
Rodent-Borne Orthohantaviruses in Vietnam, Madagascar and Japan

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Trapping and Sample Collection
2.2. RNA Extraction and RT-PCR Analysis
2.3. Host Identification
2.4. Genetic and Phylogenetic Analysis
3. Results
3.1. Trapping Surveys
3.2. Hantavirus Screening and Sequence Analyses
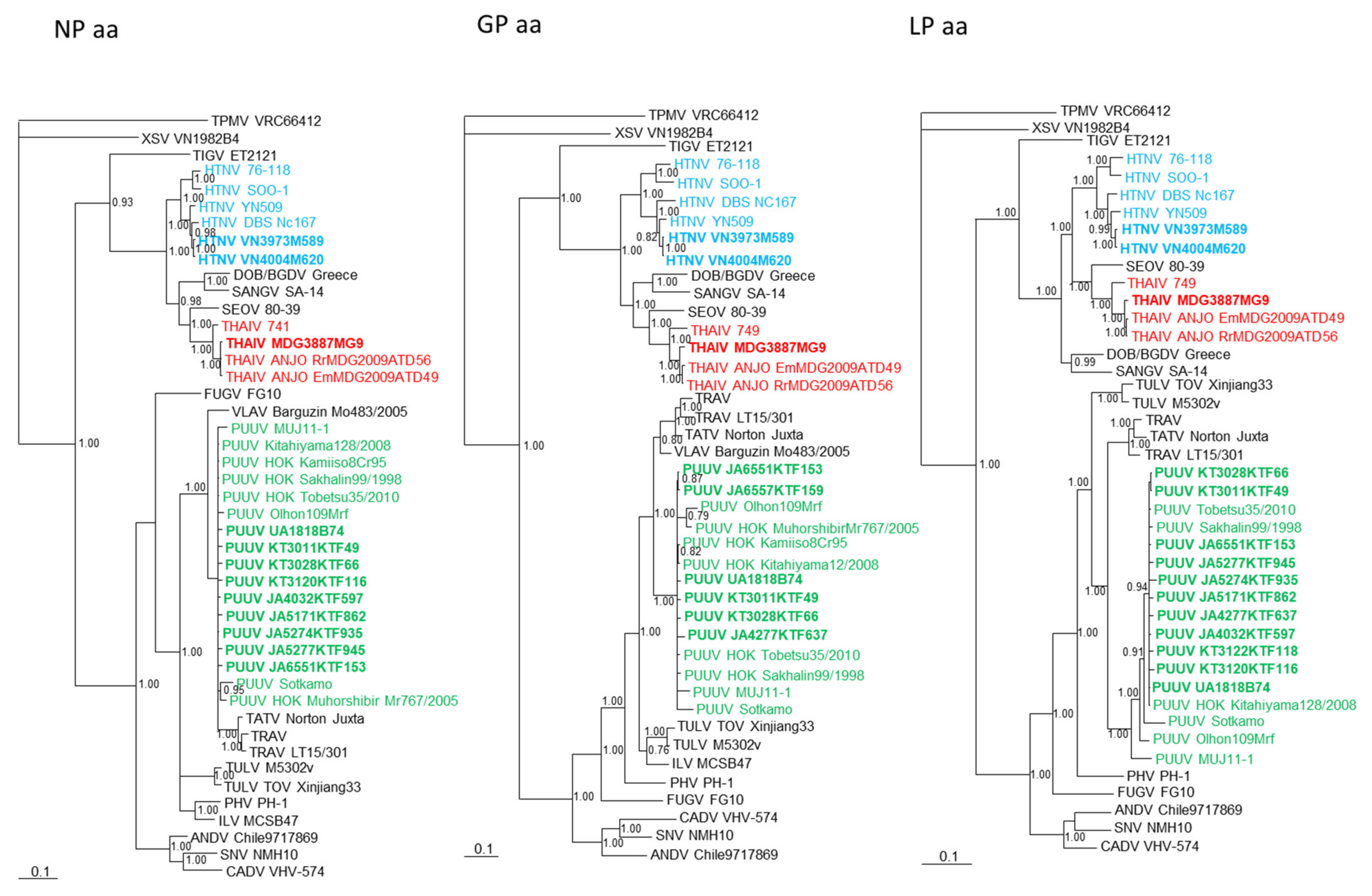
3.3. Phylogenetic Analysis and Host Species Identification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laenen, L.; Vergote, V.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Maes, P. Hantaviridae: Current classification and future perspectives. Viruses 2019, 11, 788. [Google Scholar] [CrossRef]
- Plyusnin, A.; Vapalahti, O.; Vaheri, A. Hantaviruses: Genome structure, expression and evolution. J. Gen. Virol. 1996, 77, 2677–2687. [Google Scholar] [CrossRef]
- Hepojoki, J.; Strandin, T.; Lankinen, H.; Vaheri, A. Hantavirus structure—Molecular interactions behind the scene. J. Gen. Virol. 2012, 93, 1631–1644. [Google Scholar] [CrossRef]
- Colleen, B.; Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef]
- Arai, S.; Yanagihara, R. Genetic diversity and geographic distribution of bat-borne hantaviruses. Curr. Issues Mol. Biol. 2020, 39, 1–28. [Google Scholar] [CrossRef]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the etiologic agent of Korean hemorrhagic fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef]
- Brummer-Korvenkontio, M.; Vaheri, A.; Hovi, T.; von Bonsdorff, C.H.; Vuorimies, J.; Manni, T.; Penttinen, K.; Oker-Blom, N.; Lähdevirta, J. Nephropathia epidemica: Detection of antigen in bank voles and serologic diagnosis of human infection. J. Infect. Dis. 1980, 141, 131–134. [Google Scholar] [CrossRef]
- Kariwa, H.; Yoshizumi, S.; Arikawa, J.; Yoshimatsu, K.; Takahashi, K.; Takashima, I.; Hashimoto, N. Evidence for the existence of Puumula-related virus among Clethrionomys rufocanus in Hokkaido, Japan. Am. J. Trop. Med. Hyg. 1995, 53, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Sanada, T.; Seto, T.; Ozaki, Y.; Saasa, N.; Yoshimatsu, K.; Arikawa, J.; Yoshii, K.; Kariwa, H. Isolation of Hokkaido virus, genus Hantavirus, using a newly established cell line derived from the kidney of the grey red-backed vole (Myodes rufocanus bedfordiae). J. Gen. Virol. 2012, 93, 2237–2246. [Google Scholar] [CrossRef] [PubMed]
- Yashina, L.N.; Abramov, S.A.; Dupal, T.A.; Danchinova, G.A.; Malyshev, B.S.; Hay, J.; Gu, S.H.; Yanagihara, R. Hokkaido genotype of Puumala virus in the grey red-backed vole (Myodes rufocanus) and northern red-backed vole (Myodes rutilus) in Siberia. Infect. Genet. Evol. 2015, 33, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Song, K.J.; Baek, L.J.; Moon, S.; Ha, S.J.; Kim, S.H.; Park, K.S.; Klein, T.A.; Sames, W.; Kim, H.C.; Lee, J.S.; et al. Muju virus, a novel hantavirus harboured by the arvicolid rodent Myodes regulus in Korea. J. Gen. Virol. 2007, 88, 3121–3129. [Google Scholar] [CrossRef]
