
Identification of Receptor Binding Proteins in Flagellotropic Agrobacterium Phage 7-7-1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Media and Growth Conditions
2.3. Phage Propagation and Purification
2.4. Phage 7-7-1 DNA Library Construction
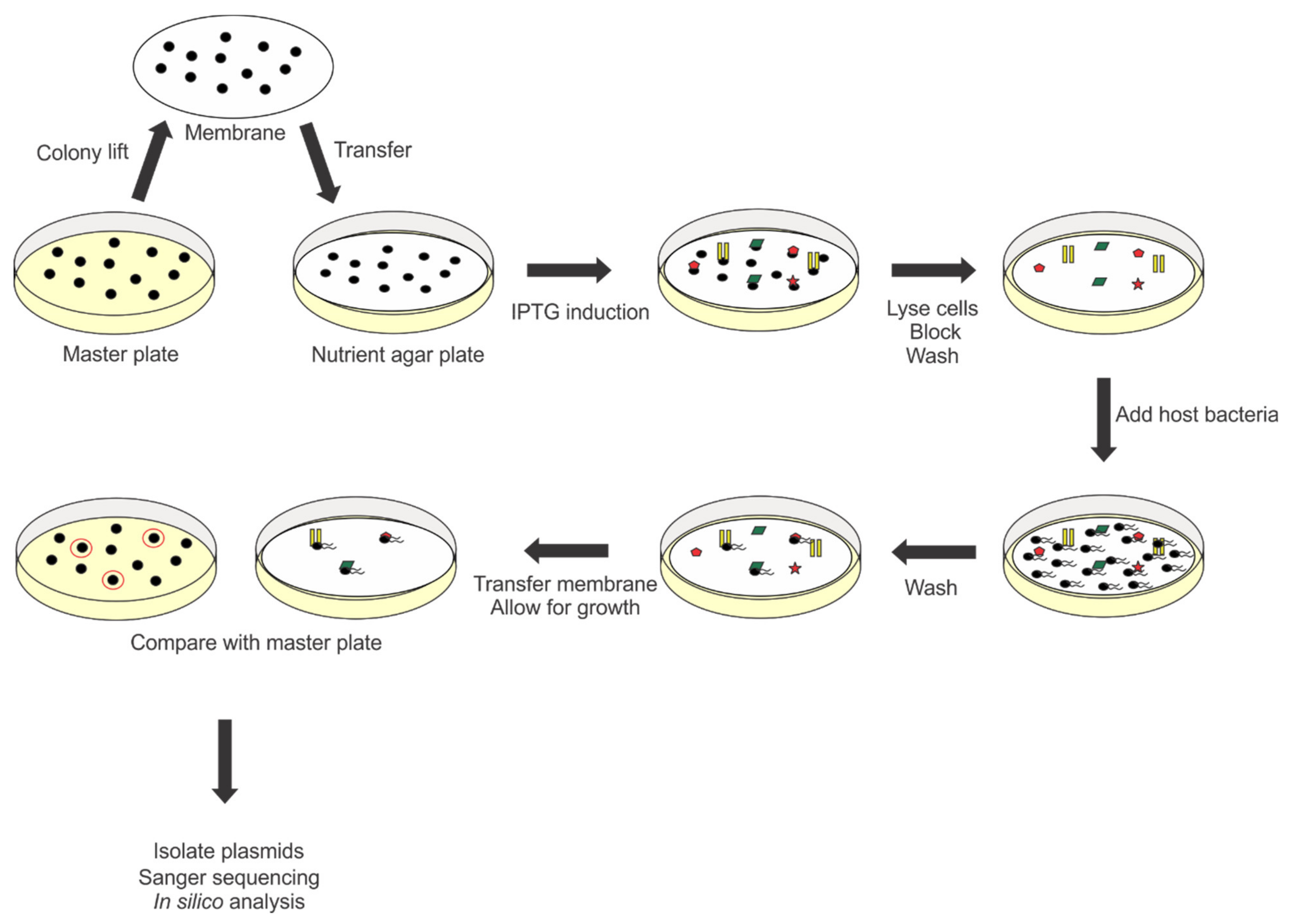
2.5. RBP Screen
2.6. Genetic Construction of Recombinant Plasmids and Bacterial Strains
2.7. Recombinant Protein Expression and Purification
2.8. Liquid Clearance Assay
2.9. Growth Curve Experiments
2.10. Lawn Clearance Experiments
3. Results
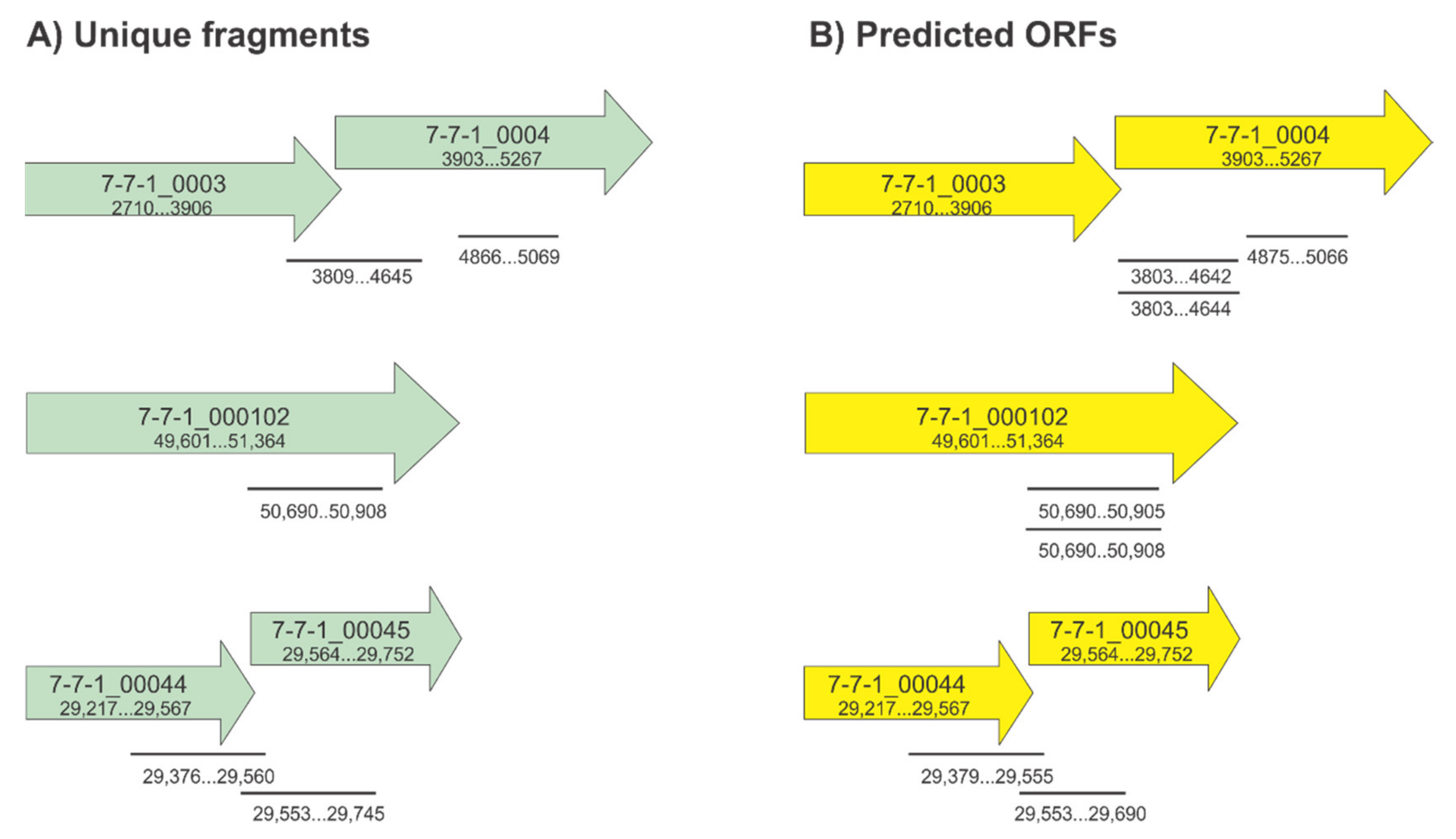
3.1. Construction of the Randomly Fragmented Phage 7-7-1 DNA Library and Identification of Candidate RBPs
3.2. Predicted Functions of Gp4, Gp102, and Gp44
3.3. Confirmation of Initial RBP Screen and Identification of Functional Plasmid Constructs
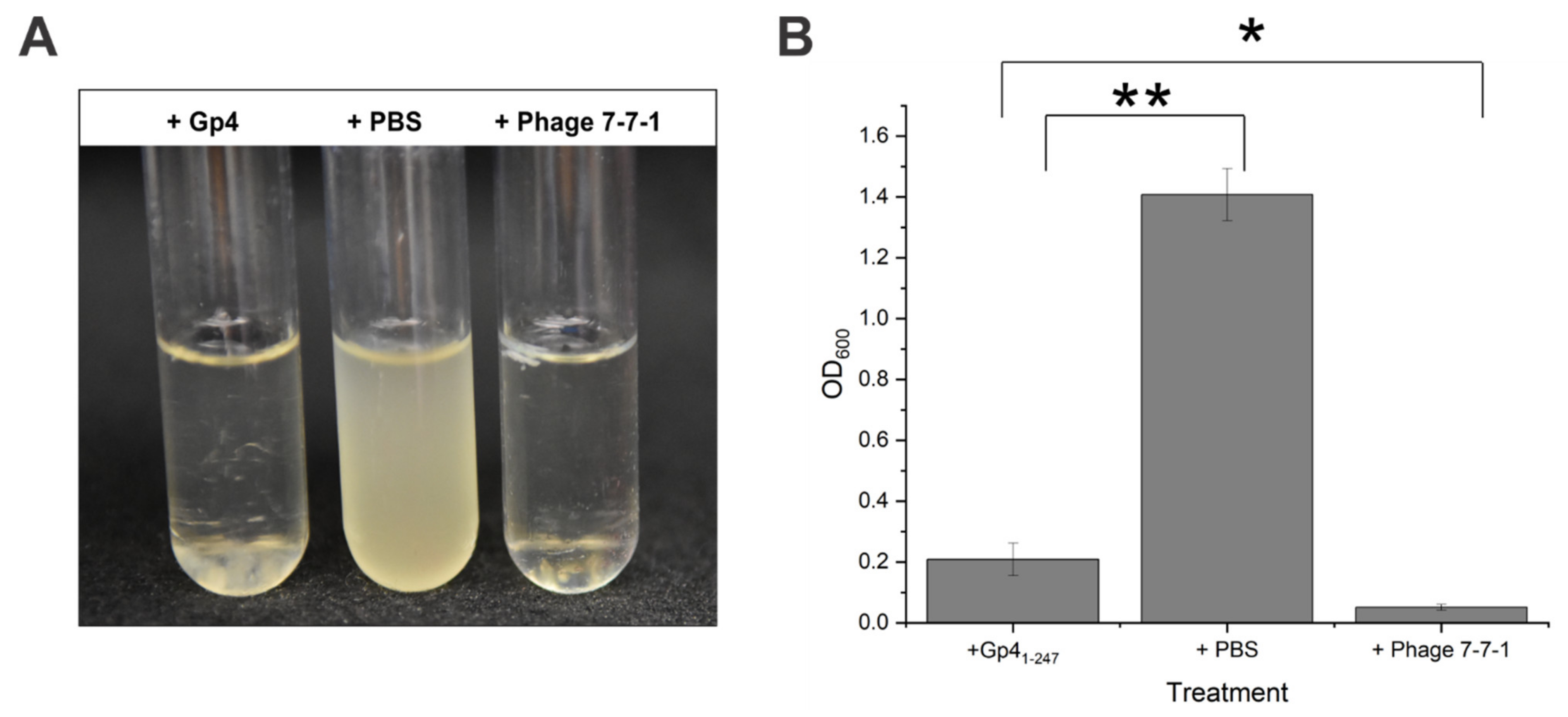
3.4. Purified Gp41-247 Inhibits the Growth of Host Cells in Liquid Cultures
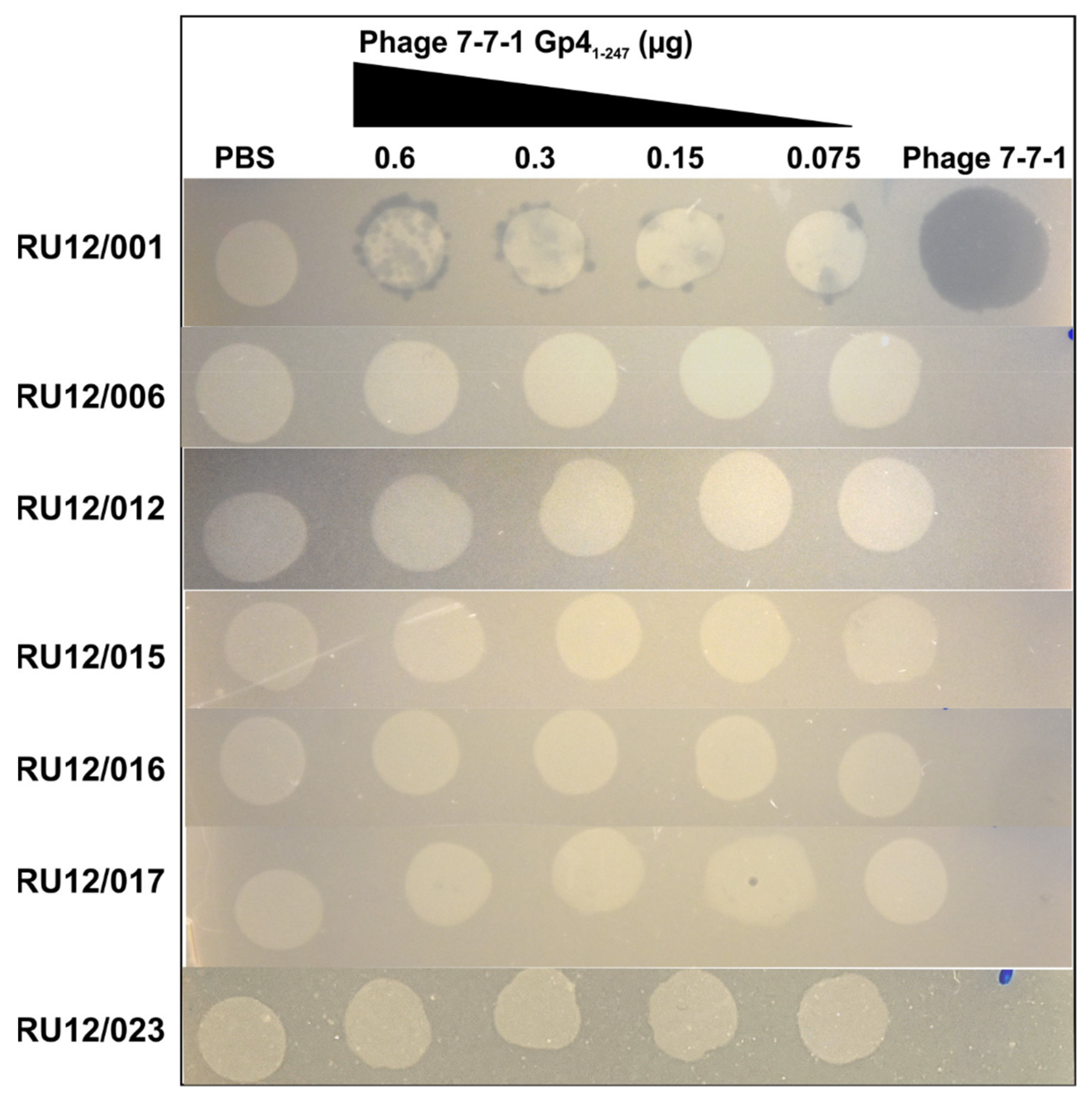
3.5. Lawn Clearance Effects of Purified Gp41-247
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Abedon, S.T. Bacteriophages and their enzymes in biofilm control. Curr. Pharm. Des. 2015, 21, 85–99. [Google Scholar] [CrossRef]
- Adriaenssens, E.; Brister, J.R. How to name and classify your phage: An informal guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortright, K.E.; Chan, B.K.; Turner, P.E. High-throughput discovery of phage receptors using transposon insertion sequencing of bacteria. Proc. Natl. Acad. Sci. USA 2020, 117, 18670. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, P.A.; von Meijenfeldt, F.A.B.; Costa, A.R.; Nobrega, F.L.; Brouns, S.J.J.; Dutilh, B.E. Adsorption Sequencing as a Rapid Method to Link Environmental Bacteriophages to Hosts. iScience 2020, 23, 101439. [Google Scholar] [CrossRef]
- Adler, B.A.; Zhong, C.; Liu, H.; Kutter, E.; Deutschbauer, A.M.; Mutalik, V.K.; Arkin, A.P. Systematic Discovery of Salmonella Phage-Host Interactions via High-Throughput Genome-Wide Screens. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lindberg, A.A. Bacteriophage Receptors. Annu. Rev. Microbiol. 1973, 27, 205–241. [Google Scholar] [CrossRef] [PubMed]
- Letarov, A.V.; Kulikov, E.E. Adsorption of bacteriophages on bacterial cells. Biochemistry 2017, 82, 1632–1658. [Google Scholar] [CrossRef]
- Schwartz, M. Interaction of Phages with their Receptor Proteins. In Virus Receptors: Part 1 Bacterial Viruses; Randall, L.L., Philipson, L., Eds.; Springer: Dordrecht, The Netherlands, 1980; pp. 59–94. [Google Scholar]
