
Chronic Hepatitis B Treatment Strategies Using Polymerase Inhibitor-Based Combination Therapy


{kind=link}
{kind=link}
Abstract
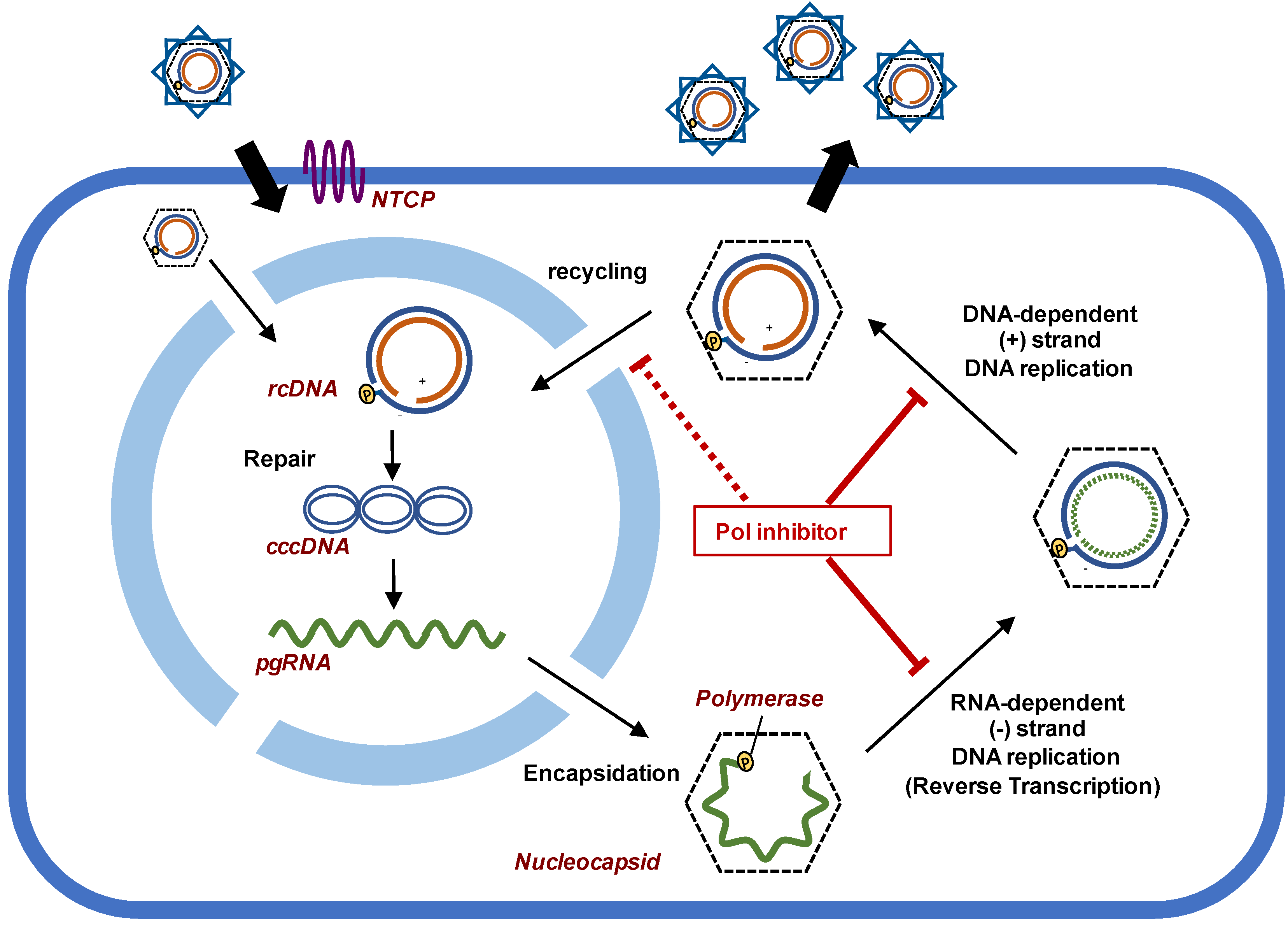
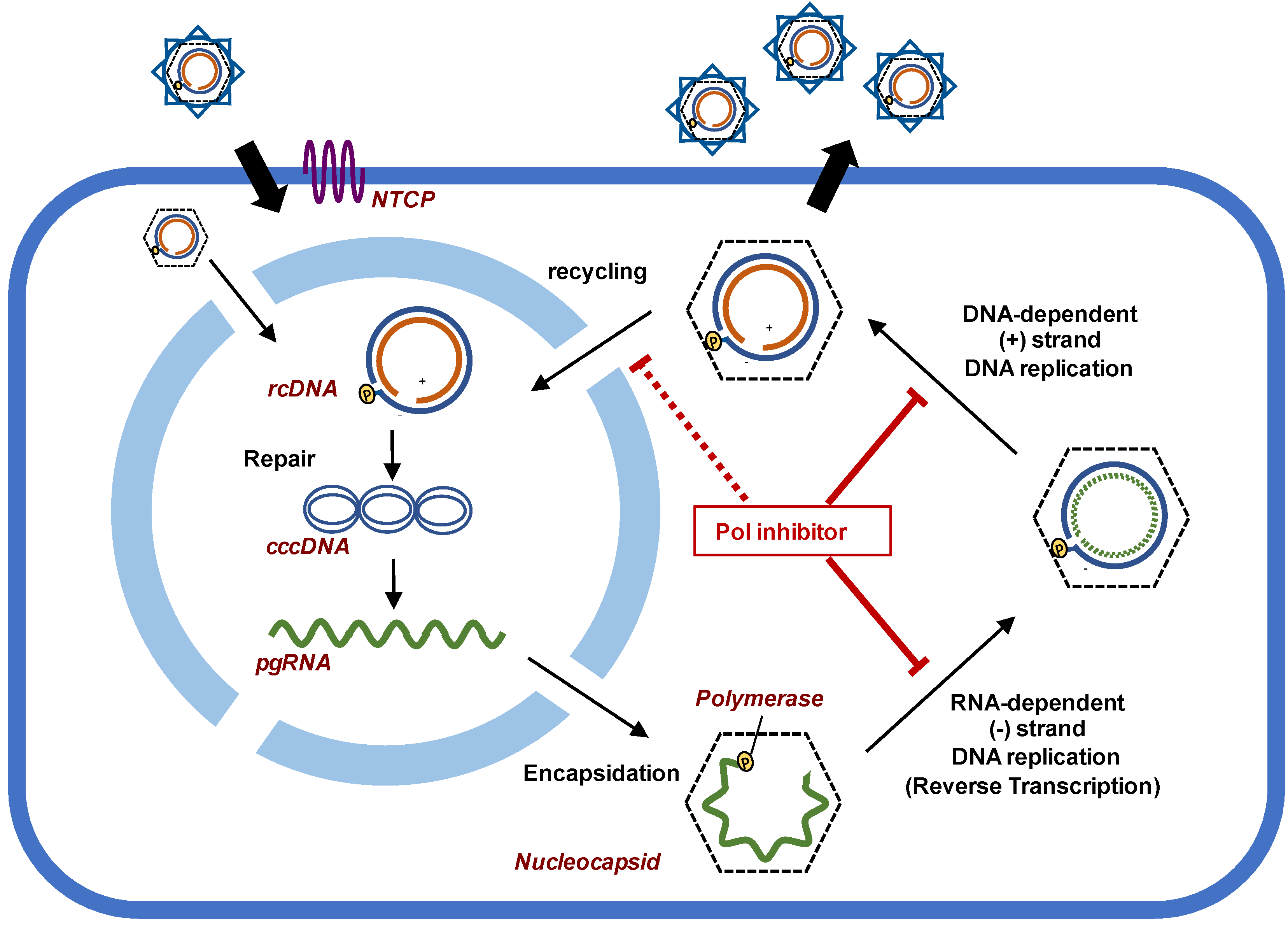
:1. Introduction
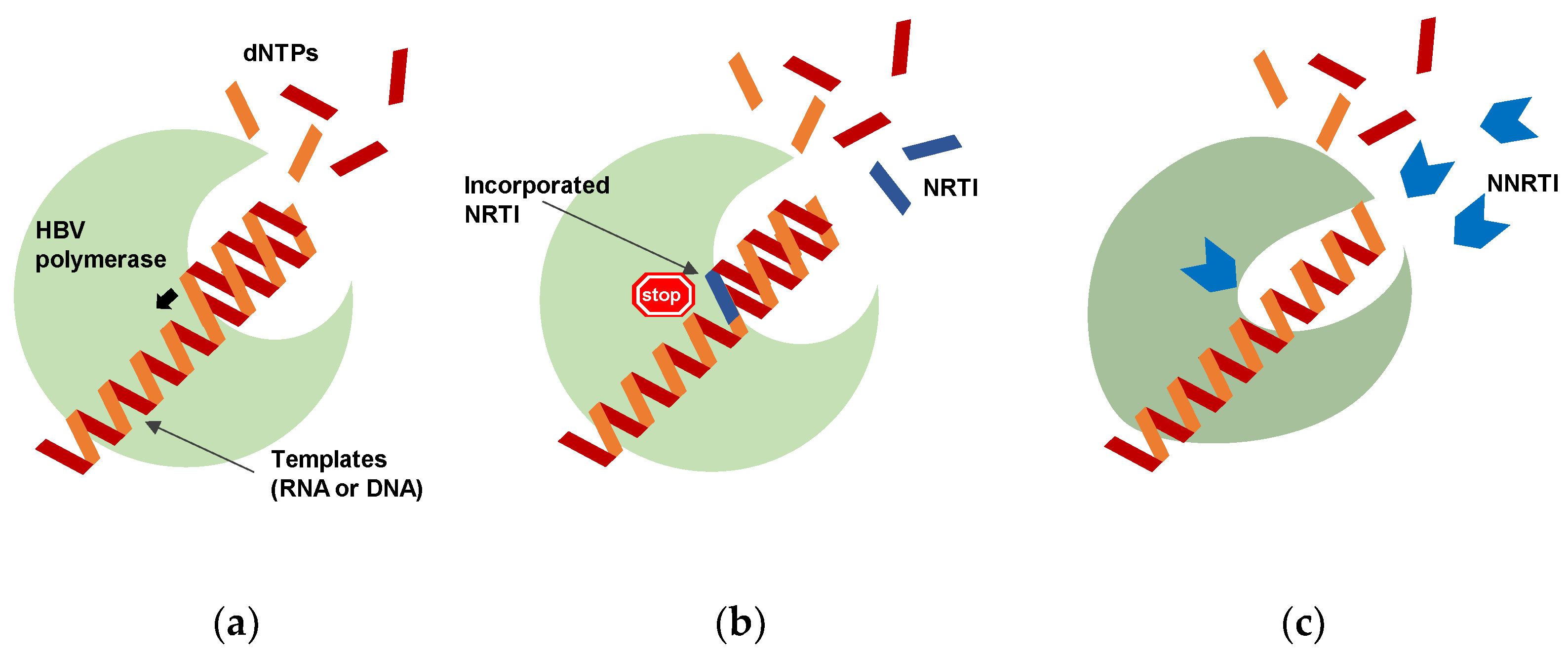
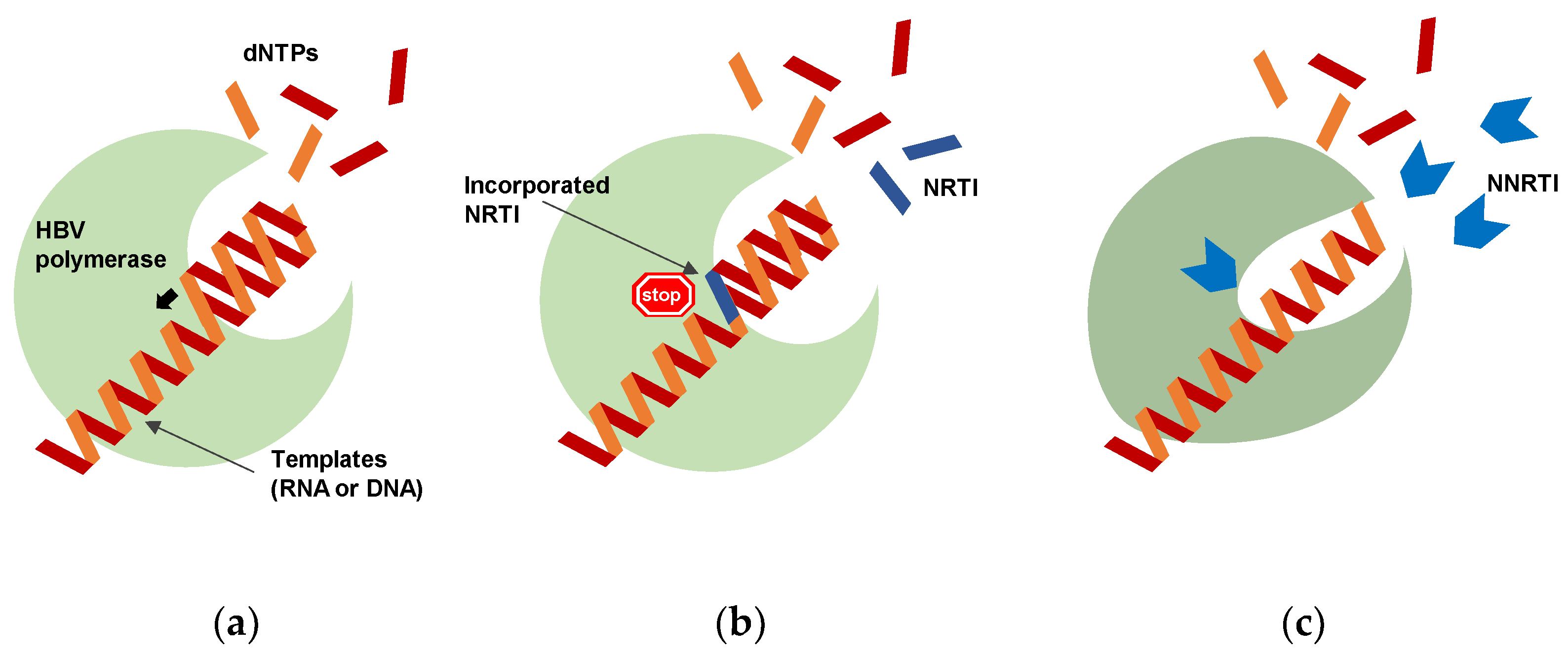
2. Potential of Polymerase Inhibitors
3. Combination Therapy Is a Prerequisite for the Elimination of Virus
3.1. Combination Therapies against Human Immunodeficiency Virus (HIV) and Hepatitis C Virus (HCV)
3.2. Combination with Interferon (IFN) and Nucleos(t)ide RT Inhibitors (NRTIs) for Hepatitis B Virus (HBV) Therapy
3.3. Combination Treatment with NRTIs and Other Agents
4. Discussion and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Lampertico, P.; Agarwal, K.; Berg, T.; Buti, M.; Janssen, H.L.A.; Papatheodoridis, G.; Zoulim, F.; Tacke, F. EASL 2017 Clinical Practice Guidelines on the Management of Hepatitis B Virus Infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucher, C.A.; Cammack, N.; Schipper, P.; Schuurman, R.; Rouse, P.; Wainberg, M.A.; Cameron, J.M. High-Level Resistance to (−) Enantiomeric 2′-Deoxy-3′-Thiacytidine In Vitro Is due to One Amino Acid Substitution in the Catalytic Site of Human Immunodeficiency Virus Type 1 Reverse Transcriptase. Antimicrob. Agents Chemother. 1993, 37, 2231–2234. [Google Scholar] [CrossRef] [Green Version]
- Cane, P.A.; Mutimer, D.; Ratcliffe, D.; Cook, P.; Beards, G.; Elias, E.; Pillay, D. Analysis of Hepatitis B Virus Quasispecies Changes during Emergence and Reversion of Lamivudine Resistance in Liver Transplantation. Antivir. Ther. 1999, 4, 7–14. [Google Scholar] [PubMed]
- Dienstag, J.L.; Perrillo, R.P.; Schiff, E.R.; Bartholomew, M.; Vicary, C.; Rubin, M. A Preliminary Trial of Lamivudine for Chronic Hepatitis B Infection. N. Engl. J. Med. 1995, 333, 1657–1661. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.A.; Bonhoeffer, S.; Hill, A.M.; Boehme, R.; Thomas, H.C.; McDade, H. Viral Dynamics in Hepatitis B Virus Infection. Proc. Natl. Acad. Sci. USA 1996, 93, 4398–4402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, T.M.; Locarnini, S.; McMahon, B.J.; Rehermann, B.; Peters, M.G. Use of Current and New Endpoints in the Evaluation of Experimental Hepatitis B Therapeutics. Clin. Infect. Dis. 2017, 64, 1283–1288. [Google Scholar] [CrossRef] [Green Version]