- Lee, J.G.; Gu, S.H.; Baek, L.J.; Shin, O.S.; Park, K.S.; Kim, H.C.; Klein, T.A.; Yanagihara, R.; Song, J.-W. Muju virus, harbored by Myodes regulus in Korea, might represent a genetic variant of Puumala virus, the prototype arvicolid rodent-borne hantavirus. Viruses 2014, 6, 1701–1714. [Google Scholar] [CrossRef]
- Elwell, M.R.; Ward, G.S.; Tingpalapong, M.; Leduc, J.W. Serologic evidence of Hantaan-like virus in rodents and man in Thailand. S. Asian J. Trop. Med. Public Health 1985, 16, 349–354. [Google Scholar]
- Hugot, J.P.; Plyusnina, A.; Herbreteau, V.; Nemirov, K.; Laakkonen, J.; Lundkvist, A.; Supputamongkol, Y.; Henttonen, H.; Plyusnin, A. Genetic analysis of Thailand hantavirus in Bandicota indica trapped in Thailand. Virol. J. 2006, 3, 72. [Google Scholar] [CrossRef] [PubMed]
- Pattamadilok, S.; Lee, B.H.; Kumperasart, S.; Yoshimatsu, K.; Okumura, M.; Nakamura, I.; Araki, K.; Khoprasert, Y.; Dangsupa, P.; Panlar, P.; et al. Geographical distribution of hantaviruses in Thailand and potential human health significance of Thailand virus. Am. J. Trop. Med. Hyg. 2006, 75, 994–1002. [Google Scholar] [CrossRef]
- Plyusnina, A.; Ibrahim, I.N.; Plyusnin, A. A newly recognized hantavirus in the Asian house rat (Rattus tanezumi) in Indonesia. J. Gen. Virol. 2009, 90, 205–209. [Google Scholar] [CrossRef]
- Johansson, P.; Yap, G.; Low, H.T.; Siew, C.C.; Kek, R.; Ng, L.C.; Bucht, G. Molecular characterization of two hantavirus strains from different rattus species in Singapore. Virol. J. 2010, 7, 15. [Google Scholar] [CrossRef]
- Reynes, J.M.; Razafindralambo, N.K.; Lacoste, V.; Olive, M.M.; Barivelo, T.A.; Soarimalala, V.; Heraud, J.M.; Lavergne, A. Anjozorobe hantavirus, a new genetic variant of Thailand virus detected in rodents from Madagascar. Vector Borne Zoonotic Dis. 2014, 14, 212–219. [Google Scholar] [CrossRef]
- Ramsden, C.; Holmes, E.C.; Charleston, M.A. Hantavirus evolution in relation to its rodent and insectivore hosts: No evidence for codivergence. Mol. Biol. Evol. 2009, 26, 143–153. [Google Scholar] [CrossRef]
- Bennett, S.N.; Gu, S.H.; Kang, H.J.; Arai, S.; Yanagihara, R. Reconstructing the evolutionary origins and phylogeography of hantaviruses. Trends Microbiol. 2014, 22, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Guterres, A.; de Oliveira, R.C.; Fernandes, J.; de Lemos, E.R.S. The mystery of the phylogeographic structural pattern in rodent-borne hantaviruses. Mol. Phylogenet. Evol. 2019, 136, 35–43. [Google Scholar] [CrossRef]