- Tremblay, D.M.; Tegoni, M.; Spinelli, S.; Campanacci, V.; Blangy, S.; Huyghe, C.; Desmyter, A.; Labrie, S.; Moineau, S.; Cambillau, C. Receptor-Binding Protein of Lactococcus lactis Phages: Identification and Characterization of the Saccharide Receptor-Binding Site. J. Bacteriol. 2006, 188, 2400–2410. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Ferreira, R.C.; Viollier, P.H.; Ely, B.; Poindexter, J.S.; Georgieva, M.; Jensen, G.J.; Wright, E.R. Alternative mechanism for bacteriophage adsorption to the motile bacterium Caulobacter crescentus. Proc. Natl. Acad. Sci. USA 2011, 108, 9963–9968. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, F.; Helm, R.F.; Broadway, K.M.; Scharf, B.E. More than Rotating Flagella: Lipopolysaccharide as a Secondary Receptor for Flagellotropic Phage 7-7-1. J. Bacteriol. 2018, 200, e00363-18. [Google Scholar] [CrossRef] [Green Version]
- Esteves, N.C.; Porwollik, S.; McClelland, M.; Scharf, B.E. The multi-drug efflux system AcrABZ-TolC is essential for infection of Salmonella Typhimurium by the flagellum-dependent bacteriophage Chi. J. Virol. 2021. [Google Scholar] [CrossRef]
- Waseh, S.; Hanifi-Moghaddam, P.; Coleman, R.; Masotti, M.; Ryan, S.; Foss, M.; MacKenzie, R.; Henry, M.; Szymanski, C.M.; Tanha, J. Orally administered P22 phage tailspike protein reduces Salmonella colonization in chickens: Prospects of a novel therapy against bacterial infections. PLoS ONE 2010, 5, e13904. [Google Scholar] [CrossRef]
- Squeglia, F.; Maciejewska, B.; Łątka, A.; Ruggiero, A.; Briers, Y.; Drulis-Kawa, Z.; Berisio, R. Structural and Functional Studies of a Klebsiella Phage Capsule Depolymerase Tailspike: Mechanistic Insights into Capsular Degradation. Structure 2020, 28, 613–624.e4. [Google Scholar] [CrossRef]
- Poshtiban, S.; Javed, M.A.; Arutyunov, D.; Singh, A.; Banting, G.; Szymanski, C.M.; Evoy, S. Phage receptor binding protein-based magnetic enrichment method as an aid for real time PCR detection of foodborne bacteria. Analyst 2013, 138, 5619–5626. [Google Scholar] [CrossRef]
- Denyes, J.M.; Dunne, M.; Steiner, S.; Mittelviefhaus, M.; Weiss, A.; Schmidt, H.; Klumpp, J.; Loessner, M.J. Modified Bacteriophage S16 Long Tail Fiber Proteins for Rapid and Specific Immobilization and Detection Salmonella Cells. Appl. Environ. Microbiol. 2017, 83, e00277-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, M.; Rupf, B.; Tala, M.; Qabrati, X.; Ernst, P.; Shen, Y.; Sumrall, E.; Heeb, L.; Plückthun, A.; Loessner, M.J.; et al. Reprogramming Bacteriophage Host Range through Structure-Guided Design of Chimeric Receptor Binding Proteins. Cell Rep. 2019, 29, 1336–1350.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbacher, S.; Seckler, R.; Miller, S.; Steipe, B.; Huber, R.; Reinemer, P. Crystal structure of P22 tailspike protein: Interdigitated subunits in a thermostable trimer. Science 1994, 265, 383. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.; Fiedler, C.; Grassl, R.; Biebl, M.; Rachel, R.; Hermo-Parrado, X.L.; Llamas-Saiz, A.L.; Seckler, R.; Miller, S.; van Raaij, M.J. Structure of the Receptor-Binding Protein of Bacteriophage Det7: A Podoviral Tail Spike in a Myovirus. J. Virol. 