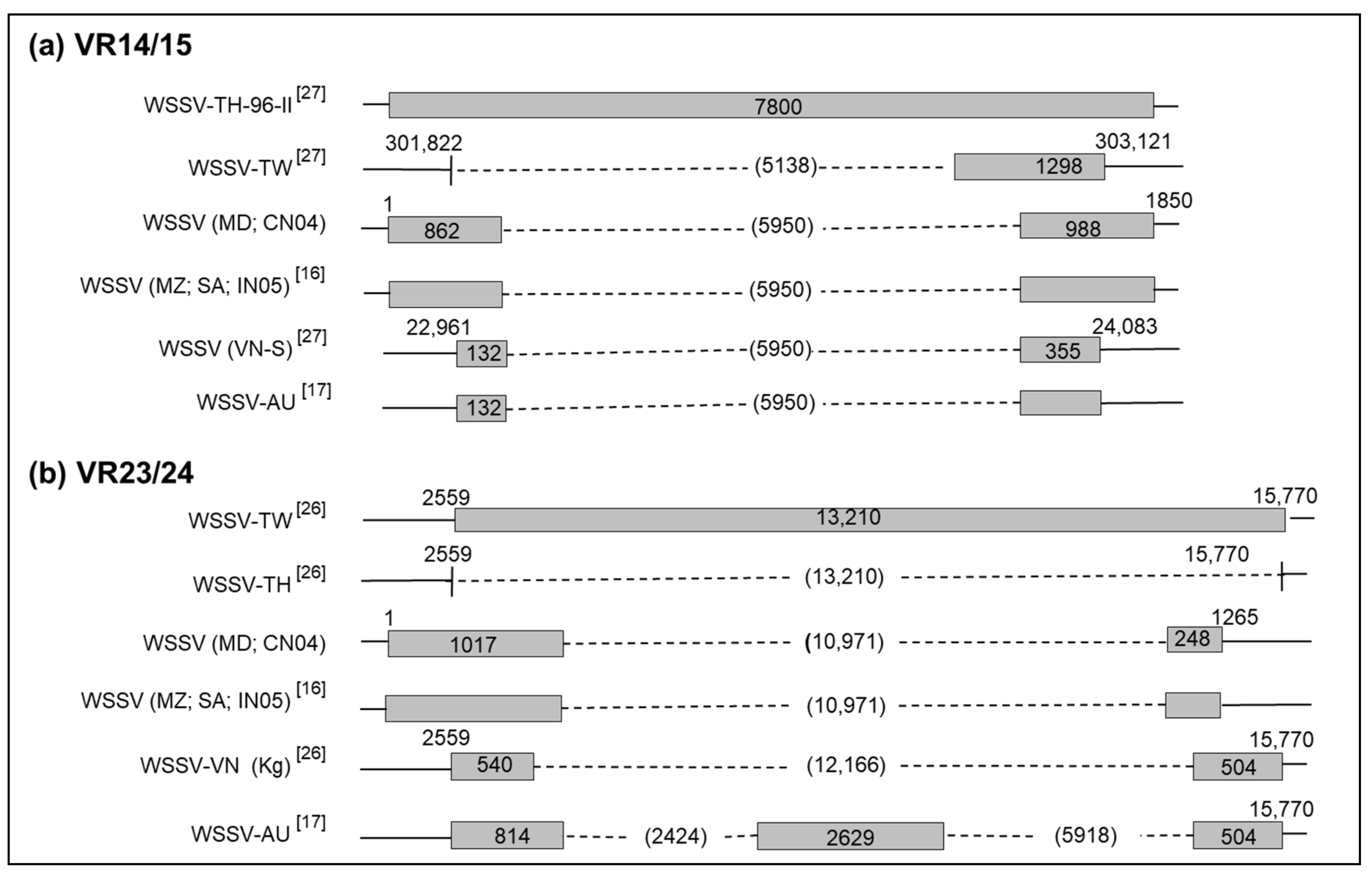
3.5. Deletion in VR14/15
3.6. Deletion in VR23/24
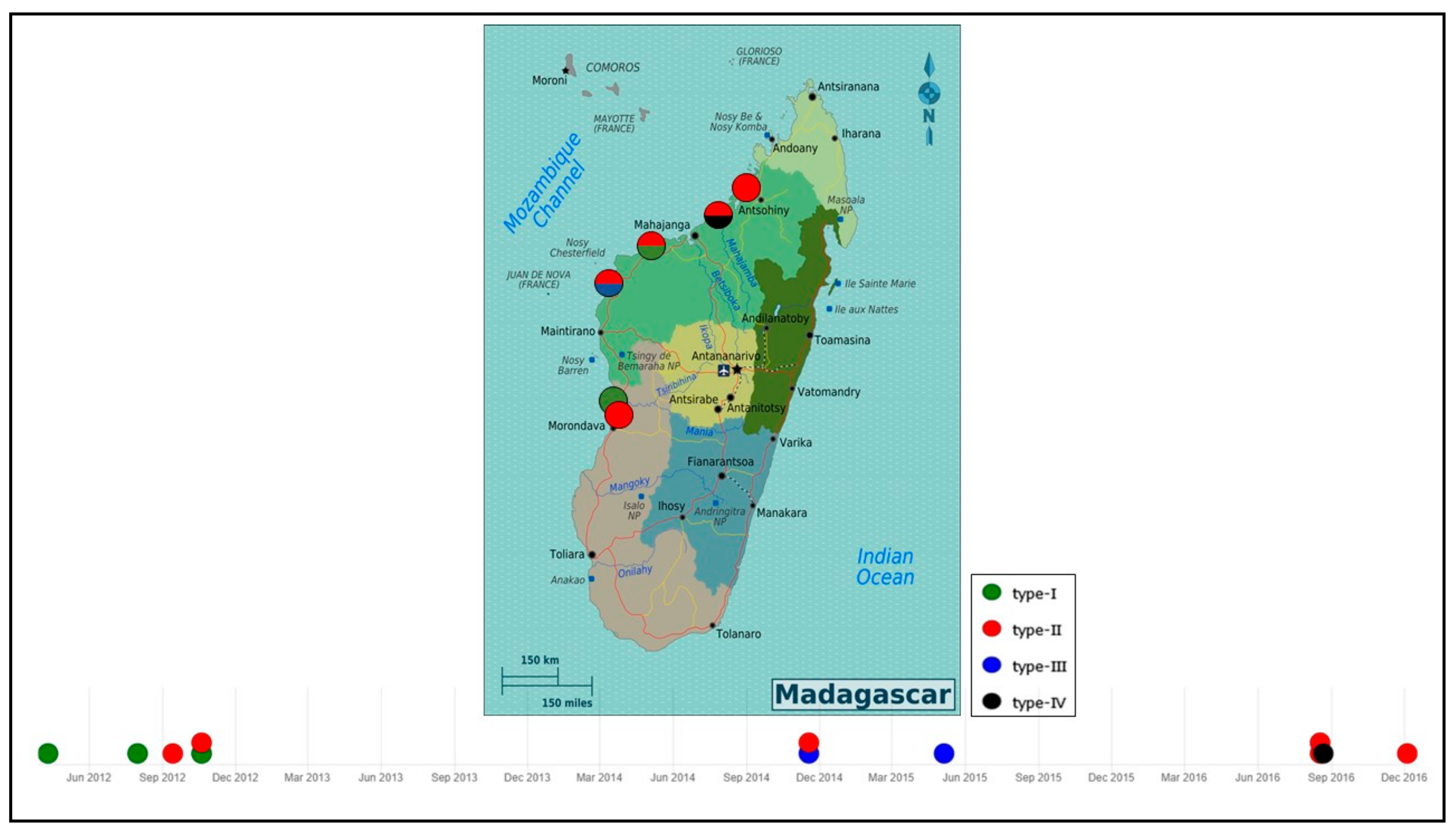
3.7. WSSV Genotype and Its Circulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mayo, M.A. Virus taxonomy—Houston 2002. Arch. Virol. 2002, 147, 1071–1076. [Google Scholar] [PubMed]
- Lillehammer, M.; Bangera, R.; Salazar, M.; Vela, S.; Erazo, E.C.; Suarez, A.; Cock, J.; Rye, M.; Robinson, N.A. Genomic selection for white spot syndrome virus resistance in whiteleg shrimp boosts survival under an experimental challenge test. Sci. Rep. 2020, 10, 20571. [Google Scholar] [CrossRef] [PubMed]
- Flegel, T.W. Major viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J. Microbiol. Biotechnol. 1997, 13, 433–442. [Google Scholar] [CrossRef]
- OIE. Chapter 2.2.8. Infection with White Spot Syndrome Virus. In Manual of Diagnostic Tests for Aquatic Animals; World Organisation for Animal Health: Paris, France, 2021. [Google Scholar]
- Lo, C.-F.; Ho, C.; Peng, S.; Chen, C.; Hsu, H.; Chiu, Y.; Chang, C.; Liu, K.; Su, M.; Wang, C.; et al. White Spot Syndrome Baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods. Dis. Aquat. Org. 1996, 27, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, A.; Rajkumar, M.; Sun, J.; Wang, M.; Senthil Kumar, K. Mud crab, Scylla tranquebarica (Decapoda: Portunidae), a new host for the white spot syndrome virus. Aquac. Res. 2011, 42, 308–312. [Google Scholar] [CrossRef]
- Gunasekaran, T.; Gopalakrishnan, A.; Deivasigamani, B.; Seralathan, M.V.; Kathirkaman, P. Spontaneous white spot syndrome virus (WSSV) infection in mud crab (Scylla serrata Forskal 1775) fattening pens farm of south east coast of India. Comp. Clin. Path. 2018, 27, 413–419. [Google Scholar] [CrossRef]
- Ross, E.P.; Behringer, D.C.; Bojko, J. White spot syndrome virus and the Caribbean spiny lobster, Panulirus argus: Susceptibility and behavioral immunity. J. Invertebr. Pathol. 2019, 162, 1–9. [Google Scholar] [CrossRef]
- Diggles, B.K.; Chong, R.; Hawkins, K.; Underwood, D.; Genge, B.; Driver, L.; Kistler, C. Investigation into white spots in the carapace of a moribund mud crab (Scylla serrata) from a white spot syndrome virus (WSSV) positive zone in Moreton Bay, Australia. Aust. Vet. J. 2020, 98, 550–554. [Google Scholar] [CrossRef]