- Sikes, R.S. Animal Care and Use Committee of the American Society of Mammalogists. 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, G.L., Jr. Guidelines for the Capture, Handling, and Care of Mammals as Approved by the American Society of Mammalogists. J. Mammal. 1998, 79, 1416–1431. [Google Scholar] [CrossRef]
- Arai, S.; Kang, H.J.; Gu, S.H.; Ohdachi, S.D.; Cook, J.A.; Yashina, L.N.; Tanaka-Taya, K.; Abramov, S.A.; Morikawa, S.; Okabe, N.; et al. Genetic diversity of Artybash virus in the Laxmann’s shrew (Sorex caecutiens). Vector Borne Zoonotic Dis. 2016, 16, 468–475. [Google Scholar] [CrossRef]
- Arai, S.; Ohdachi, S.D.; Asakawa, M.; Kang, H.J.; Mocz, G.; Arikawa, J.; Okabe, N.; Yanagihara, R. Molecular phylogeny of a newfound hantavirus in the Japanese shrew mole (Urotrichus talpoides). Proc. Natl. Acad. Sci. USA 2008, 105, 16296–16301. [Google Scholar] [CrossRef]
- Song, J.-W.; Gu, S.H.; Bennett, S.N.; Arai, S.; Puorger, M.; Hilbe, M.; Yanagihara, R. Seewis virus, a genetically distinct hantavirus in the Eurasian common shrew (Sorex araneus). Virol. J. 2007, 4, 114. [Google Scholar] [CrossRef]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Barriere, P.; Koivogui, L.; ter Meulen, J.; Krüger, D.H. Novel hantavirus sequences in shrew, Guinea. Emerg. Infect. Dis. 2007, 13, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Denys, C.; Koivogui, L.; ter Meulen, J.; Krüger, D.H. Hantavirus in African wood mouse, Guinea. Emerg. Infect. Dis. 2006, 12, 838–840. [Google Scholar] [CrossRef]
- Song, J.-W.; Kang, H.J.; Song, K.J.; Truong, T.T.; Bennett, S.N.; Arai, S.; Truong, N.U.; Yanagihara, R. Newfound hantavirus in Chinese mole shrew, Vietnam. Emerg. Infect. Dis. 2007, 13, 1784–1787. [Google Scholar] [CrossRef]
- Arai, S.; Taniguchi, S.; Aoki, K.; Yoshikawa, Y.; Kyuwa, S.; Tanaka-Taya, K.; Masangkay, J.S.; Omatsu, T.; Puentespina, R., Jr.; Watanabe, S.; et al. Molecular phylogeny of a genetically divergent hantavirus harbored by the Geoffroy’s rousette (Rousettus amplexicaudatus), a frugivorous bat species in the Philippines. Infect. Genet. Evol. 2016, 45, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, F.; Aoki, K.; Ohdachi, S.D.; Tsuchiya, K.; Motokawa, M.; Jogahara, T.; Son, N.T.; Bawm, S.; Lin, K.S.; Thwe, T.L.; et al. Genetic diversity and phylogeography of Thottapalayam thottimvirus (Hantaviridae) in Asian house shrew (Suncus murinus) in Eurasia. Front. Cell. Infect. Microbiol. 2020, 10, 438. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Tippmann, H.F. Analysis for free: Comparing programs for sequence analysis. Brief. Bioinform. 