2008, 82, 2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, D.J.; Sacher, J.C.; Szymanski, C.M. Development of an Assay for the Identification of Receptor Binding Proteins from Bacteriophages. Viruses 2016, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M.; Van den Bossche, A.; Lavigne, R.; Noben, J.-P.; Babinger, P.; Schmitt, R. Genome and proteome analysis of 7-7-1, a flagellotropic phage infecting Agrobacterium sp. H13-3. Virol. J. 2012, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, D.J.; Sacher, J.C.; Szymanski, C.M. Exploring the interactions between bacteriophage-encoded glycan binding proteins and carbohydrates. Curr. Opin. Struct. Biol. 2015, 34, 69–77. [Google Scholar] [CrossRef]
- Steinbacher, S.; Baxa, U.; Miller, S.; Weintraub, A.; Seckler, R.; Huber, R. Crystal structure of phage P22 tailspike protein complexed with Salmonella sp. O-antigen receptors. Proc. Natl. Acad. Sci. USA 1996, 93, 10584. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-R.; Lin, T.-L.; Pan, Y.-J.; Hsieh, P.-F.; Wang, J.-T. Isolation of a Bacteriophage Specific for a New Capsular Type of Klebsiella pneumoniae and Characterization of Its Polysaccharide Depolymerase. PLoS ONE 2013, 8, e70092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.-L.; Hsieh, P.-F.; Huang, Y.-T.; Lee, W.-C.; Tsai, Y.-T.; Su, P.-A.; Pan, Y.-J.; Hsu, C.-R.; Wu, M.-C.; Wang, J.-T. Isolation of a Bacteriophage and Its Depolymerase Specific for K1 Capsule of Klebsiella pneumoniae: Implication in Typing and Treatment. J. Infect. Dis. 2014, 210, 1734–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Paff, M.L.; Molineux, I.J.; Bull, J.J. Therapeutic Application of Phage Capsule Depolymerases against K1, K5, and K30 Capsulated E. coli in Mice. Front. Microbiol. 2017, 8, 2257. [Google Scholar] [CrossRef]
- Majkowska-Skrobek, G.; Łątka, A.; Berisio, R.; Maciejewska, B.; Squeglia, F.; Romano, M.; Lavigne, R.; Struve, C.; Drulis-Kawa, Z. Capsule-Targeting Depolymerase, Derived from Klebsiella KP36 Phage, as a Tool for the Development of Anti-Virulent Strategy. Viruses 2016, 8, 324. [Google Scholar] [CrossRef]
- Stummeyer, K.; Dickmanns, A.; Mühlenhoff, M.; Gerardy-Schahn, R.; Ficner, R. Crystal structure of the polysialic acid–degrading endosialidase of bacteriophage K1F. Nat. Struct. Mol. Biol. 2005, 12, 90–96. [Google Scholar] [CrossRef]
- Schwarzer, D.; Browning, C.; Stummeyer, K.; Oberbeck, A.; Mühlenhoff, M.; Gerardy-Schahn, R.; Leiman, P.G. Structure and biochemical characterization of bacteriophage phi92 endosialidase. Virology 2015, 477, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Andres, D.; Roske, Y.; Doering, C.; Heinemann, U.; Seckler, R.; Barbirz, S. Tail morphology controls DNA release in two Salmonella phages with one lipopolysaccharide receptor recognition system. Mol. Micro. 2012, 83, 1244–1253. [Google Scholar] [CrossRef]