- Königer, C.; Wingert, I.; Marsmann, M.; Rösler, C.; Beck, J.; Nassal, M. Involvement of the Host DNA-Repair Enzyme TDP2 in Formation of the Covalently Closed Circular DNA Persistence Reservoir of Hepatitis B Viruses. Proc. Natl. Acad. Sci. USA 2014, 111, E4244–E4253. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Gao, Z.; Xu, G.; Peng, B.; Liu, C.; Yan, H.; Yao, Q.; Sun, G.; Liu, Y.; Tang, D.; et al. DNA Polymerase κ Is a Key Cellular Factor for the Formation of Covalently Closed Circular DNA of Hepatitis B Virus. PLoS Pathog. 2016, 12, e1005893. [Google Scholar] [CrossRef] [Green Version]
- Sheraz, M.; Cheng, J.; Tang, L.; Chang, J.; Guo, J.-T. Cellular DNA Topoisomerases Are Required for the Synthesis of Hepatitis B Virus Covalently Closed Circular DNA. J. Virol. 2019, 93, e02230-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, K.; Que, L.; Shimadu, M.; Koura, M.; Ishihara, Y.; Wakae, K.; Nakamura, T.; Watashi, K.; Wakita, T.; Muramatsu, M. Flap Endonuclease 1 Is Involved in CccDNA Formation in the Hepatitis B Virus. PLoS Pathog. 2018, 14, e1007124. [Google Scholar] [CrossRef]
- Mason, W.S.; Halpern, M.S.; England, J.M.; Seal, G.; Egan, J.; Coates, L.; Aldrich, C.; Summers, J. Experimental Transmission of Duck Hepatitis B Virus. Virology 1983, 131, 375–384. [Google Scholar] [CrossRef]
- Pourcel, C.; Summers, J. Formation of the Pool of Covalently Closed Circular Viral DNA in Hepadnavirus-Infected Cells. Cell 1986, 47, 451–460. [Google Scholar] [CrossRef]
- Wu, T.-T.; Coates, L.; Aldrics, C.E.; Summers, J.; Mason, W.S. In Hepatocytes Infected with Duck Hepatitis B Virus, the Template for Viral RNA Synthesis Is Amplified by an Intracellular Pathway. Virology 1990, 175, 255–261. [Google Scholar] [CrossRef]
- Ko, C.; Chakraborty, A.; Chou, W.-M.; Hasreiter, J.; Wettengel, J.M.; Stadler, D.; Bester, R.; Asen, T.; Zhang, K.; Wisskirchen, K.; et al. Hepatitis B Virus Genome Recycling and de Novo Secondary Infection Events Maintain Stable CccDNA Levels. J. Hepatol. 2018, 69, 1231–1241. [Google Scholar] [CrossRef]
- Wong, D.K.-H.; Seto, W.-K.; Cheung, K.-S.; Chong, C.-K.; Huang, F.-Y.; Fung, J.; Lai, C.-L.; Yuen, M.-F. Hepatitis B Virus Core-Related Antigen as a Surrogate Marker for Covalently Closed Circular DNA. Liver Int. 2017, 37, 995–1001. [Google Scholar] [CrossRef]
- Guo, F.; Zhao, Q.; Sheraz, M.; Cheng, J.; Qi, Y.; Su, Q.; Cuconati, A.; Wei, L.; Du, Y.; Li, W.; et al. HBV Core Protein Allosteric Modulators Differentially Alter CccDNA Biosynthesis from De Novo Infection and Intracellular Amplification Pathways. PLoS Pathog. 2017, 13, e1006658. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, V.L.; Feaver, W.J.; Fischhaber, P.L.; Friedberg, E.C. Purification and Characterization of Polκ, a DNA Polymerase Encoded by the Human DINB1 Gene. J. Biol. Chem. 2001, 276, 92–98. [Google Scholar] [CrossRef] [Green Version]