2004, 5, 82–87. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Jackson, A.P.; Charleston, M.A. A cophylogenetic perspective of RNA-virus evolution. Mol. Biol. Evol. 2004, 21, 45–57. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Charleston, M.A.; Robertson, D.L. Preferential host switching by primate lentiviruses can account for phylogenetic similarity with the primate phylogeny. Syst. Biol. 2002, 51, 528–535. [Google Scholar] [CrossRef]
- Lokugamage, N.; Kariwa, H.; Lokugamage, K.; Iwasa, M.A.; Hagiya, T.; Yoshii, K.; Tachi, A.; Ando, S.; Fukushima, H.; Tsuchiya, K.; et al. Epizootiological and epidemiological study of hantavirus infection in Japan. Microbiol. Immunol. 2004, 48, 843–851. [Google Scholar] [CrossRef]
- Lee, B.H.; Yoshimatsu, K.; Araki, K.; Ogino, M.; Okumura, M.; Tsuchiya, K.; Kariwa, H.; Arikawa, J. Detection of antibody for the serodiagnosis of hantavirus infection in different rodent species. Arch. Virol. 2003, 148, 1885–1897. [Google Scholar] [CrossRef]
- Ge, D.; Lu, L.; Abramov, A.V.; Wen, Z.; Cheng, J.; Xia, L.; Vogler, A.P.; Yang, Q. Coalescence models reveal the rise of the white-bellied rat (Niviventer confucianus) following the loss of Asian megafauna. J. Mammal. Evol. 2019, 26, 423–434. [Google Scholar] [CrossRef]
- Balakirev, A.E.; Abramov, A.V.; Rozhnov, V.V. Phylogenetic relationships in the Niviventer-Chiromyscus complex (Rodentia, Muridae) inferred from molecular data, with description of a new species. ZooKeys 2014, 451, 109–136. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.W.; Zuo, S.Q.; Gong, Z.D.; Zhan, L.; Bian, C.L.; Zhang, P.H.; Yang, H.; Zhang, J.S.; Zhao, Q.M.; Jia, N.; et al. Genetic analysis of a hantavirus strain carried by Niviventer confucianus in Yunnan province, China. Virus Res. 2010, 153, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Raharinosy, V.; Olive, M.M.; Andriamiarimanana, F.M.; Andriamandimby, S.F.; Ravalohery, J.P.; Andriamamonjy, S.; Filippone, C.; Rakoto, D.A.D.; Telfer, S.; Heraud, J.M. Geographical distribution and relative risk of Anjozorobe virus (Thailand orthohantavirus) infection in black rats (Rattus rattus) in Madagascar. Virol. J. 2018, 15, 83. [Google Scholar] [CrossRef] [PubMed]
- Hurles, M.E.; Sykes, B.C.; Jobling, M.A.; Forster, P. The dual origin of the Malagasy in Island Southeast Asia and East Africa: Evidence from maternal and paternal lineages. Am. J. Hum. Genet. 2005, 76, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Ranorosoa, M.C.; Kinoshita, G.; Shimoji, H.; Tsuchiya, K.; Ohdachi, S.D.; Arai, S.; Tanaka, C.; Ramino, H.; Suzuki, H. Variation in the coat-color-controlling genes, Mc1r and Asip, in the house mouse Mus musculus from Madagascar. Mammal Study 2016, 41, 131–140. [Google Scholar] [CrossRef]