- Olszak, T.; Shneider, M.M.; Latka, A.; Maciejewska, B.; Browning, C.; Sycheva, L.V.; Cornelissen, A.; Danis-Wlodarczyk, K.; Senchenkova, S.N.; Shashkov, A.S.; et al. The O-specific polysaccharide lyase from the phage LKA1 tailspike reduces Pseudomonas virulence. Sci. Rep. 2017, 7, 16302. [Google Scholar] [CrossRef] [Green Version]
- Barbirz, S.; Müller, J.J.; Uetrecht, C.; Clark, A.J.; Heinemann, U.; Seckler, R. Crystal structure of Escherichia coli phage HK620 tailspike: Podoviral tailspike endoglycosidase modules are evolutionarily related. Mol. Micro. 2008, 69, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.J.; Barbirz, S.; Heinle, K.; Freiberg, A.; Seckler, R.; Heinemann, U. An Intersubunit Active Site between Supercoiled Parallel β Helices in the Trimeric Tailspike Endorhamnosidase of Shigella flexneri Phage Sf6. Structure 2008, 16, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Bales, P.; Greenfield, J.; Heselpoth, R.D.; Nelson, D.C.; Herzberg, O. Crystal Structure of ORF210 from E. coli O157:H1 Phage CBA120 (TSP1), a Putative Tailspike Protein. PLoS ONE 2014, 9, e93156. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.E.; Pourhossein, M.; Waterhouse, A.; Hudson, T.; Goldrick, M.; Derrick, J.P.; Roberts, I.S. The K5 Lyase KflA Combines a Viral Tail Spike Structure with a Bacterial Polysaccharide Lyase Mechanism. J. Biol. Chem. 2010, 285, 23963–23969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokhorov, N.S.; Riccio, C.; Zdorovenko, E.L.; Shneider, M.M.; Browning, C.; Knirel, Y.A.; Leiman, P.G.; Letarov, A.V. Function of bacteriophage G7C esterase tailspike in host cell adsorption. Mol. Micro. 2017, 105, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M.; Arutyunov, D.; Foss, M.; Cunningham, A.; Ding, W.; Singh, A.; Pavlov, A.R.; Henry, M.; Evoy, S.; Kelly, J. Genome and proteome of Campylobacter jejuni bacteriophage NCTC 12673. Appl. Environ. Microbiol. 2011, 77, 8265–8271. [Google Scholar] [CrossRef] [Green Version]
- Lotz, W.; Acker, G.; Schmitt, R. Bacteriophage 7-7-1 Adsorbs to the Complex Flagella of Rhizobium lupini H13-3. J. Gen. Virol. 1977, 34, 9–17. [Google Scholar] [CrossRef]
- Yen, J.Y.; Broadway, K.M.; Scharf, B.E. Minimum requirements of flagellation and motility for infection of Agrobacterium sp. strain H13-3 by flagellotropic bacteriophage 7-7-1. Appl. Environ. Microbiol. 2012, 78, 7216–7222. [Google Scholar] [CrossRef] [Green Version]