- García-Díaz, M.; Bebenek, K.; Sabariegos, R.; Domínguez, O.; Rodríguez, J.; Kirchhoff, T.; García-Palomero, E.; Picher, A.J.; Juárez, R.; Ruiz, J.F.; et al. DNA Polymerase λ, a Novel DNA Repair Enzyme in Human Cells. J. Biol. Chem. 2002, 277, 13184–13191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Sheraz, M.; McGrane, M.; Chang, J.; Guo, J.-T. DNA Polymerase Alpha Is Essential for Intracellular Amplification of Hepatitis B Virus Covalently Closed Circular DNA. PLoS Pathog. 2019, 15, e1007742. [Google Scholar] [CrossRef]
- Zhu, Y.; Yamamoto, T.; Cullen, J.; Saputelli, J.; Aldrich, C.E.; Miller, D.S.; Litwin, S.; Furman, P.A.; Jilbert, A.R.; Mason, W.S. Kinetics of Hepadnavirus Loss from the Liver during Inhibition of Viral DNA Synthesis. J. Virol. 2001, 75, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addison, W.R.; Walters, K.-A.; Wong, W.W.S.; Wilson, J.S.; Madej, D.; Jewell, L.D.; Tyrrell, D.L.J. Half-Life of the Duck Hepatitis B Virus Covalently Closed Circular DNA Pool In Vivo Following Inhibition of Viral Replication. J. Virol. 2002, 76, 6356–6363. [Google Scholar] [CrossRef] [Green Version]
- Cox, E.C. On the Organization of Higher Chromosomes. Nat. New Biol. 1972, 239, 133–134. [Google Scholar] [CrossRef] [PubMed]
- McVean, G.T.; Hurst, L.D. Evidence for a Selectively Favourable Reduction in the Mutation Rate of the X Chromosome. Nature 1997, 386, 388–392. [Google Scholar] [CrossRef]
- Smith, N.G.C.; Hurst, L.D. The Causes of Synonymous Rate Variation in the Rodent Genome: Can Substitution Rates Be Used to Estimate the Sex Bias in Mutation Rate? Genetics 1999, 152, 661–673. [Google Scholar] [CrossRef]
- Maepa, M.B.; Roelofse, I.; Ely, A.; Arbuthnot, P. Progress and Prospects of Anti-HBV Gene Therapy Development. Int. J. Mol. Sci. 2015, 16, 17589–17610. [Google Scholar] [CrossRef] [Green Version]
- Seeger, C. Complete Spectrum of CRISPR/Cas9-Induced Mutations on HBV CccDNA. Cell Ther. 2016, 24, 9. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K.; Maepa, M.B.; Ely, A.; Arbuthnot, P. Gene Therapy for Chronic HBV—Can We Eliminate CccDNA? Genes 2018, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Fanning, G.C.; Zoulim, F.; Hou, J.; Bertoletti, A. Therapeutic Strategies for Hepatitis B Virus Infection: Towards a Cure. Nat. Rev. Drug Discov. 2019, 18, 827–844. [Google Scholar] [CrossRef] [PubMed]
