
Function of Host Protein Staufen1 in Rabies Virus Replication


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Inoculation
2.2. Construction of STAU1 Eukaryotic Expression Vector and Overexpression
2.3. Establishment of STAU1 Gene Interference in SH-SY-5Y Cell Line
2.4. Immunofluorescence Assay
2.5. TCID50 Assay
2.6. Western Blot
2.7. Total RNA Extraction, Reverse Transcription Quantitative Real-Time PCR
2.8. Statistical Analyssis
3. Results
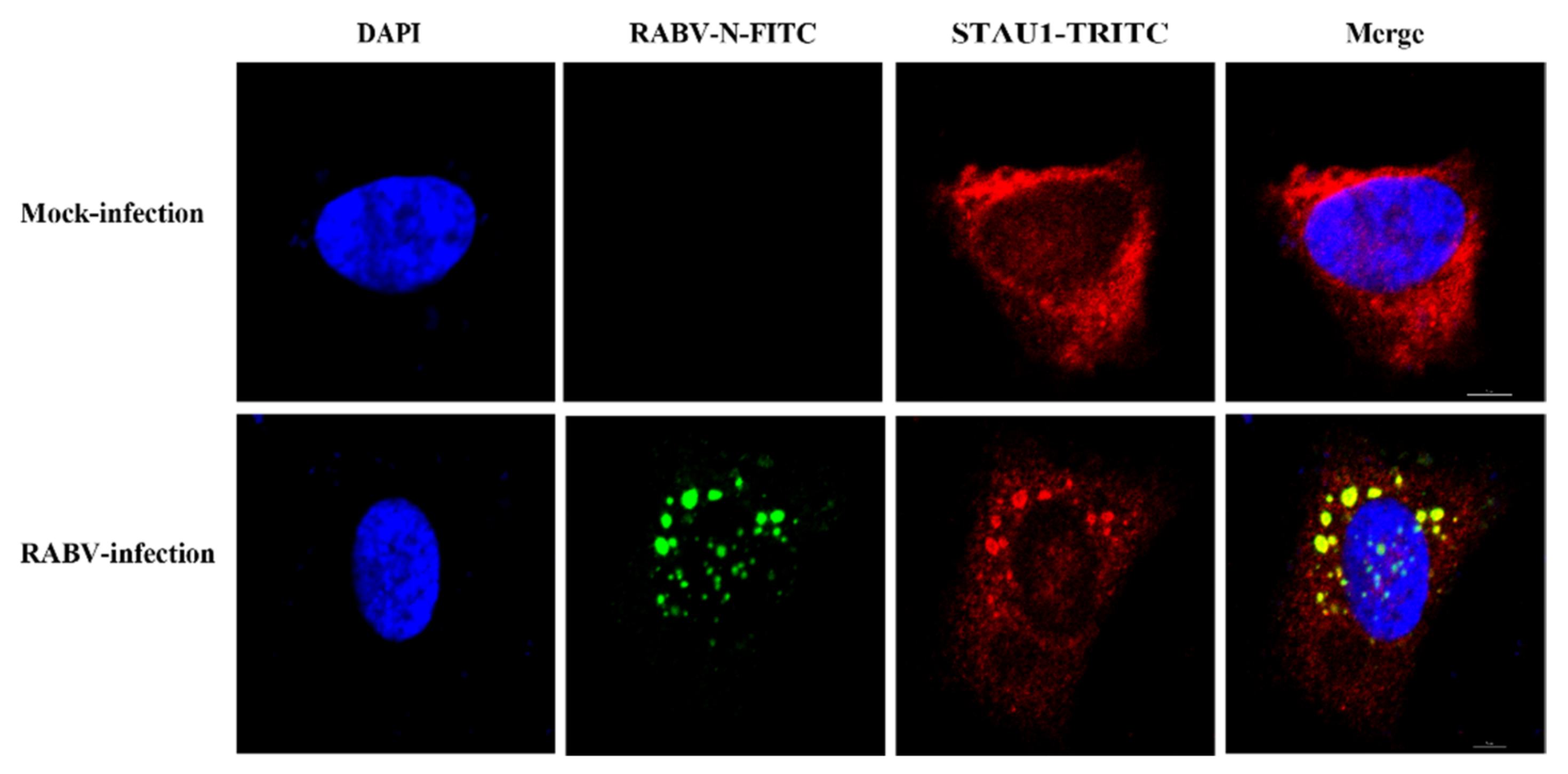
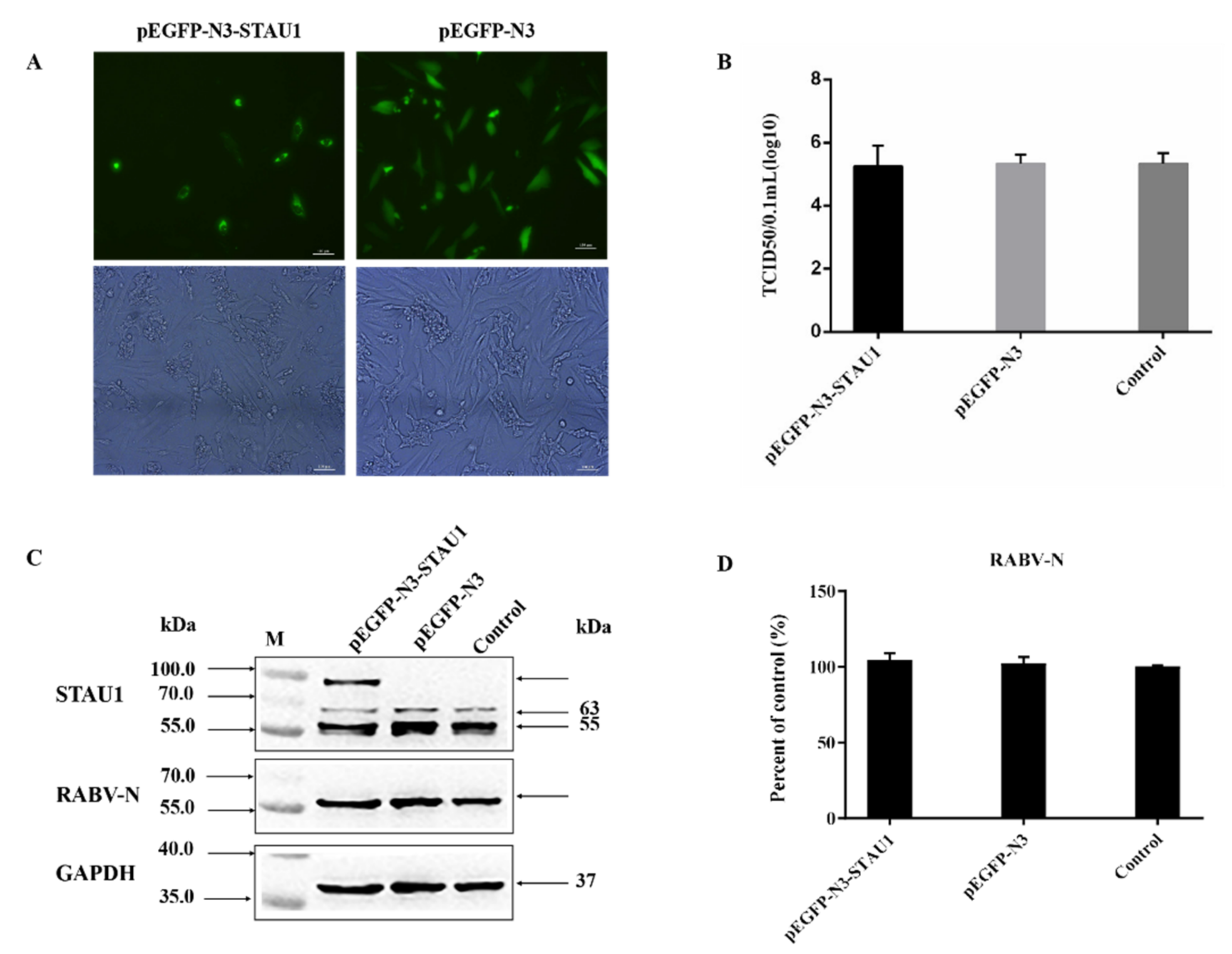
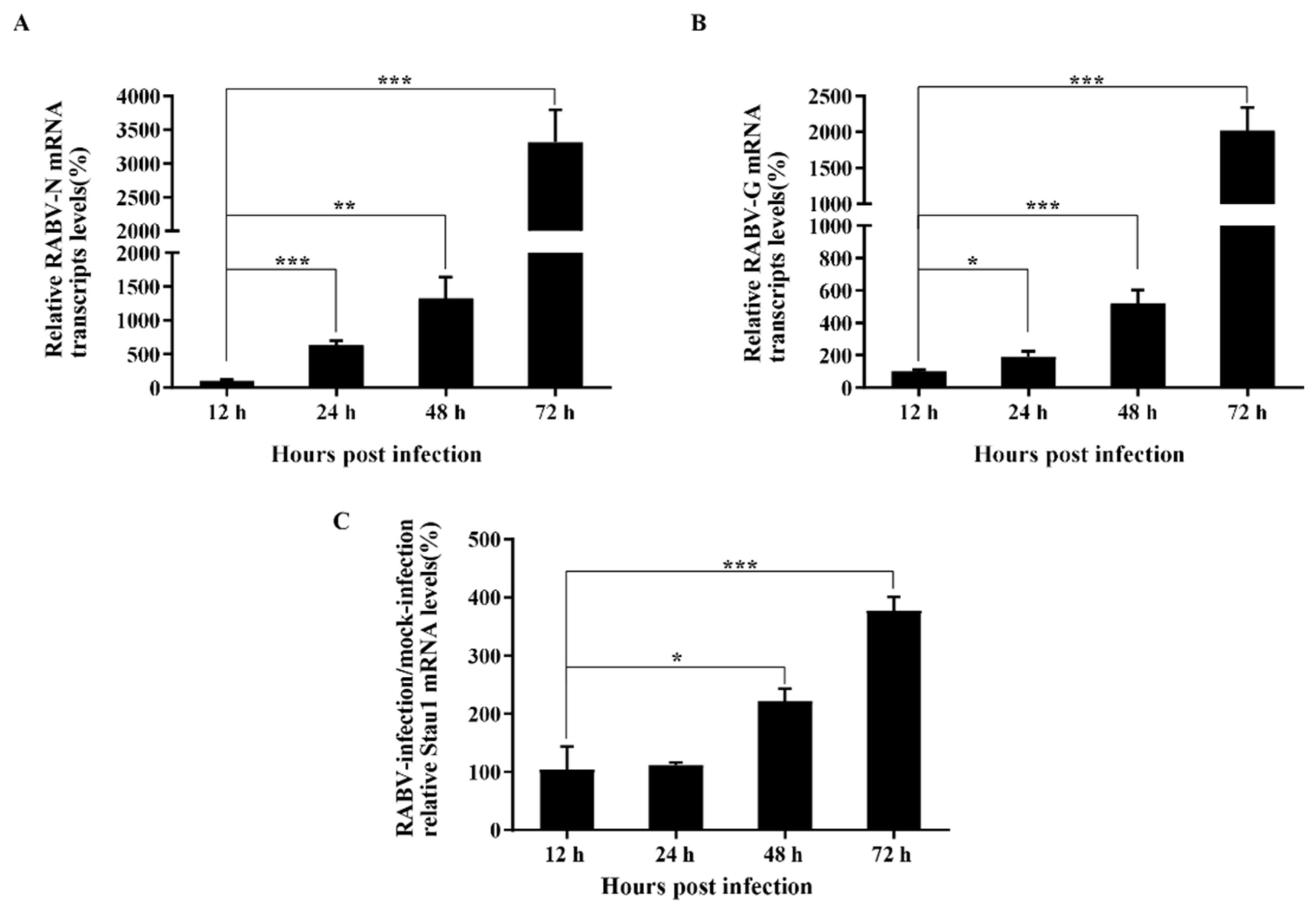
3.1. STAU1 Is Recruited to RABV Negri Bodies (NBs)
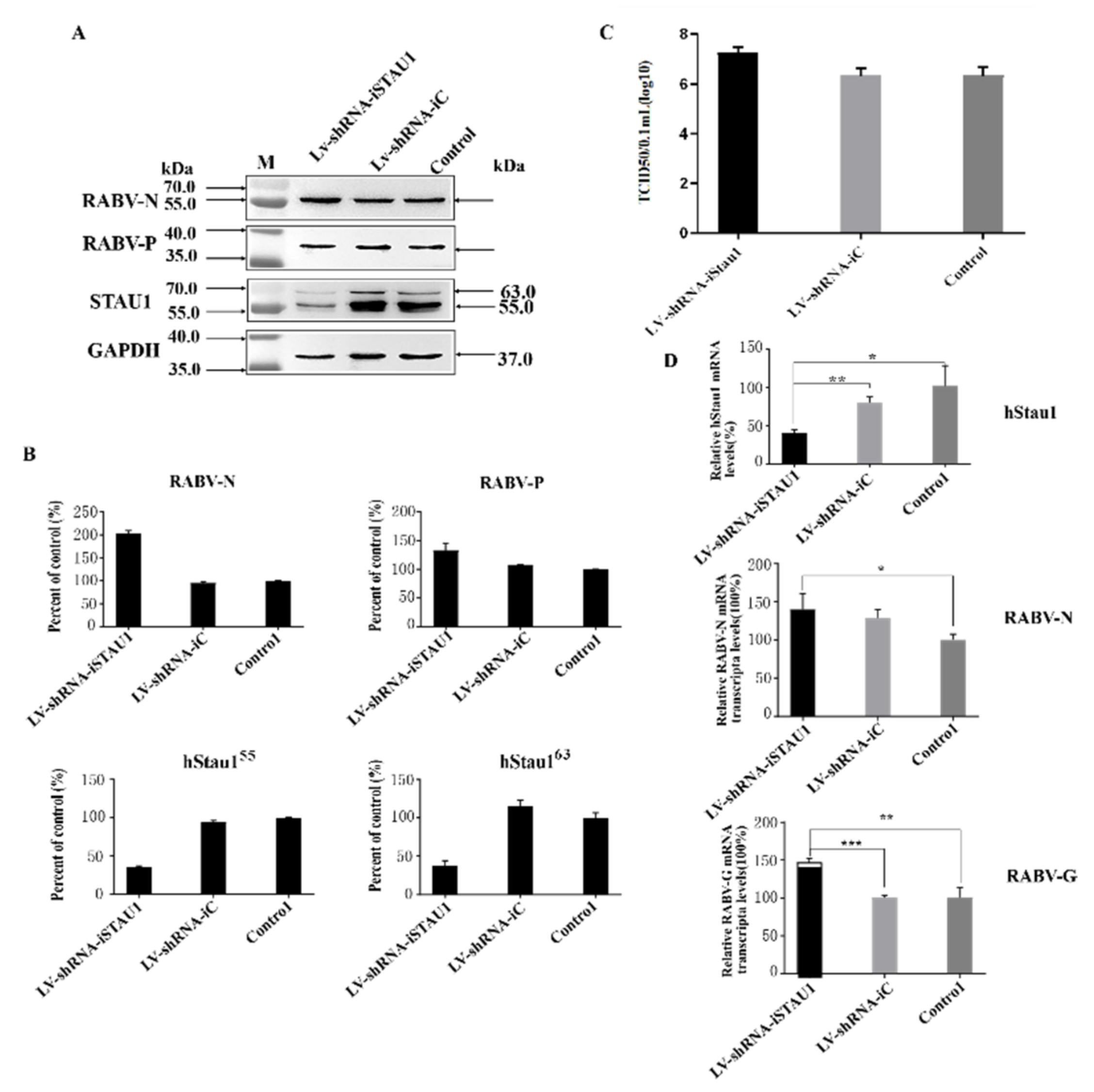
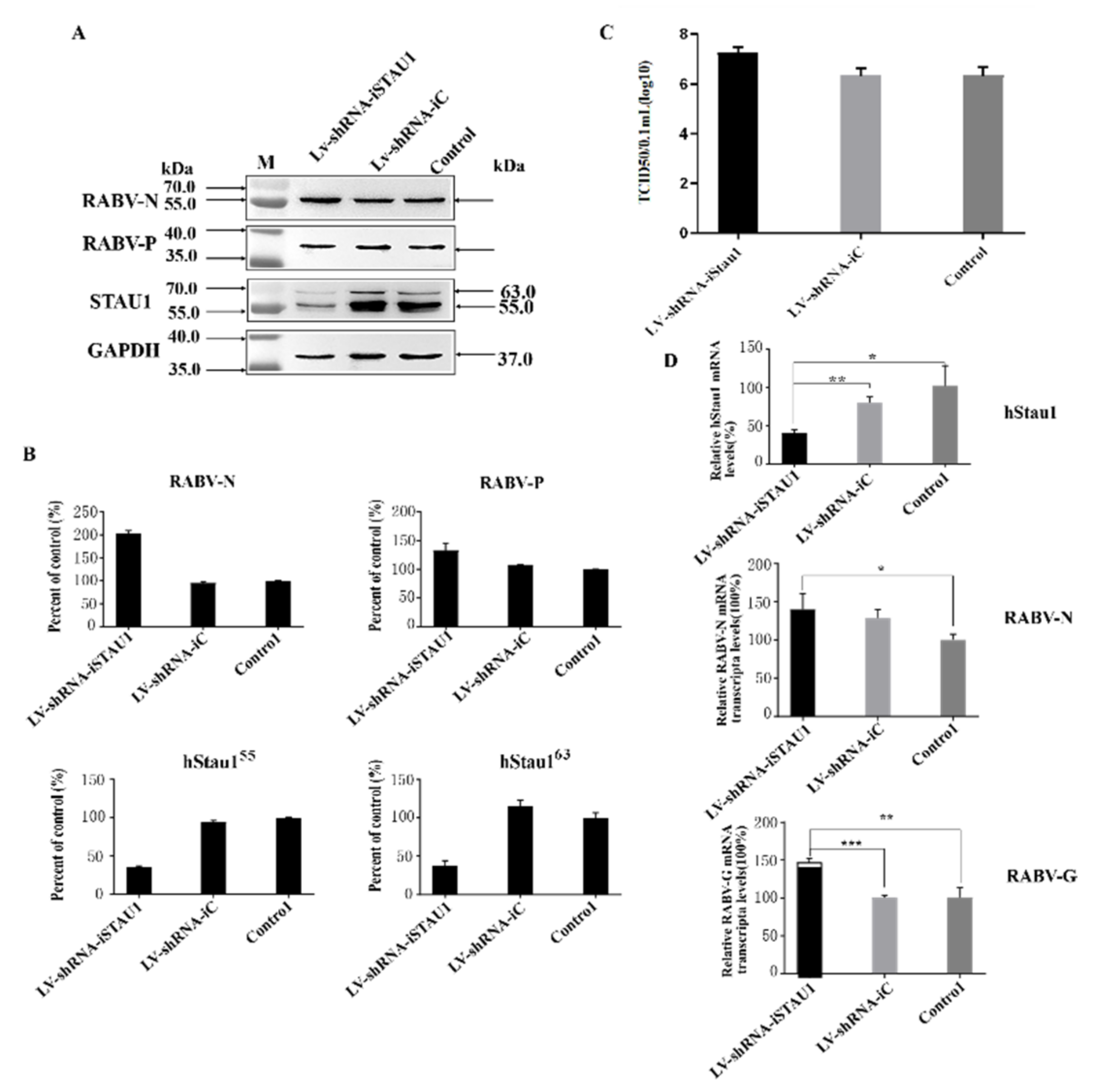
3.2. STAU1 Is Involved in Host Resistance to RABV Replication
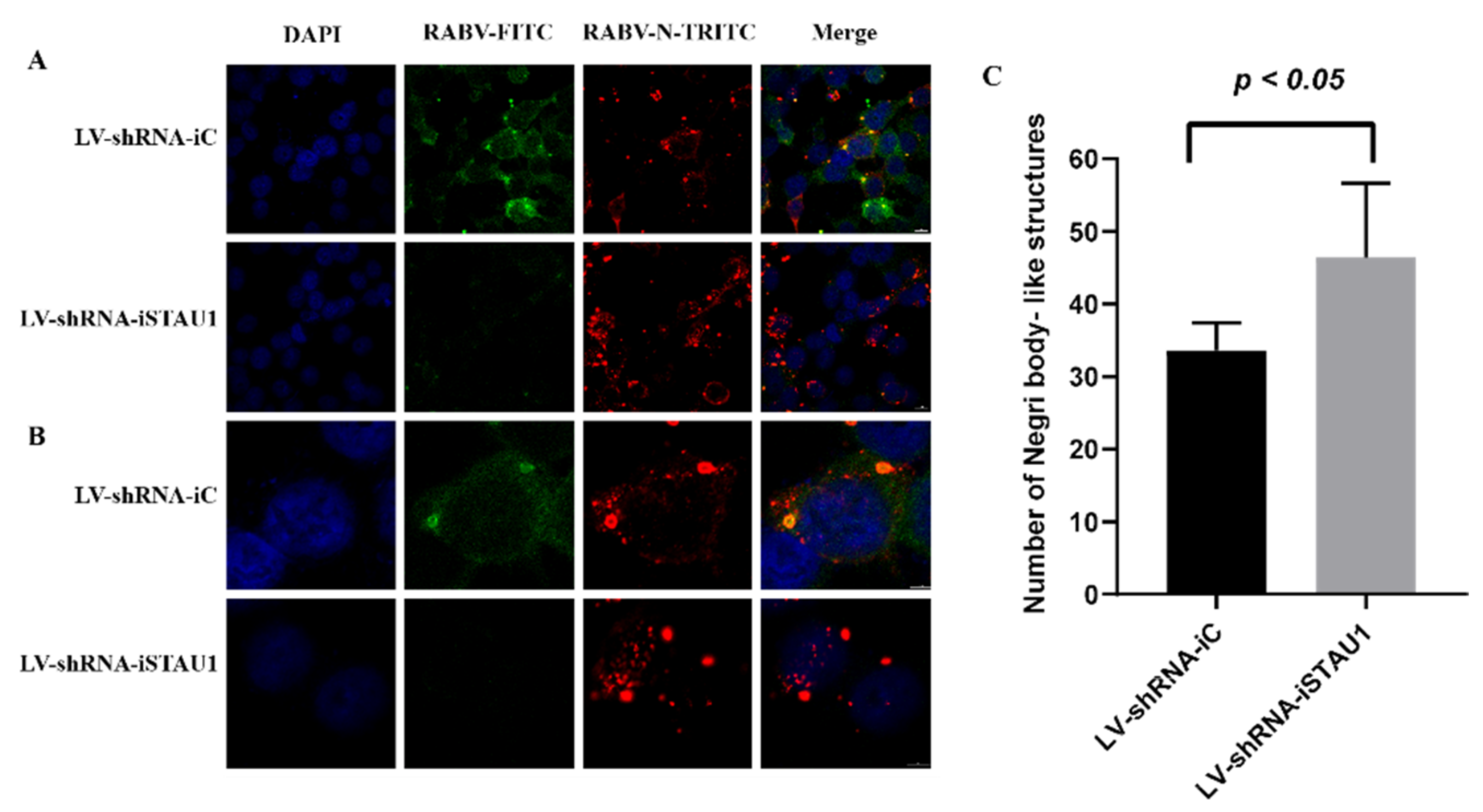
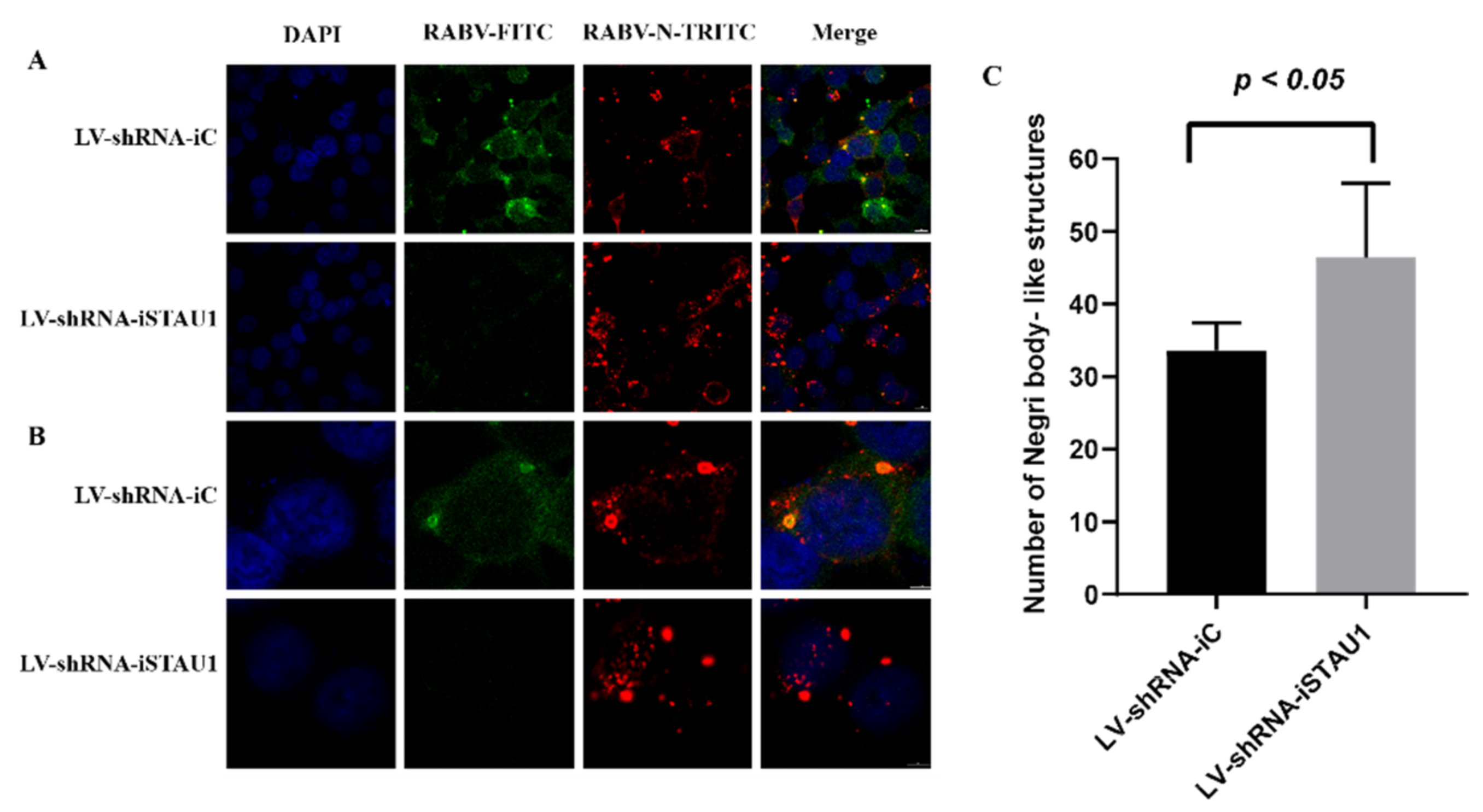
3.3. STAU1 Interferes with the Formation of Negri Bodies
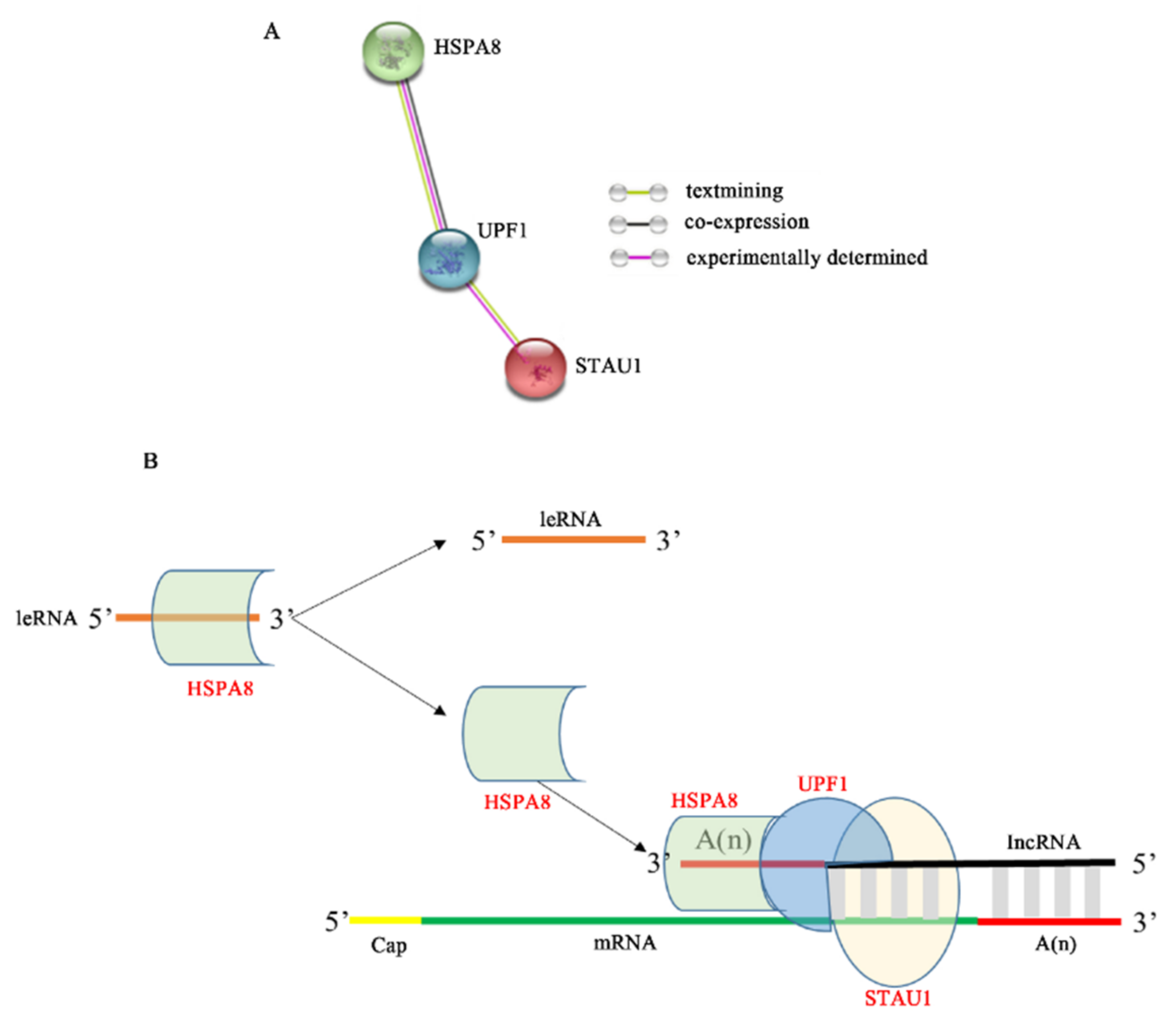
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Li, F.; Zhang, J.; Wang, L.; Wang, J.; Wen, Z.; Wang, Z.; Shuai, L.; Wang, X.; Ge, J.; et al. The ATPase ATP6V1A facilitates rabies virus replication by promoting virion uncoating and interacting with the viral matrix protein. J. Biol. Chem. 2021, 296. [Google Scholar] [CrossRef]
- Zan, J.; Liu, S.; Sun, D.N.; Mo, K.K.; Yan, Y.; Liu, J.; Hu, B.L.; Gu, J.Y.; Liao, M.; Zhou, J.Y. Rabies Virus Infection Induces Microtubule Depolymerization to Facilitate Viral RNA Synthesis by Upregulating HDAC6. Front. Cell Infect. Microbiol. 2017, 7, 146. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.L.; Nolan, T.; on behalf of the SAGE WG on Rabies. The WHO position on rabies immunization—2018 updates. Vaccine 2019, 37 (Suppl. 