- Schade, S.Z.; Adler, J.; Ris, H. How bacteriophage chi attacks motile bacteria. J. Virol. 1967, 1, 599–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Shin, H.; Lee, J.-H.; Ryu, S. Identification and Characterization of a Novel Flagellum-Dependent Salmonella Infecting Bacteriophage, iEPS5. Appl. Environ. Microbiol. 2013, 79, 4829–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montemayor, E.J.; Ploscariu, N.T.; Sanchez, J.C.; Parrell, D.; Dillard, R.S.; Shebelut, C.W.; Ke, Z.; Guerrero-Ferreira, R.C.; Wright, E.R. Flagellar Structures from the Bacterium Caulobacter crescentus and Implications for Phage ϕCbK Predation of Multiflagellin Bacteria. J. Bacteriol. 2021, 203, e00399-20. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, L.M.; Lundh, N.P.; Martinez, R.J. Primary Adsorption Site of Phage PBS1: The Flagellum of Bacillus subtilis. J. Virol. 1968, 2, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Javed, M.A.; van Alphen, L.B.; Sacher, J.; Ding, W.; Kelly, J.; Nargang, C.; Smith, D.F.; Cummings, R.D.; Szymanski, C.M. A receptor-binding protein of Campylobacter jejuni bacteriophage NCTC 12673 recognizes flagellin glycosylated with acetamidino-modified pseudaminic acid. Mol. Micro. 2015, 95, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Manson, M.D.; Tedesco, P.; Berg, H.C.; Harold, F.M.; Van der Drift, C. A protonmotive force drives bacterial flagella. Proc. Natl. Acad. Sci. USA 1977, 74, 3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attmannspacher, U.; Scharf, B.; Schmitt, R. Control of speed modulation (chemokinesis) in the unidirectional rotary motor of Sinorhizobium meliloti. Mol. Microbiol. 2005, 56, 708–718. [Google Scholar] [CrossRef]
- Scharf, B. Real-Time Imaging of Fluorescent Flagellar Filaments of Rhizobium lupini H13-3: Flagellar Rotation and pH-Induced Polymorphic Transitions. J. Bacteriol. 2002, 184, 5979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, A.D.T.; Pitta, T.P.; Ryu, W.S.; Danese, P.N.; Leung, E.C.W.; Berg, H.C. Flagellar determinants of bacterial sensitivity to χ-phage. Proc. Natl. Acad. Sci. USA 1999, 96, 9863–9866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gábor, M. Transformation of streptomycin markers in rough strains of Rhizobium lupini. II. The relation between the derterminant of streptomycin dependence and those for streptomycin resistance and sensitiveness. Genetics 1965, 52, 905–913. [Google Scholar] [CrossRef]
- Hanahan, D.; Meselson, M. Plasmid screening at high colony density. Gene 1980, 10, 63–67. [Google Scholar] [CrossRef]
- Simon, R.; O’Connell, M.; Labes, M.; Pühler, A. Plasmid vectors for the genetic analysis and manipulation of rhizobia and other gram-negative bacteria. Methods Enzymol. 1986, 118, 640–659. [Google Scholar] [PubMed]
- Scharf, B.; Schuster-Wolff-Bühring, H.; Rachel, R.; Schmitt, R. Mutational Analysis of the Rhizobium lupini H13-3 and Sinorhizobium meliloti Flagellin Genes: Importance of Flagellin A for Flagellar Filament Structure and Transcriptional Regulation. J. Bacteriol. 2001, 183, 5334–5342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [Green Version]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30 (Suppl. 1), S162–S173. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Wagner, S.; Klepsch, M.M.; Schlegel, S.; Appel, A.; Draheim, R.; Tarry, M.; Högbom, M.; van Wijk, K.J.; Slotboom, D.J.; Persson, J.O.; et al. Tuning Escherichia coli for membrane protein overexpression. Proc. Natl. Acad. Sci. USA 2008, 105, 14371–14376. [Google Scholar] [CrossRef] [Green Version]