- Iannacone, M.; Guidotti, L.G. Immunobiology and Pathogenesis of Hepatitis B Virus Infection. Nat. Rev. Immunol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Gallois-Montbrun, S.; Schneider, B.; Chen, Y.; Giacomoni-Fernandes, V.; Mulard, L.; Morera, S.; Janin, J.; Deville-Bonne, D.; Veron, M. Improving Nucleoside Diphosphate Kinase for Antiviral Nucleotide Analogs Activation. J. Biol. Chem. 2002, 277, 39953–39959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sluis-Cremer, N.; Tachedjian, G. Mechanisms of Inhibition of HIV Replication by Non-Nucleoside Reverse Transcriptase Inhibitors. Virus Res. 2008, 134, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Deval, J. Antimicrobial Strategies: Inhibition of viral polymerase by 3′-hydroxyl nucleosides. Drugs 2009, 69, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Terrault, N.A.; Lok, A.S.F.; McMahon, B.J.; Chang, K.-M.; Hwang, J.P.; Jonas, M.M.; Brown, R.S.; Bzowej, N.H.; Wong, J.B. Update on Prevention, Diagnosis, and Treatment of Chronic Hepatitis B: AASLD 2018 Hepatitis B Guidance. Hepatology 2018, 67, 1560–1599. [Google Scholar] [CrossRef] [PubMed]
- Sarin, S.K.; Kumar, M.; Lau, G.K.; Abbas, Z.; Chan, H.L.Y.; Chen, C.J.; Chen, D.S.; Chen, H.L.; Chen, P.J.; Chien, R.N.; et al. Asian-Pacific Clinical Practice Guidelines on the Management of Hepatitis B: A 2015 Update. Hepatol. Int. 2016, 10, 1–98. [Google Scholar] [CrossRef] [PubMed]
- Werle–Lapostolle, B.; Bowden, S.; Locarnini, S.; Wursthorn, K.; Petersen, J.; Lau, G.; Trepo, C.; Marcellin, P.; Goodman, Z.; Delaney IV, W.E. Persistence of CccDNA during the Natural History of Chronic Hepatitis B and Decline during Adefovir Dipivoxil Therapy. Gastroenterology 2004, 126, 1750–1758. [Google Scholar] [CrossRef] [Green Version]
- Wursthorn, K.; Lutgehetmann, M.; Dandri, M.; Volz, T.; Buggisch, P.; Zollner, B.; Longerich, T.; Schirmacher, P.; Metzler, F.; Zankel, M.; et al. Peginterferon Alpha-2b plus Adefovir Induce Strong CccDNA Decline and HBsAg Reduction in Patients with Chronic Hepatitis B. Hepatology 2006, 44, 675–684. [Google Scholar] [CrossRef]
- Murata, K.; Asano, M.; Matsumoto, A.; Sugiyama, M.; Nishida, N.; Tanaka, E.; Inoue, T.; Sakamoto, M.; Enomoto, N.; Shirasaki, T.; et al. Induction of IFN-Λ3 as an Additional Effect of Nucleotide, Not Nucleoside, Analogues: A New Potential Target for HBV Infection. Gut 2018, 67, 362–371. [Google Scholar] [CrossRef]