- Heiske, M.; Alva, O.; Pereda-Loth, V.; Van Schalkwyk, M.; Radimilahy, C.; Letellier, T.; Rakotarisoa, J.-A.; Pierron, D. Genetic evidence and historical theories of the Asian and African origins of the present Malagasy population. Hum. Mol. Genet. 2021, 30, R72–R78. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.Y.; Diglisic, G.; Avšič-Županc, T.; LeDuc, J.W. Dobrava virus as a new hantavirus: Evidenced by comparative sequence analysis. J. Med. Virol. 1993, 39, 152–155. [Google Scholar] [CrossRef]
- Plyusnina, A.; Ferenczi, E.; Racz, G.R.; Nemirov, K.; Lundkvist, A.; Vaheri, A.; Vapalahti, O.; Plyusnin, A. Co-circulation of three pathogenic hantaviruses: Puumala, Dobrava, and Saaremaa in Hungary. J. Med. Virol. 2009, 81, 2045–2052. [Google Scholar] [CrossRef]
- Dzagurova, T.K.; Witkowski, P.T.; Tkachenko, E.A.; Klempa, B.; Morozov, V.G.; Auste, B.; Zavora, D.; Iunicheva, I.V.; Mutnih, E.S.; Krüger, D.H. Isolation of Sochi virus from a fatal case of hantavirus disease with fulminant clinical course. Clin. Infect. Dis. 2012, 54, e1–e4. [Google Scholar] [CrossRef]
- Yashina, L.N.; Abramov, S.A.; Gutorov, V.V.; Dupal, T.A.; Krivopalov, A.V.; Panov, V.V.; Danchinova, G.A.; Vinogradov, V.V.; Luchnikova, E.M.; Hay, J.; et al. Seewis virus: Phylogeography of a shrew-borne hantavirus in Siberia, Russia. Vector Borne Zoonotic Dis. 2010, 10, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.H.; Hejduk, J.; Markowski, J.; Kang, H.J.; Markowski, M.; Połatyńska, M.; Sikorska, B.; Liberski, P.P.; Yanagihara, R. Co-circulation of soricid- and talpid-borne hantaviruses in Poland. Infect. Genet. Evol. 2014, 28, 296–303. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Japan | Madagascar | Myanmar | Vietnam | Total | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2014 | 2015 | 2012 | 2013 | 2014 | 2015 | 2012 | 2013 | 2014 | ||
| Apodemus argenteus | 43 | 13 | 62 | 18 | 30 | 2 | 168 | ||||||||||
| Apodemus speciosus | 43 | 9 | 87 | 7 | 29 | 1 | 13 | 189 | |||||||||
| Bandicota bengalensis | 18 | 2 | 4 | 24 | |||||||||||||
| Bandicota indica | 2 | 10 | 12 | ||||||||||||||
| Bandicota savilei | 4 | 4 | |||||||||||||||
| Berylmys berdmorei | 3 | 3 | |||||||||||||||
| Berylmys bowersi | 1 | 2 | 1 | 4 | |||||||||||||
| Dacnomys millardi | 1 | 1 | |||||||||||||||
| Eothenomys andersoni | 2 | 2 | |||||||||||||||
| Eothenomys smithi | 4 | 4 | |||||||||||||||
| Eothenomys sp. | 1 | 1 | |||||||||||||||
| Leopoldamys sabanus | 1 | 1 | |||||||||||||||
| Maxomys moi | 1 | 1 | |||||||||||||||
| Maxomys surifer | 1 | 11 | 12 | ||||||||||||||