- Plattner, M.; Shneider, M.M.; Arbatsky, N.P.; Shashkov, A.S.; Chizhov, A.O.; Nazarov, S.; Prokhorov, N.S.; Taylor, N.M.I.; Buth, S.A.; Gambino, M.; et al. Structure and Function of the Branched Receptor-Binding Complex of Bacteriophage CBA120. J. Mol. Biol. 2019, 431, 3718–3739. [Google Scholar] [CrossRef]
- Xiang, Y.; Rossmann, M.G. Structure of bacteriophage phi29 head fibers has a supercoiled triple repeating helix-turn-helix motif. Proc. Natl. Acad. Sci. USA 2011, 108, 4806–4810. [Google Scholar] [CrossRef] [Green Version]
- Berg, H.C. Constraints on models for the flagellar rotary motor. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 491–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trachtenberg, S.; DeRosier, D.J.; Macnab, R.M. Three-dimensional structure of the complex flagellar filament of Rhizobium lupini and its relation to the structure of the plain filament. J. Mol. Biol. 1987, 195, 603–620. [Google Scholar] [CrossRef]
- Trachtenberg, S.; DeRosier, D.J.; Aizawa, S.; Macnab, R.M. Pairwise perturbation of flagellin subunits. The structural basis for the differences between plain and complex bacterial flagellar filaments. J. Mol. Biol. 1986, 190, 569–576. [Google Scholar] [CrossRef]
- Javed, M.A.; Sacher, J.C.; van Alphen, L.B.; Patry, R.T.; Szymanski, C.M. A Flagellar Glycan-Specific Protein Encoded by Campylobacter Phages Inhibits Host Cell Growth. Viruses 2015, 7, 6661–6674. [Google Scholar] [CrossRef] [Green Version]
- Sacher, J.C.; Shajahan, A.; Butcher, J.; Patry, R.T.; Flint, A.; Hendrixson, D.R.; Stintzi, A.; Azadi, P.; Szymanski, C.M. Binding of Phage-Encoded FlaGrab to Motile Campylobacter jejuni Flagella Inhibits Growth, Downregulates Energy Metabolism, and Requires Specific Flagellar Glycans. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Ardissone, S.; Kint, N.; Viollier, P.H. Specificity in glycosylation of multiple flagellins by the modular and cell cycle regulated glycosyltransferase FlmG. eLife 2020, 9, e60488. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Plasmid | Relevant Characteristics a | Source or Reference |
|---|---|---|
| Strain | ||
| E. coli | ||
| BL21 (DE3) | F− ompT hsdSB (rB− mB−) gal dcm λ (DE3) | Novagen |
| DH5α | recA1 endA1 | [53] |
| Lemo21 (DE3) | fhuA2 [lon] ompT gal (λ DE3) [dcm] ∆hsdS/ pLemo(Cmr) pLemo = pACYC184-PrhaBAD-lysY | NEB |
| S17-1 | recA endA thi hsdR RP4-2 Tc::Mu::Tn7 Tpr Smr | [54] |