- Yamada, N.; Murayama, A.; Shiina, M.; Aly, H.H.; Iwamoto, M.; Tsukuda, S.; Watashi, K.; Tanaka, T.; Moriishi, K.; Nishitsuji, H.; et al. Anti-viral Effects of Interferon-λ3 on Hepatitis B Virus Infection in Cell Culture. Hepatol. Res. 2020, 50, 283–291. [Google Scholar] [CrossRef]
- Das, K.; Martinez, S.E.; Bauman, J.D.; Arnold, E. HIV-1 Reverse Transcriptase Complex with DNA and Nevirapine Reveals Non-Nucleoside Inhibition Mechanism. Nat. Struct. Mol. Biol. 2012, 19, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsaki, E.; Ueda, K. Screening and Evaluation of Novel Compounds against Hepatitis B Virus Polymerase Using Highly Purified Reverse Transcriptase Domain. Viruses 2020, 12, 840. [Google Scholar] [CrossRef]
- Liu, X.; Ohsaki, E.; Ueda, K. Establishment of a System for Finding Inhibitors of ε RNA Binding with the HBV Polymerase. Genes Cells 2020, 25, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Chong, P.; Sebahar, P.; Youngman, M.; Garrido, D.; Zhang, H.; Stewart, E.L.; Nolte, R.T.; Wang, L.; Ferris, R.G.; Edelstein, M.; et al. Rational Design of Potent Non-Nucleoside Inhibitors of HIV-1 Reverse Transcriptase. J. Med. Chem. 2012, 55, 10601–10609. [Google Scholar] [CrossRef] [PubMed]
- Shafer, R.W.; Vuitton, D.A. Highly Active Antiretroviral Therapy (Haart) for the Treatment of Infection with Human Immunodeficiency Virus Type 1. Biomed. Pharmacother. 1999, 53, 73–86. [Google Scholar] [CrossRef]
- Brechtl, J.R.; Breitbart, W.; Galietta, M.; Krivo, S.; Rosenfeld, B. The Use of Highly Active Antiretroviral Therapy (HAART) in Patients with Advanced HIV Infection: Impact on Medical, Palliative Care, and Quality of Life Outcomes. J. Pain Symptom Manag. 2001, 21, 11. [Google Scholar] [CrossRef]
- Esposito, F.; Corona, A.; Tramontano, E. HIV-1 Reverse Transcriptase Still Remains a New Drug Target: Structure, Function, Classical Inhibitors, and New Inhibitors with Innovative Mechanisms of Actions. Mol. Biol. Int. 2012, 2012, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Kohlstaedt, L.A.; Wang, J.; Friedman, J.M.; Rice, P.A.; Steitz, T.A. Crystal Structure at 3.5Å Resolution of HIV-1 Reverse Transcriptase Complexed with an Inhibitor. Science 1992, 256, 9. [Google Scholar] [CrossRef] [Green Version]
- Rittinger, K.; Divita, G.; Goody, R.S. Human Immunodeficiency Virus Reverse Transcriptase Substrate-Induced Conformational Changes and the Mechanism of Inhibition by Nonnucleoside Inhibitors. Proc. Natl. Acad. Sci. USA 1995, 92, 8046–8049. [Google Scholar] [CrossRef] [Green Version]