| Micromys minutus | 2 | 2 | |||||||||||||||
| Microtus montebelli | 1 | 1 | |||||||||||||||
| Mus caroli | 10 | 1 | 11 | ||||||||||||||
| Mus cookii | 1 | 1 | |||||||||||||||
| Mus fragilicauda | 1 | 1 | |||||||||||||||
| Mus lepidoides | 1 | 1 | |||||||||||||||
| Mus musculus | 9 | 4 | 25 | 1 | 7 | 46 | |||||||||||
| Mus musculus castaneus | 1 | 1 | 5 | 3 | 10 | ||||||||||||
| Mus musculus musculus | 3 | 3 | |||||||||||||||
| Mus nitidulus | 11 | 3 | 14 | ||||||||||||||
| Mus pahari | 1 | 1 | |||||||||||||||
| Myodes rex | 1 | 1 | 2 | ||||||||||||||
| Myodes rufocanus bedfordiae | 5 | 7 | 27 | 3 | 36 | 3 | 12 | 93 | |||||||||
| Myodes rutilus mikado | 1 | 1 | 4 | 6 | |||||||||||||
| Niviventer cf. confucianus | 2 | 32 | 34 | ||||||||||||||
| Niviventer fulvescens | 1 | 1 | 2 | ||||||||||||||
| Niviventer sp. | 1 | 1 | |||||||||||||||
| Niviventer huang | 6 | 6 | |||||||||||||||
| Niviventer langbianis | 1 | 1 | |||||||||||||||
| Rattus andamanensis | 13 | 13 | |||||||||||||||
| Rattus exulans | 3 | 8 | 7 | 22 | 40 | ||||||||||||
| Rattus nitidus | 2 | 2 | |||||||||||||||
| Rattus norvegicus | 28 | 26 | 4 | 2 | 5 | 65 | |||||||||||
| Rattus rattus species complex | 1 | 8 | 2 | 110 | 3 | 1 | 7 | 4 | 136 | ||||||||
| Rattus tanezumi species complex | 44 | 1 | 4 | 5 | 5 | 1 | 1 | 61 | |||||||||
| Rattus tiomanicus | 2 | 2 | |||||||||||||||
| Total | 94 | 65 | 213 | 37 | 126 | 121 | 79 | 2 | 22 | 1 | 34 | 45 | 18 | 20 | 67 | 37 | 981 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikuchi, F.; Senoo, K.; Arai, S.; Tsuchiya, K.; Sơn, N.T.; Motokawa, M.; Ranorosoa, M.C.; Bawm, S.; Lin, K.S.; Suzuki, H.; et al. Rodent-Borne Orthohantaviruses in Vietnam, Madagascar and Japan. Viruses 2021, 13, 1343. https://doi.org/10.3390/v13071343
Kikuchi F, Senoo K, Arai S, Tsuchiya K, Sơn NT, Motokawa M, Ranorosoa MC, Bawm S, Lin KS, Suzuki H, et al. Rodent-Borne Orthohantaviruses in Vietnam, Madagascar and Japan. Viruses. 2021; 13(7):1343. https://doi.org/10.3390/v13071343
Chicago/Turabian StyleKikuchi, Fuka, Kae Senoo, Satoru Arai, Kimiyuki Tsuchiya, Nguyễn Trường Sơn, Masaharu Motokawa, Marie Claudine Ranorosoa, Saw Bawm, Kyaw San Lin, Hitoshi Suzuki, and et al. 2021. "Rodent-Borne Orthohantaviruses in Vietnam, Madagascar and Japan" Viruses 13, no. 7: 1343. https://doi.org/10.3390/v13071343
APA StyleKikuchi, F., Senoo, K., Arai, S., Tsuchiya, K., Sơn, N. T., Motokawa, M., Ranorosoa, M. C., Bawm, S., Lin, K. S., Suzuki, H., Unno, A., Nakata, K., Harada, M., Tanaka-Taya, K., Morikawa, S., Suzuki, M., Mizutani, T., & Yanagihara, R. (2021). Rodent-Borne Orthohantaviruses in Vietnam, Madagascar and Japan. Viruses, 13(7), 1343. https://doi.org/10.3390/v13071343