| TOP10 | F- mcrA Δ(mrr-hsdRMS-mcrBC) Φ80lacZΔM15 Δ lacX74 recA1 araD139 Δ(araleu)7697 galU galK rpsL (Smr) endA1 nupG | Gift from Earl Petzold |
| Agrobacterium sp. H13-3 | ||
| RU12/001 | Smr; spontaneous streptomycin-resistant wild-type strain | [55] |
| RU12/006 | Smr; ΔflaA, ΔflaB, ΔflaD | [55] |
| RU12/012 | Smr; ΔmotA | [42] |
| RU12/015 | Smr; ΔAGROH133_08824 | This study |
| RU12/016 | Smr; ΔAGROH133_07337 | This study |
| RU12/017 | Smr; ΔAGROH133_13050 | This study |
| RU12/023 | Smr; ΔfliK | This study |
| Plasmid | ||
| pET30a | Kmr, expression vector (N- or C- terminal 6x histidine tag) | Gift from Christine Szymanski |
| pBS1218 | Kmr, 7-7-1_0004 fragment (741 bp) cloned into pET30a restriction sites NcoI/XhoI with N-terminal 6x histidine tag | This study |
| pBS1219 | Kmr, 7-7-1_0004 fragment (741 bp) cloned into pET30a restriction sites BamHI/SacI with C-terminal 6x histidine tag | This study |
| pBS1220 | Kmr, full length 7-7-1_000102 (1758 bp) cloned into pET30a restriction sites NcoI/BamHI with N-terminal 6x histidine tag | This study |
| pBS1221 | Kmr, full length 7-7-1_000102 (1761 bp) cloned into pET30a restriction sites NcoI/XhoI with C-terminal 6x histidine tag | This study |
| pBS1222 | Kmr, full length 7-7-1_0004 (1362 bp) cloned into pET30a restriction sites BamHI/SacI with N-terminal 6x histidine tag | This study |
| pBS1223 | Kmr, full length 7-7-1_00044 (348 bp) cloned into pET30a restriction sites BamHI/SacI with N-terminal 6x histidine tag | This study |
| pBS1224 | Kmr, full length 7-7-1_00044 (348 bp) cloned into pET30a restriction sites SalI/XhoI with C-terminal 6x histidine tag | This study |
| pBS1225 | Kmr, 7-7-1_00044 fragment (189 bp) cloned into pET30a restriction sites BamHI/SacI with N-terminal 6x histidine tag | This study |
| pBS1226 | Kmr, 7-7-1_00044 fragment (189 bp) cloned into pET30a restriction sites SalI/XhoI with C-terminal 6x histidine tag | This study |
| pBS1227 | Kmr, 7-7-1_000102 fragment (222 bp) cloned into pET30a restriction sites NcoI/XhoI with C-terminal 6x histidine tag | This study |
| pBS1228 | Kmr, full length 7-7-1_0004 cloned into pET30a restriction sites NcoI/XhoI with C-terminal 6x histidine tag | This study |
| pK18mobsacB | Kmr; lacZ mob sacB | [56] |
| Gene and Location | ID | Fragment Range (bp) | # of Unique Fragments | Coverage per Gene (bp) | Snapgene Predicted ORF Range | N or C Terminal His Tag |
|---|---|---|---|---|---|---|
| 7-7-1_0003 (2710…3906) 7-7-1_0004 (3903…5267) | FG7 FG8 FG15 FG19-1 FG28 FG89 | 3809…4645 3809…4645 4866…5069 3809…4645 3809…4645 3809…4645 | 2 | 16/1197 (03) | 3903…4644 3903…4642 4875…5066 3903…4642 4875…5066 3903…4642 | C C N C N C |
| 741/1365 (04) | ||||||
| or | ||||||
| 204/1365 (04) | ||||||
| 7-7-1_000102 (49,601…51,634) | FG9 | 50,690…50,908 | 2 | 219/1764 (102) | 50,693…50,905 | N |
| FG10 | 50,690…50,908 | 50,693…50,905 | N | |||