- Durand, S.; Lightner, D.V.; Redman, R.M.; Bonami, J.R. Ultrastructure and morphogenesis of White Spot Syndrome Baculovirus (WSSV). Dis. Aquat. Org. 1997, 29, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Paz, A. White spot syndrome virus: An overview on an emergent concern. Vet. Res. 2010, 41, 43. [Google Scholar] [CrossRef] [Green Version]
- Chou, H.; Huang, C.; Wang, C.; Chiang, H.; Lo, C. Pathogenicity of a baculovirus infection causing white spot syndrome in cultured penaeid shrimp in Taiwan. Dis. Aquat. Org. 1995, 23, 165–173. [Google Scholar] [CrossRef]
- Lo, C.-F.; Leu, J.; Ho, C.; Chen, C.; Peng, S.; Chen, Y.-T.; Chou, C.; Yeh, P.; Huang, C.; Chou, H.; et al. Detection of baculovirus associated with white spot syndrome (WSBV) in penaeid shrimps using polymerase chain reaction. Dis. Aquat. Org. 1996, 25, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Lightner, D.V.; Redman, R.M.; Poulos, B.T.; Nunan, L.M.; Mari, J.L.; Hasson, K.W. Risk of spread of penaeid shrimp viruses in the Americas by the international movement of live and frozen shrimp. Rev. Sci. Tech. 1997, 16, 146–160. [Google Scholar] [CrossRef]
- Tang, K.F.; Navarro, S.A.; Pantoja, C.R.; Aranguren, F.L.; Lightner, D.V. New genotypes of white spot syndrome virus (WSSV) and Taura syndrome virus (TSV) from the Kingdom of Saudi Arabia. Dis. Aquat. Org. 2012, 99, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.F.; Le Groumellec, M.; Lightner, D.V. Novel, closely related, white spot syndrome virus (WSSV) genotypes from Madagascar, Mozambique and the Kingdom of Saudi Arabia. Dis. Aquat. Org. 2013, 106, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Oakey, H.J.; Smith, C.S. Complete genome sequence of a white spot syndrome virus associated with a disease incursion in Australia. Aquaculture 2018, 484, 152–159. [Google Scholar] [CrossRef]
- Knibb, W.; Le, C.; Katouli, M.; Bar, I.; Lloyd, C. Assessment of the origin of white spot syndrome virus DNA sequences in farmed Penaeus monodon in Australia. Aquaculture 2018, 494, 26–29. [Google Scholar] [CrossRef]
- Lightner, D.V. A Handbook of Shrimp Pathology and Diagnostic Procedures for Diseases of Cultured Penaeid Shrimp; World Aquaculture Society: Baton Rouge, LA, USA, 1996. [Google Scholar]