1), A85–A87. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Yuan, Y.; Yang, Y.; Luo, Z.; Sui, B.; Zhou, M.; Fu, Z.F.; Zhao, L. Interferon-Inducible GTPase 1 Impedes the Dimerization of Rabies Virus Phosphoprotein and Restricts Viral Replication. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Nikolic, J.; Lagaudriere-Gesbert, C.; Scrima, N.; Blondel, D.; Gaudin, Y. Structure and Function of Negri Bodies. Adv. Exp. Med. Biol. 2019, 1215, 111–127. [Google Scholar] [CrossRef]
- Nikolic, J.; Civas, A.; Lama, Z.; Lagaudriere-Gesbert, C.; Blondel, D. Rabies Virus Infection Induces the Formation of Stress Granules Closely Connected to the Viral Factories. PLoS Pathog. 2016, 12, e1005942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Realegeno, S.; Niezgoda, M.; Yager, P.A.; Kumar, A.; Hoque, L.; Orciari, L.; Sambhara, S.; Olson, V.A.; Satheshkumar, P.S. An ELISA-based method for detection of rabies virus nucleoprotein-specific antibodies in human antemortem samples. PLoS ONE 2018, 13, e0207009. [Google Scholar] [CrossRef]
- Lingappa, U.F.; Wu, X.; Macieik, A.; Yu, S.F.; Atuegbu, A.; Corpuz, M.; Francis, J.; Nichols, C.; Calayag, A.; Shi, H.; et al. Host-rabies virus protein-protein interactions as druggable antiviral targets. Proc. Natl. Acad. Sci. USA 2013, 110, E861–E868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zandi, F.; Khalaj, V.; Goshadrou, F.; Meyfour, A.; Gholami, A.; Enayati, S.; Mehranfar, M.; Rahmati, S.; Kheiri, E.V.; Badie, H.G.; et al. Rabies virus matrix protein targets host actin cytoskeleton: A protein-protein interaction analysis. Pathog. Dis. 2021, 79. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Y.; Huang, Y.; Gnanadurai, C.W.; Zhou, M.; Zhao, L.; Fu, Z.F. The ectodomain of rabies virus glycoprotein determines dendritic cell activation. Antivir. Res. 2017, 141, 1–6. [Google Scholar] [CrossRef]
- Nakagawa, K.; Kobayashi, Y.; Okada, K.; Makino, M.; Ito, N.; Goto, H.; Suzuki, Y.; Takahashi, T.; Sugiyama, M. Molecular Function Analysis of Rabies Virus RNA Polymerase L Protein by Using an L Gene-Deficient Virus. J. Virol. 2017, 91, e00826-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.; Gleghorn, M.L.; Maquat, L.E. Staufen2 functions in Staufen1-mediated mRNA decay by binding to itself and its paralog and promoting UPF1 helicase but not ATPase activity. Proc. Natl. Acad. Sci. USA 2013, 110, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Furic, L.; Desgroseillers, L.; Maquat, L.E. Mammalian Staufen1 recruits Upf1 to specific mRNA 3’ UTRs so as to elicit mRNA decay. Cell 2005, 120, 195–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, C.; Maquat, L.E. lncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 3’ UTRs via Alu elements. Nature 2011, 470, 284–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.; Maquat, L.E. Staufen-mediated mRNA decay. Wiley Interdiscip. Rev. RNA 2013, 4, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.; Tang, Y.; Maquat, L.E. mRNA-mRNA duplexes that autoelicit Staufen1-mediated mRNA decay. Nat. Struct. Mol. Biol. 2013, 20, 1214–1220. [Google Scholar] [CrossRef] [Green Version]