| FG26 | 50,690…50,908 | 50,690…50,908 | N | |||
| 46,134…46,017 | ||||||
| FG44 | 50,690…50,908 | 50,693…50,905 | N | |||
| 7-7-1_00044 (29,217…29,567) 7-7-1_00045 (29,564…29,752) | FG5 FG6 FG42 | 29,376…29,560 29,376…29,560 29,553…29,745 | 2 | 185/351 (44) | 29,379…29,555 29,379…29,555 29,553…29,690 | N N N |
| or | ||||||
| 14/351 (44) | ||||||
| 127/189 (45) |
| Protein | BLASTP Function | Phyre2 | ||||
|---|---|---|---|---|---|---|
| Top Template | Amino Acids Aligned | % Coverage | % Identity | Confidence Value | ||
| Gp4 (454 AA) | Tail biosynthetic protein | Prophage MuSo2 tail protein from Shewanella oneidensis | 2-366 | 80 | 21 | 100 |
| Gp102 (587 AA) | Putative tail fiber | Hydrolase XylC from Thermoanaerobacterium saccharolyticum JW/SL-YS485 | 192-248 | 9 | 36 | 96.3 |
| Gp44 (116 AA) | Hypothetical protein | Aldolase from Kordia algicida OT-1 | 65-109 | 37 | 14 | 15 |
| Gene | Plasmids | Protein Expressed | Binding c |
|---|---|---|---|
| 7-7-1_0004 | pBS1218 | 6xHis-Gp41-247 | +++ |
| pBS1219 | Gp41-247-6xHis | ++ | |
| pBS1222 | 6xHis-Gp4 | + | |
| pBS1228 | Gp4-6xHis | + | |
| FG19-1 | Gp41-246-6xHis a | ++ | |
| pET30a | --- | + | |
| 7-7-1_000102 | pBS1220 | 6xHis-Gp102 | + |
| pBS1221 | Gp102-6xHis | ++ | |
| pBS1227 | Gp102363-436-6xHis | + | |
| FG9 | 6xHis-Gp102363-436 | +++ | |
| pET30a | --- | + | |
| 7-7-1_00044 | pBS1223 | 6xHis-Gp44 | + |
| pBS1224 | Gp44-6xHis | +++ | |
| pBS1225 | 6xHis-Gp4453-115 | ++ | |
| pBS1226 | Gp4453-115-6xHis | + | |
| FG5 | 6xHis-Gp4453-114 b | +++ | |
| pET30a | ---- | + |
| Strain | Genotype | Flagellar or Motility Phenotype | Other Phenotypes |
|---|---|---|---|
| RU12/001 | Wild type | Normal motility | n/a |
| RU12/006 | ΔflaA ΔflaB ΔflaD | No flagella | n/a |
| RU12/012 | ΔmotA | Non-motile | n/a |
| RU12/015 | ΔAGROH133_08824 | Normal motility | Mutant LPS |
| RU12/016 | ΔAGROH133_07337 | Normal motility | Mutant LPS |
| RU12/017 | ΔAGROH133_13050 | Normal motility | Mutant LPS |
| RU12/023 | ΔfliK | Non-motile; abnormal hook | n/a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez, F.; Scharf, B.E. Identification of Receptor Binding Proteins in Flagellotropic Agrobacterium Phage 7-7-1. Viruses 2021, 13, 1267. https://doi.org/10.3390/v13071267
Gonzalez F, Scharf BE. Identification of Receptor Binding Proteins in Flagellotropic Agrobacterium Phage 7-7-1. Viruses. 2021; 13(7):1267. https://doi.org/10.3390/v13071267
Chicago/Turabian StyleGonzalez, Floricel, and Birgit E. Scharf. 2021. "Identification of Receptor Binding Proteins in Flagellotropic Agrobacterium Phage 7-7-1" Viruses 13, no. 7: 1267. https://doi.org/10.3390/v13071267
APA StyleGonzalez, F., & Scharf, B. E. (2021). Identification of Receptor Binding Proteins in Flagellotropic Agrobacterium Phage 7-7-1. Viruses, 13(7), 1267. https://doi.org/10.3390/v13071267