- Lightner, D.; Redman, R.; Pantoja, C.; Tang, K.; Noble, B.; Schofield, P.; Mohney, L.; Nunan, L.; Navarro, S. Historic emergence, impact and current status of shrimp pathogens in the Americas. J. Invertebr. Pathol. 2012, 110, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Stentiford, G.; Neil, D.; Peeler, E.; Shields, J.; Small, H.; Flegel, T.; Vlak, J.; Jones, B.; Morado, F.; Moss, S.; et al. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J. Invertebr. Pathol. 2012, 110, 141–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, P.K.; Geetha, R.; Ravisankar, T.; Avunje, S.; Solanki, H.G.; Abraham, T.J.; Vinoth, S.P.; Jithendran, K.P.; Alavandi, S.V.; Vijayan, K.K. Economic loss due to diseases in Indian shrimp farming with special reference to Enterocytozoon hepatopenaei (EHP) and white spot syndrome virus (WSSV). Aquaculture 2021, 533, 736231. [Google Scholar] [CrossRef]
- Gillett, R. Global Study of Shrimp Fisheries; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008; Volume 475, p. 331. [Google Scholar]
- Goedefroit, S.; Chaboud, C.; Breton, Y. La ruée vers l’or rose: Regards croisés sur la pêche crevettière traditionnelle à Madagascar; IRD Editions: Paris, France, 2002. [Google Scholar]
- Chamberlain, G.; Lightner, D.; Towner, R.; van Wyk, P.; Villarreal, M.; Akazawa, N.; Alvial, A. Case Study of the Outbreak of White Spot Syndrome Virus at Shrimp Farms in Mozambique and Madagascar: Impacts and Management Recommendations; Responsible Aquaculture Foundation: Portsmouth, NH, USA, 2013. [Google Scholar]
- Dieu, B.T.M.; Marks, H.; Siebenga, J.J.; Goldbach, R.W.; Zuidema, D.; Duong, T.P.; Vlak, J.M. Molecular epidemiology of white spot syndrome virus within Vietnam. J. Gen. Virol. 2004, 85, 3607–3618. [Google Scholar] [CrossRef] [PubMed]
- Dieu, B.T.M.; Marks, H.; Zwart, M.P.; Vlak, J.M. Evaluation of white spot syndrome virus variable DNA loci as molecular markers of virus spread at intermediate spatiotemporal scales. J. Gen. Virol. 2010, 91, 1164–1172. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Marks, H.; Van Duijse, J.J.A.; Zuidema, D.; Van Hulten, M.C.W.; Vlak, J.M. Fitness and virulence of an ancestral White Spot Syndrome Virus isolate from shrimp. Virus Res. 2005, 110, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-F.; Yu, H.