- Esnouf, R.; Ren, J.; Ross, C.; Jones, Y.; Stammers, D.; Stuart, D. Mechanism of Inhibition of HIV-1 Reverse Transcriptase by Non-Nucleoside Inhibitors. Nat. Struct. Mol. Biol. 1995, 2, 303–308. [Google Scholar] [CrossRef]
- Yu, F.; Li, W.; Wang, L.; Dai, Y.; Lu, X.; Wang, Q.; Xie, L.; Jiang, S. Combining New Non-Nucleoside Reverse Transcriptase Inhibitors (RTIs) with AZT Results in Strong Synergism against Multi-RTI-Resistant HIV-1 Strains. Molecules 2018, 23, 1599. [Google Scholar] [CrossRef] [Green Version]
- Benítez-Gutiérrez, L.; Barreiro, P.; Labarga, P.; de Mendoza, C.; Fernandez-Montero, J.V.; Arias, A.; Peña, J.M.; Soriano, V. Prevention and Management of Treatment Failure to New Oral Hepatitis C Drugs. Expert Opin. Pharmacother. 2016, 17, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Buti, M.; Riveiro-Barciela, M.; Esteban, R. Management of Direct-Acting Antiviral Agent Failures. J. Hepatol. 2015, 63, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Jadoul, M.; Martin, P. Hepatitis C Treatment in Chronic Kidney Disease Patients: The Kidney Disease Improving Global Outcomes Perspective. Blood Purif. 2017, 43, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Lawitz, E.; Poordad, F.F.; Pang, P.S.; Hyland, R.H.; Ding, X.; Mo, H.; Symonds, W.T.; McHutchison, J.G.; Membreno, F.E. Sofosbuvir and Ledipasvir Fixed-Dose Combination with and without Ribavirin in Treatment-Naive and Previously Treated Patients with Genotype 1 Hepatitis C Virus Infection (LONESTAR): An Open-Label, Randomised, Phase 2 Trial. Lancet 2014, 383, 515–523. [Google Scholar] [CrossRef]
- Bourlière, M.; Bronowicki, J.-P.; de Ledinghen, V.; Hézode, C.; Zoulim, F.; Mathurin, P.; Tran, A.; Larrey, D.G.; Ratziu, V.; Alric, L.; et al. Ledipasvir-Sofosbuvir with or without Ribavirin to Treat Patients with HCV Genotype 1 Infection and Cirrhosis Non-Responsive to Previous Protease-Inhibitor Therapy: A Randomised, Double-Blind, Phase 2 Trial (SIRIUS). Lancet Infect. Dis. 2015, 15, 397–404. [Google Scholar] [CrossRef]
- Charlton, M.; Everson, G.T.; Flamm, S.L.; Kumar, P.; Landis, C.; Brown, R.S.; Fried, M.W.; Terrault, N.A.; O’Leary, J.G.; Vargas, H.E.; et al. Ledipasvir and Sofosbuvir plus Ribavirin for Treatment of HCV Infection in Patients with Advanced Liver Disease. Gastroenterology 2015, 149, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.N.; Liu, L.; Rodriquez, J.L.; Zhao, L.; Schuster, L.; Li, E.; Wang, G.P.; Neely, M.N.; Yamada, W.; Drusano, G.L. Sofosbuvir (SOF) Suppresses Ledipasvir (LDV)-Resistant Mutants during SOF/LDV Combination Therapy against Genotype 1b Hepatitis C Virus (HCV). Sci. Rep. 2017, 7, 14421. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, S.; Kudo, M.; Osaki, Y.; Matsuo, H.; Inuzuka, T.; Matsumoto, A.; Tanaka, E.; Sakurai, T.; Ueshima, K.; Inoue, T.; et al. Impact of Peginterferon Alpha-2b and Entecavir Hydrate Combination Therapy on Persistent Viral Suppression in Patients with Chronic Hepatitis B. J. Med. Virol. 2013, 85, 987–995. [Google Scholar] [CrossRef]
- Hagiwara, S.; Nishida, N.; Watanabe, T.; Ida, H.; Sakurai, T.; Ueshima, K.; Takita, M.; Komeda, Y.; Nishijima, N.; Osaki, Y.; et al. Sustained Antiviral Effects and Clearance of Hepatitis Surface Antigen after Combination Therapy with Entecavir and Pegylated Interferon in Chronic Hepatitis B. Antivir. Ther. 2018, 23, 513–521. [Google Scholar] [CrossRef]
- Zheng, C.; Yan, H.; Zeng, J.; Cai, S.; Wu, X. Comparison of Pegylated Interferon Monotherapy and de Novo Pegylated Interferon plus Tenofovir Combination Therapy in Patients with Chronic Hepatitis B. Infect. Drug Resist. 2019, 12, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yan, R.; Ru, G.Q.; Yu, L.L.; Yao, J.; Wang, H. Pegylated-Interferon Consolidation Treatment versus Nucleos(t)Ide Analogue Consolidation Treatment in Non-Cirrhotic Hepatitis B Patients with Hepatitis B e Antigen Seroconversion: An Open-Label Pilot Trial. Hepatol. Int. 2019, 13, 422–430. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, K.; Chen, W.; Liao, J.; Luo, X.; Chen, R. Switching to PegIFNα-2b Leads to HBsAg Loss in Patients with Low HBsAg Levels and HBV DNA Suppressed by NAs. Sci. Rep. 2017, 7, 13383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Campenhout, M.J.H.; Brouwer, W.P.; Xie, Q.; Guo, S.; Chi, H.; Qi, X.; Tabak, F.; Streinu-Cercel, A.; Wang, J.-Y.; Zhang, N.-P.; et al. Long-Term Follow-up of Patients Treated with Entecavir and Peginterferon Add-on Therapy for HBeAg-Positive Chronic Hepatitis B Infection: ARES Long-Term Follow-Up. J. Viral Hepat. 2019, 26, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Bourlière, M.; Rabiega, P.; Ganne-Carrie, N.; Serfaty, L.; Marcellin, P.; Barthe, Y.; Thabut, D.; Guyader, D.; Hezode, C.; Picon, M.; et al. Effect on HBs Antigen Clearance of Addition of Pegylated Interferon Alfa-2a to Nucleos(t)Ide Analogue Therapy versus Nucleos(t)Ide Analogue Therapy Alone in Patients with HBe Antigen-Negative Chronic Hepatitis B and Sustained Undetectable Plasma Hepatitis B Virus DNA: A Randomised, Controlled, Open-Label Trial. Lancet Gastroenterol. Hepatol. 