- Abrahamyan, L.G.; Chatel-Chaix, L.; Ajamian, L.; Milev, M.P.; Monette, A.; Clement, J.F.; Song, R.; Lehmann, M.; DesGroseillers, L.; Laughrea, M.; et al. Novel Staufen1 ribonucleoproteins prevent formation of stress granules but favour encapsidation of HIV-1 genomic RNA. J. Cell Sci. 2010, 123, 369–383. [Google Scholar] [CrossRef] [Green Version]
- Blackham, S.L.; McGarvey, M.J. A host cell RNA-binding protein, Staufen1, has a role in hepatitis C virus replication before virus assembly. J. Gen. Virol. 2013, 94, 2429–2436. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Ahn, S.H.; Kim, K.M.; Kim, Y.K. Non-structural protein 1 of influenza viruses inhibits rapid mRNA degradation mediated by double-stranded RNA-binding protein, staufen1. FEBS Lett. 2013, 587, 2118–2124. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.M.; Ou, B.T.; Chen, C.Y.; Chan, H.H.; Chen, C.J.; Wang, R.Y. Staufen1 Protein Participates Positively in the Viral RNA Replication of Enterovirus 71. Viruses 2019, 11, 142. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Pietzsch, C.; Ramanathan, P.; Santos, R.I.; Ilinykh, P.A.; Garcia-Blanco, M.A.; Bukreyev, A.; Bradrick, S.S. Staufen1 Interacts with Multiple Components of the Ebola Virus Ribonucleoprotein and Enhances Viral RNA Synthesis. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Yu, Z.; Xiong, Y.; Wang, Y.; Ruan, Y.; Guo, Y.; Chen, M.; Luan, S.; Zhang, E.; Liu, H. STAU1 binds to IBDV genomic double-stranded RNA and promotes viral replication via attenuation of MDA5-dependent beta interferon induction. FASEB J. 2019, 33, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Pan, T.; Fang, C.; Yi, Z.; Yang, P.; Yuan, Z. Subproteomic analysis of the cellular proteins associated with the 3’ untranslated region of the hepatitis C virus genome in human liver cells. Biochem. Biophys. Res. Commun. 2006, 347, 683–691. [Google Scholar] [CrossRef]