-T.; Tzeng, H.-F.; Leu, J.-H.; Chou, C.-M.; Huang, C.-J.; Wang, C.-H.; Lin, J.-Y.; Kou, G.-H.; Lo, C.-F. Identification and characterization of a shrimp white spot syndrome virus (WSSV) gene that encodes a novel chimeric polypeptide of cellular-type thymidine kinase and thymidylate kinase. Virology 2000, 277, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Dieu, B.T.M.; Zwart, M.P.; Vlak, J.M. Can VNTRs be used to study genetic variation within white spot syndrome virus isolates? J. Fish Dis. 2010, 33, 689–693. [Google Scholar] [CrossRef]
- Wongteerasupaya, C.; Pungchai, P.; Withyachumnarnkul, B.; Boonsaeng, V.; Panyim, S.; Flegel, T.; Walker, P. High variation in repetitive DNA fragment length for White Spot Syndrome Virus (WSSV) isolates in Thailand. Dis. Aquat. Org. 2003, 54, 253–257. [Google Scholar] [CrossRef]
- Piamsomboon, P.; Chaweepack, T.; Han, J.E.; Tang, K.F.; Wongtavatchai, J. Genetic variation of white spot syndrome virus (WSSV) in Pacific white shrimp Litopenaeus vannamei (Boone 1931) culture of Thailand. Aquac. Res. 2018, 49, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Xiao, J.; Liu, L.; Pan, Y.; Yan, S.; Wang, Y. Characterization and prevalence of a novel white spot syndrome viral genotype in naturally infected wild crayfish, Procambarus clarkii, in Shanghai, China. Virus Dis. 2017, 28, 250–261. [Google Scholar] [CrossRef]
- Argimón, S.; AbuDahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and sharing data for genomic epidemiology and phylogeography. Microb. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- République Fédérale Islamique des Comores. Atlas des Ressources Côtières de l’Afrique Orientale; Programme des Nations Unies pour l’Environnement: Nairobi, Kenya, 2002. [Google Scholar]
- Sangamaheswaran, A.P.; Jeyaseelan, M.J.P. White Spot Viral Disease in Penaeid Shrimp—A Review. Naga, ICLARM Q. 2001, 24, 16–22. [Google Scholar]
- PriyanghaS, J.; Gopalakrishnan, A.; MuhilVannan, S.; Gunasekaran, T.; Somasundaram, S.T. First report of pedunculate barnacle (Octolasmis neptuni), as potential asymptomatic carrier of white spot syndrome virus (WSSV). Comp. Haematol. Int. 2020, 29, 631–638. [Google Scholar]