2017, 2, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Curtis, M.; Qi, X.; Miller, M.D.; Borroto-Esoda, K. Anti-Hepatitis B Virus Activity in Vitro of Combinations of Tenofovir with Nucleoside/Nucleotide Analogues. Antivir. Chem. Chemother. 2009, 19, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Qiang, R.; Lu, J.; Tuo, X.; Yang, X.; Li, X. Enhanced Antiviral Benefit of Combination Therapy with Anti-HBV and Anti-PD1 GRNA/Cas9 Produces a Synergistic Antiviral Effect in HBV Infection. Mol. Immunol. 2021, 130, 7–13. [Google Scholar] [CrossRef]
- Li, X.; Ke, Z.; Lian, D.; Yuan, J.; Pan, Y. Combination of Saikosaponin c and Telbivudine Synergistically Enhances the Anti-HBV Activity. Inflamm. Res. 2020, 69, 545–547. [Google Scholar] [CrossRef]
- Pan, Y.; Ke, Z.; Ye, H.; Sun, L.; Ding, X.; Shen, Y.; Zhang, R.; Yuan, J. Saikosaponin C Exerts Anti-HBV Effects by Attenuating HNF1α and HNF4α Expression to Suppress HBV PgRNA Synthesis. Inflamm. Res. 2019, 68, 1025–1034. [Google Scholar] [CrossRef]
- Huang, Q.; Cai, D.; Yan, R.; Li, L.; Zong, Y.; Guo, L.; Mercier, A.; Zhou, Y.; Tang, A.; Henne, K.; et al. Preclinical Profile and Characterization of the Hepatitis B Virus Core Protein Inhibitor ABI-H0731. Antimicrob. Agents Chemother. 2020, 64, e01463-20. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, L.; Ye, C.; Chen, C.; Hang, S.; Zhu, Z.; Shen, H.; Huang, Z.; Chen, W.; Xue, Y. Evolution of Drug-Resistant Mutations in HBV Genomes in Patients with Treatment Failure during the Past Seven Years (2010–2016). Virus Genes 2018, 54, 41–47. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohsaki, E.; Suwanmanee, Y.; Ueda, K. Chronic Hepatitis B Treatment Strategies Using Polymerase Inhibitor-Based Combination Therapy. Viruses 2021, 13, 1691. https://doi.org/10.3390/v13091691
Ohsaki E, Suwanmanee Y, Ueda K. Chronic Hepatitis B Treatment Strategies Using Polymerase Inhibitor-Based Combination Therapy. Viruses. 2021; 13(9):1691. https://doi.org/10.3390/v13091691
Chicago/Turabian StyleOhsaki, Eriko, Yadarat Suwanmanee, and Keiji Ueda. 2021. "Chronic Hepatitis B Treatment Strategies Using Polymerase Inhibitor-Based Combination Therapy" Viruses 13, no. 9: 1691. https://doi.org/10.3390/v13091691
APA StyleOhsaki, E., Suwanmanee, Y., & Ueda, K. (2021). Chronic Hepatitis B Treatment Strategies Using Polymerase Inhibitor-Based Combination Therapy. Viruses, 13(9), 1691. https://doi.org/10.3390/v13091691