- Chatel-Chaix, L.; Boulay, K.; Mouland, A.J.; Desgroseillers, L. The host protein Staufen1 interacts with the Pr55Gag zinc fingers and regulates HIV-1 assembly via its N-terminus. Retrovirology 2008, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raoa, S.; Hassinec, S.; Monettea, A.; Amorima, R.; DesGroseillersc, L.; Moulanda, A.J. HIV-1 requires Staufen1 to dissociate stress granules and to produce infectious viral particles. RNA 2019, 25, 727–736. [Google Scholar] [CrossRef]
- Toro-Ascuy, D.; Rojas-Araya, B.; Valiente-Echeverria, F.; Soto-Rifo, R. Interactions between the HIV-1 Unspliced mRNA and Host mRNA Decay Machineries. Viruses 2016, 8, 320. [Google Scholar] [CrossRef] [PubMed]
- De Lucas, S.; Peredo, J.; Marion, R.M.; Sanchez, C.; Ortin, J. Human Staufen1 protein interacts with influenza virus ribonucleoproteins and is required for efficient virus multiplication. J. Virol. 2010, 84, 7603–7612. [Google Scholar] [CrossRef] [Green Version]
- Kristensson, K.; Dastur, D.K.; Manghani, D.K.; Tsiang, H.; Bentivoglio, M. Rabies: Interactions between neurons and viruses. A review of the history of Negri inclusion bodies. Neuropathol. Appl. Neurobiol. 1996, 22, 179–187. [Google Scholar] [CrossRef]
- Shin, T.; Weinstock, D.; Castro, M.D.; Hamir, A.N.; Wampler, T.; Walter, M.; Kim, H.Y.; Acland, H. Immunohistochemical localization of endothelial and inducible nitric oxide synthase within neurons of cattle with rabies. J. Vet. Med Sci. 2004, 66, 539–541. [Google Scholar] [CrossRef] [Green Version]