- Mohan, C.V.; Phillips, M.J.; Bhat, B.V.; Umesh, N.R.; Padiyar, P.A. Farm-level plans and husbandry measures for aquatic animal disease emergencies. Rev. Sci. Tech. 2008, 27, 161–173. [Google Scholar] [CrossRef]
- Oakey, J.; Smith, C.; Underwood, D.; Afsharnasab, M.; Alday-Sanz, V.; Dhar, A.; Sivakumar, S.; Hameed, A.S.S.; Beattie, K.; Crook, A. Global distribution of white spot syndrome virus genotypes determined using a novel genotyping assay. Arch. Virol. 2019, 164, 2061–2082. [Google Scholar] [CrossRef] [Green Version]
- Waikhom, G.; John, K.R.; George, M.R.; Jeyaseelan, M.J.P. Differential host passaging alters pathogenicity and induces genomic variation in white spot syndrome virus. Aquaculture 2006, 261, 54–63. [Google Scholar] [CrossRef]
- Hoa, T.T.T.; Zwart, M.P.; Phuong, N.T.; De Jong, M.C.M.; Vlak, J.M. Low numbers of repeat units in variable number of tandem repeats (VNTR) regions of white spot syndrome virus are correlated with disease outbreaks. J. Fish Dis. 2012, 35, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Gao, M.; Xu, L.; Yang, F. Comparative genomic analysis of three white spot syndrome virus isolates of different virulence. Virus Genes 2017, 53, 249–258. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primers | Forward/Reverse Primers | Sequence (5’–3’) | Cycling Conditions | PCR Product Size (bp) | References | |
|---|---|---|---|---|---|---|
| WSSV screening | 146-1 (first step) | 146-F1 | ACTACTAACTTCAGCCTATCTAG | 94 °C 4 min, 55 °C 1 min, 72 °C 2 min; 40 × [94 °C 4 min, 55 °C 1 min, 72 °C 4 min]; 72 °C 5 min | 1447 | [13] |
| 146-R1 | TAATGCGGGTGTAATGTTCTTACGA | |||||
| 146-2 (second step) | 146-F2 | GTAACTGCCCCTTCCATCTCCA | 40 × [94 °C 1 min, 62 °C 1 min, 72 °C 2 min]; 72 °C 5 min | 941 | ||
| 146-R2 | TACGGCAGCTGCTGCACCTTGT | |||||
| Variable loci First PCR | ORF75-flank | ORF75-flank-F | GAAGCAGTATCTCTAACAC | 94 °C 4 min; 40 × [94 °C 1 min, 49/50 °C 80 s, 72 °C 1 min]; 72 °C 5 min | 868 | [26] |
| ORF75-flank-R | CAACAGGTGCGTAAAAGAAG | |||||
| ORF94 | ORF94-F | TCTACTCGAGGAGGTGACGAC | 94 °C 3 min; 35 × [94 °C 30 s, 55 °C 30 s, 72 °C 1 min]; 72 °C 7 min | 506 to 1262 | [33] | |
| ORF94-R | AGCAGGTGTGTACACATTTCATG | |||||
| ORF125-flank | ORF125 flank-F | CGAAATCTTGATATGTTGTGC | 94 °C 3 min; 35 × [94 °C 30 s, 55 °C 30 s, 72 °C 1 min]; 72 °C 7 min | 652 | [26] | |
| ORF125 flank-R | CCATATCCATTGCCCTTCTC | |||||
| VR14/15-complete | VR14/15-complete-F | AATATGGAACGACGGGTG | 94 °C 3; 35 × [94 °C 30 s, 50 °C 30 s, 72 °C 2 min]; 72 °C 7 min | 1851 | [29] | |
| VR14/15-complete-R | GACCAGCGCCTCTTCAG | |||||
| VR23/24-south | VR23/24-south-F | GTAGTGCATGTTTCTCTAAC | 94 °C 3 min; 35 × [94 °C 30 s, 45 °C 30 s, 72 °C 2 min]; 72 °C 7 min | 1264 | [27] | |
| VR23/24-south-R | GTAAGTTTATTGCTGAGAAG | |||||
| Variable loci Second PCR | ORF73/ORF77 | ORF73-F | CTTTCACCGCTCTCACCAAC | 94 °C 3 min; 35 × [94 °C 30 s, 55 °C 30 s, 72 °C 2 min]; 72 °C 7 min | 1739 | [16] |
| ORF77-R | GGGTTCACCAGAGAGACAGG | |||||
| ORF93/ORF96 | ORF93-F1 | CGCCCTATTACCATTGATGC | 94 °C 4 min; 40 × [94 °C 1 min, 58 °C 60 min, 72 °C 1 min]; 72 °C 5 min | 348 | [15] | |
| ORF96-R1 | GCAACAAATTCCCCTTTCAA | |||||
| TJW14/15 | TJW14/15-F | TCAACAACCCAAATCCCATT | 94 °C 3 min; 40 × [94 °C 15 s, 60 °C 15 s, 72 °C 1 min]; 72 °C 5 min | 3343 | [34] | |
| TJW14/15-R | CTCTCAATCTTCCCCCAACA | |||||
| VR14/15-screen | VR14/15-screen-F | GAGATGCGAACCACTAAAAG | 94 °C 3 min; 40 × [94 °C 15 s, 60 °C 15 s, 72 °C 1 min]; 72 °C 5 min | 600 | [26] | |
| VR14/15-screen-R | ATGGAGGCGAGACTTGC | |||||
| VR23/24-screen | VR23/24-screen-F | CACACTTGAAAAATACACCAG | 94 °C 3 min; 40 × [94 °C 15 s, 49 °C 65 s, 72 °C 1 min]; 72 °C 5 min | 548 | ||
| VR23/24-screen-R | GTAAGTTTATTGCTGAGAAG | |||||
| Variable loci Third PCR | ORF75 | TJW75-F | TCTGAAGCTGGGGGAACTAA | 94 °C 3 min; 40 × [94 °C 15 s, 60 °C 15 s, 72 °C 1 min]; 72 °C 5 min | 702 | [34] |
| TJW75-R | GAGCAACTCTGCACAGCATC | |||||
| VR23/24-south04 | VR23/24-south04-F | CTACAACGGCCAAGTCAT | 94 °C 3 min; 40 × [94 °C 15 s, 60 °C 15 s, 72 °C 1 s]; 72 °C 5 min | 1500/2000 | [34] | |
| VR23/24-1 | VR23/24-1-R | ATGATTGTATTCGTCGAAGG | ||||
| ORF23/24 | ORF23/24-F | GGTAGGAGAAGGTACGCACG | 94 °C 3 min; 40 × [94 °C 30 s, 60 °C 15 s, 72 °C 1 min]; 72 °C 5 min | 4025 | [35] | |
| ORF23/24-R | GCCCAGATTGGTCATGTCCA |
| Site | Type | Host Species 1 | Sampling Date | Nb. of Samples | Nb. of Repeat Units | Deletion Size (bp) | |||
|---|---|---|---|---|---|---|---|---|---|
| ORF75 | ORF94 2 | ORF125 | VR14/15 | VR23/24 | |||||
| S1 | Farm | P. monodon | April 2012 | 4 | 3 | del | 7 | 5950 | 10,971 |