- Menager, P.; Roux, P.; Megret, F.; Bourgeois, J.P.; Le Sourd, A.M.; Danckaert, A.; Lafage, M.; Prehaud, C.; Lafon, M. Toll-like receptor 3 (TLR3) plays a major role in the formation of rabies virus Negri Bodies. PLoS Pathog. 2009, 5, e1000315. [Google Scholar] [CrossRef] [PubMed]
- Lahaye, X.; Vidy, A.; Fouquet, B.; Blondel, D. Hsp70 protein positively regulates rabies virus infection. J. Virol. 2012, 86, 4743–4751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouquet, B.; Nikolic, J.; Larrous, F.; Bourhy, H.; Wirblich, C.; Lagaudriere-Gesbert, C.; Blondel, D. Focal adhesion kinase is involved in rabies virus infection through its interaction with viral phosphoprotein P. J. Virol. 2015, 89, 1640–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Ye, C.; Ruan, X.; Zan, J.; Xu, Y.; Liao, M.; Zhou, J. The chaperonin CCTalpha is required for efficient transcription and replication of rabies virus. Microbiol. Immunol. 2014, 58, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, X.; Zan, J.; Wu, Y.; Ye, C.; Ruan, X.; Zhou, J. Cellular Chaperonin CCTγ Contributes to Rabies Virus Replication during Infection. J. Virol. 2013, 87, 7608–7621. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Han, Q.; Song, Y.; Qiang, C.; Xia, X. Analysis of Subcellular Prefoldin 1 Redistribution During Rabies Virus Infection. Jundishapur J. Microbiol. 2015, 8, e24757. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Jiang, T.; Zhong, Z.; Zhao, L.; Yang, S.; Xia, X. Inhibition of rabies virus replication by interferon-stimulated gene 15 and its activating enzyme UBA7. Infect. Genet. Evol. 2017, 56, 44–53. [Google Scholar] [CrossRef]