| S2 | P. monodon | September 2012 | 2 | 3 | del | 6 | 5950 | 10,971 | |
| P. monodon | December 2014 | 2 | 3 | del | 6 | 5950 | 10,971 | ||
| P. monodon | May 2015 | 1 | 3 | del | 5 | 5950 | 10,971 | ||
| F. indicus | November 2014 | 4 | 3 | del | 5 | 5950 | 10,971 | ||
| F. indicus | November 2014 | 2 | 3 | del | 6 | 5950 | 10,971 | ||
| S3 | P. monodon | October 2012 | 1 | 3 | del | 6 | 5950 | 10,971 | |
| F. indicus | October 2012 | 2 | 3 | del | 6 | 5950 | 10,971 | ||
| Narinda bay | Wild | F. indicus | December 2016 | 1 | 3 | del | 6 | 5950 | 10,971 |
| M. monoceros | December 2016 | 1 | 3 | del | 6 | 5950 | 10,971 | ||
| Mahajamba bay | P. monodon | December 2016 | 1 | 3 | del | 6 | 5950 | 10,971 | |
| M. monoceros | December 2016 | 1 | 3 | del | 6 | 5950 | 10,971 | ||
| Tsiribihina estuary | F. indicus | August 2016 | 8 | 3 | del | 6 | 5950 | 10,971 | |
| F. indicus | August 2016 | 4 | 3 | del | 8 | 5950 | 10,971 | ||
| M. monoceros | August 2016 | 3 | 3 | del | 6 | 5950 | 10,971 | ||
| M. japonicus | August 2016 | 1 | 3 | del | 6 | 5950 | 10,971 | ||
| M. rosenbergii | August 2016 | 2 | 3 | del | 6 | 5950 | 10,971 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onihary, A.M.; Razanajatovo, I.M.; Rabetafika, L.; Bastaraud, A.; Heraud, J.-M.; Rasolofo, V. Genotype Diversity and Spread of White Spot Syndrome Virus (WSSV) in Madagascar (2012–2016). Viruses 2021, 13, 1713. https://doi.org/10.3390/v13091713
Onihary AM, Razanajatovo IM, Rabetafika L, Bastaraud A, Heraud J-M, Rasolofo V. Genotype Diversity and Spread of White Spot Syndrome Virus (WSSV) in Madagascar (2012–2016). Viruses. 2021; 13(9):1713. https://doi.org/10.3390/v13091713
Chicago/Turabian StyleOnihary, Alain Moïse, Iony Manitra Razanajatovo, Lydia Rabetafika, Alexandra Bastaraud, Jean-Michel Heraud, and Voahangy Rasolofo. 2021. "Genotype Diversity and Spread of White Spot Syndrome Virus (WSSV) in Madagascar (2012–2016)" Viruses 13, no. 9: 1713. https://doi.org/10.3390/v13091713
APA StyleOnihary, A. M., Razanajatovo, I. M., Rabetafika, L., Bastaraud, A., Heraud, J.-M., & Rasolofo, V. (2021). Genotype Diversity and Spread of White Spot Syndrome Virus (WSSV) in Madagascar (2012–2016). Viruses, 13(9), 1713. https://doi.org/10.3390/v13091713