- Ricci, E.P.; Kucukural, A.; Cenik, C.; Mercier, B.C.; Singh, G.; Heyer, E.E.; Ashar-Patel, A.; Peng, L.; Moore, M.J. Staufen1 senses overall transcript secondary structure to regulate translation. Nat. Struct. Mol. Biol. 2014, 21, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Kim, K.M.; Han, S.; Choe, J.; Park, S.G.; Choi, S.S.; Kim, Y.K. Staufen1-mediated mRNA decay functions in adipogenesis. Mol. Cell 2012, 46, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Nikolic, J.; Le Bars, R.; Lama, Z.; Scrima, N.; Lagaudriere-Gesbert, C.; Gaudin, Y.; Blondel, D. Negri bodies are viral factories with properties of liquid organelles. Nat. Commun. 2017, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Lahaye, X.; Vidy, A.; Pomier, C.; Obiang, L.; Harper, F.; Gaudin, Y.; Blondel, D. Functional characterization of Negri bodies (NBs) in rabies virus-infected cells: Evidence that NBs are sites of viral transcription and replication. J. Virol. 2009, 83, 7948–7958. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Benjamin, B.; Balakrishnan, K.; Ghosh, P.; Banerjee, S. Human protein Staufen-2 promotes HIV-1 proliferation by positively regulating RNA export activity of viral protein Rev. Retrovirology 2014, 11, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Asgari, S. MicroRNA-like viral small RNA from Dengue virus 2 autoregulates its replication in mosquito cells. Proc. Natl. Acad. Sci. USA 2014, 111, 2746–2751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Rossi, J.J. MicroRNAs and their potential involvement in HIV infection. Trends Pharm. Sci. 2011, 32, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Liu, C.; Cao, Y.; Jamal, M.; Chen, X.; Zheng, J.; Li, L.; You, J.; Zhu, Q.; Liu, S.; et al. Rabies viruses leader RNA interacts with host Hsc70 and inhibits virus replication. Oncotarget 2017, 8, 43822–43837. [Google Scholar] [CrossRef]
- Lee, J.H.; Oh, J.Y.; Pascua, P.N.; Kim, E.G.; Choi, Y.K.; Kim, H.K. Impairment of the Staufen1-NS1 interaction reduces influenza viral replication. Biochem. Biophys. Res. Commun. 2011, 414, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Liu, S.; Zhong, Z.; Jiang, T.; Weng, R.; Xie, M.; Yang, S.; Xia, X. Analysis of expression profiles of long noncoding RNAs and mRNAs in brains of mice infected by rabies virus by RNA sequencing. Sci. Rep. 2018, 8, 11858. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Chen, C.; Xu, R.; Yang, M.; Han, Q.; Wang, B.; Song, Y.; Xia, X.; Zhang, J. Function of Host Protein Staufen1 in Rabies Virus Replication. Viruses 2021, 13, 1426. https://doi.org/10.3390/v13081426
Liu G, Chen C, Xu R, Yang M, Han Q, Wang B, Song Y, Xia X, Zhang J. Function of Host Protein Staufen1 in Rabies Virus Replication. Viruses. 2021; 13(8):1426. https://doi.org/10.3390/v13081426
Chicago/Turabian StyleLiu, Gaowen, Congjie Chen, Ruixian Xu, Ming Yang, Qinqin Han, Binghui Wang, Yuzhu Song, Xueshan Xia, and Jinyang Zhang. 2021. "Function of Host Protein Staufen1 in Rabies Virus Replication" Viruses 13, no. 8: 1426. https://doi.org/10.3390/v13081426
APA StyleLiu, G., Chen, C., Xu, R., Yang, M., Han, Q., Wang, B., Song, Y., Xia, X., & Zhang, J. (2021). Function of Host Protein Staufen1 in Rabies Virus Replication. Viruses, 13(8), 1426. https://doi.org/10.3390/v13081426